










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.84 n.1 Pretoria Jan. 2013
CASE REPORT
Incidental Mycobacterium-induced granulomatous inflammation of the follicular pharyngeal tonsils in a South African farmed ostrich (Struthio camelus)
Martina R. CroleI; John T. SoleyI; Sarah J. CliftII
IDepartment of Anatomy and Physiology, University of Pretoria, South Africa
IIDepartment of Paraclinical Sciences, University of Pretoria, South Africa
ABSTRACT
Avian mycobacteriosis (AM) is a zoonotic disease caused by Mycobacterium avium complex (MAC), which can spread from avians to other farmed animals such as cattle and pigs as well as to humans. This study is the first report of granulomatous inflammation, as a result of avian mycobacteriosis, in the follicular pharyngeal tonsils of a farmed ostrich. The head of an apparently healthy farmed adult ostrich was obtained after slaughter. Each pharyngeal fold displayed a large tissue mass. This tissue was routinely prepared for light microscopy and stained with haematoxylin and eosin, periodic acid Schiff, Grocott methenamine silver, Gram and Ziehl-Neelsen. Immunohistochemistry (IHC) and polymerase chain reaction (PCR) were performed to identify Mycobacterium spp. and Mycobacterium tuberculosis complex, respectively. Histologically, the tissue masses consisted of confluent mature micro-granulomata that were characterised by central caseous necrosis surrounded by multinucleated giant cells, macrophages and lymphoid cells and an outer mature fibrous connective tissue capsule. Within some foci of caseous necrosis were variably sized colonies of small, Gram-negative, acid-fast bacilli, which showed positive IHC labelling for Mycobacterium spp., leading to a presumptive diagnosis of AM. PCR thus proved useful in excluding the presence of notifiable Mycobacterium spp. The significance and role of the pharyngeal tonsils of ratites in diseases such as AM warrant specific attention. Moreover, as ratites are known to present with AM infections with apparently no visible loss in body condition, as presumably occurred in the present case, it is imperative that unusual masses in apparently healthy ratites be thoroughly investigated.
Introduction
Mycobacterium avium complex (MAC) is the causative agent of a bacterial infection known as avian mycobacteriosis (AM) (García, LeClear & Gaskin 2001; Tell, Woods & Cromie 2001; Thornton et al. 1999) or avian tuberculosis (ATB) (Cooper 2005; Thornton et al. 1999). However, as classical tuberculous lesions are not the only manifestation of mycobacterial infections in birds, it is advocated that the term avian mycobacteriosis be preferentially used (Tell et al. 2001). Avian mycobacteriosis is a zoonotic disease that can spread from avians to other farmed animals such as cattle and pigs (García et al. 2001; Tully & Shane 1996) as well as to humans (Kincade 1959). The disease has been extensively researched in many avian species and a thorough review is provided by Tell et al. (2001). The many subspecies of M. avium are grouped together under the umbrella term Mycobacterium avium complex (Cromie et al. 2000; Pavlik et al. 2000; Thornton et al. 1999) and M. avium subspecies avium is reported to be the most common cause of AM (Gerlach 1994; Gyimesi et al. 1999).
Avian mycobacteriosis mainly affects the digestive system (Bowes 1993; Cooper 2005; Cromie et al. 2000; García et al. 2001; Shivaprasad 2002; Tully & Shane 1996) with secondary spread of infection to other visceral organs (Bowes 1993) where the typical nodular lesions are easily detected (Doneley et al. 1999; Huchzermeyer 1997). MAC bacilli are usually shed in the faeces of affected birds (Thornton et al. 1999) and ingestion of contaminated faeces is the most common mode of infection (Bowes 1993; Cooper 2005; García et al. 2001; Tully & Shane 1996).
Avian mycobacteriosis, although of less importance to the commercial poultry industry (Cromie et al. 2000; Sanford, Rehmtulla & Josephson 1994), still plagues poultry production in developing countries, causes losses in other commercially farmed birds such as ratites and also affects wild, endangered species in captive breeding programmes (Cromie et al. 2000). Mycobacteriosis in ratites (Sanford, Rehmtulla & Josephson 1994), including the ostrich (Struthio camelus), is considered to be rare (Bowes 1993; Cooper 2005; Huchzermeyer 1997; Kazda, Pavlik & Hruska 2009; Montali et al. 1976; Shane et al. 1993) and is restricted mainly to birds kept in zoologicalcollections (Huchzermeyer 1997). However, AM has recently been diagnosed in ostriches in Canada (Bowes 1993), the Czech Republic (Levkut et al. 1998), Slovakia (Ševčíková et al. 1999), Australia (Doneley et al. 1999), the Netherlands (Pavlik et al. 2000), Poland (Wozikowski 2001), the USA (García et al. 2001) and in Japan (Ryo, Hiroaki & Yoshihiro 2006). Avian mycobacteriosis has also been described in the emu (Dromaius novaehollandiae) (Cromie et al. 2000; Krishna Mohan Roa & Chowdary 1980; Pocknell et al. 1996; Thornton et al. 1999), the greater rhea (Rhea americana) (Sanford et al. 1994; Tacconi & Valente 1981) and the kiwi (Apteryx mantelli) (Davis, Watson & Billing 1984). Mycobacteriosis caused by M. bovis has been reported in a cassowary (Casuarius casuarius) in the Czech Republic (Pavlik et al. 2002) but this can be considered a rare event (Kazda et al. 2009).
The normal morphology of the ostrich oropharynx has been documented (Porchescu 2007; Tadjalli, Mansouri & Poostpasand 2008; Tivane 2008) and, more recently, structures that can be classified as follicular tonsils have been described in the oropharynx of both the ostrich and emu (Crole & Soley 2012a). It is clear that the pharyngeal folds typically found in these birds and the associated follicular and non-follicular pharyngeal tonsils form an extensive and immunologically active region in the ostrich oropharynx (Crole & Soley 2012a). There also appear to be no reports in the literature on any clinical conditions affecting the pharyngeal folds or tonsils of the ostrich, or those of any other ratite species. When compared with the wealth of information, including numerous case reports, available on AM in various bird species, the body of literature covering AM in ratites is small, reflecting its rarity in this group of birds. However, it is clear that this disease does occur in ratites (Sanford et al. 1994), is a global phenomenon and requires specific investigation in these commercially important birds.
This study is the first report of granulomatous inflammation, as a result of AM, in the follicular pharyngeal tonsils of an ostrich and represents the first report of AM in this species in South Africa in a period of almost 20 years (Anon 1985).
Materials and methods
Specimen collection
The head of an apparently healthy (but displaying proliferative masses of the pharyngeal folds), farmed adult ostrich was obtained after slaughter as part of a routine collection of research material for a morphological study of the oropharynx. The bird was approximately 14 months old and of unknown sex. The head had been immersion fixed in 10% neutral-buffered formalin and stored for approximately 18 months in the Department of Anatomy and Physiology, Faculty of Veterinary Science, University of Pretoria. The head was incised to reveal the oropharynx (Crole & Soley 2010). The masses were described, measured and photographed in situ using a Canon EOS 5D digital camera (Canon, Japan) with a 28 mm - 135 mm lens before being excised, together with the pharyngeal folds.
Histology
Appropriate longitudinal and transverse sections representing different areas of the masses were excised. The tissue samples were routinely prepared for light microscopy using a Shandon model 2LE Automatic Tissue Processor (Shandon, Pittsburgh, PA, USA). Sections were stained with haematoxylin and eosin (H&E), periodic acid Schiff stain (PAS) (McManus 1946), Grocott methenamine silver (GMS), Gram and Ziehl-Neelsen (ZN) and viewed and micrographed using an Olympus BX50 equipped with the analySIS CC12 Soft Imaging System (Olympus Corporation, Tokyo, Japan) and an Olympus BX63 light microscope (Olympus Corporation, Tokyo, Japan) equipped with a DP72 camera and Olympus cellSens imaging software.
Immunohistochemistry
Sections submitted for immunostaining were mounted on positively-charged microscope slides (SuperFrost® Plus, Menzel-Glasser®), and dried overnight in a 38 °C - 40 °C oven. Routine dewaxing in xylol, followed by rehydration through a graded ethanol and distilled water series, took place inside a fume hood. Sections were subsequently incubated with 3.0% hydrogen peroxide in methanol for 15 min in a humidified chamber at room temperature, then rinsed in distilled water, followed by 0.1 m phosphate buffered saline (PBS), pH 7.6 containing 0.1% bovine serum albumin (BSA) for 5 min per rinse. Heat-induced epitope retrieval (HIER) was performed via microwave heating (96 °C) in citrate buffer (pH 6) for 14 min. Thereafter the test sections (still in buffer) were allowed to cool on the bench before rinsing in distilled water and then in PBS/BSA buffer for 5 min. Sections were incubated with normal goat serum (catalogue no: G9023, Sigma) and diluted 1:5 with PBS/BSA buffer, pH 7.6 for 20 min in a humidified chamber at room temperature. The blocking serum was drained and replaced by the primary polyclonal rabbit anti-Mycobacterium bovis antibody (pAbBCG) (BCG strain; catalogue no: B 0124, DakoCytomation), at a dilution of 1:1000 in PBS/BSA buffer for 90 min in a humidified chamber at room temperature. The biotinylated polyclonal goat-anti-rabbit link antibody (catalogue no: EO432, DakoCytomation) was used at a dilution of 1:500 in PBS/BSA buffer. The link antibody was incubated with the tissue sections for 30 min in a humidified chamber at room temperature, followed by rinsing in distilled water and then in PBS/BSA buffer as before. Peroxidase conjugated avidin (Vectastain ABC Kit, Elite PK6100 Standard, Vector Laboratories) was applied and incubated on the sections for 30 min in a humidified chamber at room temperature. The sections were rinsed twice, as before. The sections were immersed in a NovaRED substrate (catalogue no: SK-4800, Vector Laboratories) in a humidified chamber at room temperature. During this time, the validated positive tissue control section (see below) with the same substrate was examined carefully for signs of positive staining at 100 x magnification, using a light microscope. As soon as there was clear evidence of positive staining (focal or multifocal red granular cytoplasmic positivity in macrophages) in the positive tissue control section, the test sections were immediately rinsed in a distilled water bath to halt the substrate reaction. The sections were then counterstained with Mayer's haematoxylin for 1 min - 2 min and rinsed under running tap water for 10 min to remove excess substrate. Sections were routinely dehydrated through increasing alcohol concentrations and xylol, mounted and coverslipped. In the negative reagent control section, the primary antibody was replaced by normal goat serum. No background staining was observed in the negative control section (not shown). The formalin-fixed, paraffin-embedded, positive control tissue was from an 18-month-old male buffalo with typical mycobacterial lesions in the tonsils. The tissue cultured positive for M. bovis (PM 71.12/ S3776.12; Section of Pathology, Department of Paraclinical Sciences, University of Pretoria). Formalin-fixed tissue from a sheep with a confirmed case (via PCR) of Johne's disease (paratuberculosis) (Western Cape State Laboratory; case number 13.044) stained strongly with the pAbBCG, indicating that this antibody also recognises non-tuberculous bacteria.
Polymerase chain reaction
Preparation of template DNA and PCR amplification for Mycobacterium tuberculosis complex (M. tuberculosis, M. bovis, Mycobacterium canettii, Mycobacterium africanum, Mycobacterium microti, Mycobacterium pinnipedii, Mycobacterium caprae, M. bovis BCG) (Tuberculosis Laboratory, Zoonotic Diseases Section, Onderstepoort Veterinary Institute, South Africa) were performed according to the method described by Miller et al. (1997) with minor modifications. Paraffin sections were placed in a centrifuge tube together with 200 µ.L of water with 0.5% Tween-20. The tube contents were boiled for 10 min and followed by snap freezing (2 min at -20 °C). This step was repeated and samples were subjected to boiling for a third time for 10 min. The tube was centrifuged at 3000 x g for 10 min to pellet the tissue. Ten µ.L of the sample was tested by PCR using primers to identify M. tuberculosis complex. Mycobacterium bovis and Mycobacterium species, other than M. tuberculosis isolates, as well as water, were included as controls. Boiled cell suspension from a field strain of M. bovis was used as a positive control. The reaction contained 20 mm of each primer, 125 U of Taq polymerase and 0.2 mm of nucleotides. Following an initial cycle of 10 min at 94 °C and 135 s at 72 °C, amplification was continued for 50 cycles of 45 s at 94 °C and 75 s at 72 °C, followed by a final 10 min extension at 72 °C. Amplification products were analysed by electrophoresis in 2.0% agarose gel stained with ethidium bromide. A band size of 123 base pairs (bp) was considered positive for M. tuberculosis complex and the absence of a band or bands at any other position was considered negative. Polymerase chain reaction for MAC was not performed on the samples as the primers were not available at the time and the test was never validated in the laboratory.
Results
Gross pathology
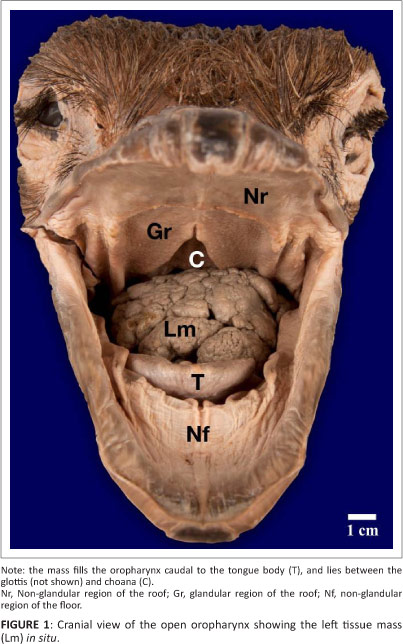
The only outward evidence of pathology of the head examined was distension of the gular region (the external region of skin between the arms of the mandible) and proximal oesophagus. Examination of the oropharynx revealed a large, round, cauliflower-like, cream-coloured (in the formalin-fixed state), firm tissue mass associated with the left side of the oropharynx, although it occupied the entire oropharynx caudal to the tongue body (Figure 1). This tissue mass was interposed between the glottis and choana (Figure 1 and Figure 2) and measured 7.0 cm x 5.0 cm with a circumference of 17.5 cm. A second tissue mass emanating from the right side of the oropharynx was visible in the proximal oesophagus in the openly displayed oropharynx (Figure 2). The right tissue mass was similar in appearance to the left tissue mass but was elongated (Figure 2 and Figure 3) and measured 8.0 cm x 4.0 cm with a circumference of 14.0 cm. The tissue masses originated from and were intimately attached to the dorsal surface of the pharyngeal folds, with the rostral, left tissue mass originating from the left pharyngeal fold and the caudal, right tissue mass originating from the right pharyngeal fold (Figure 2).
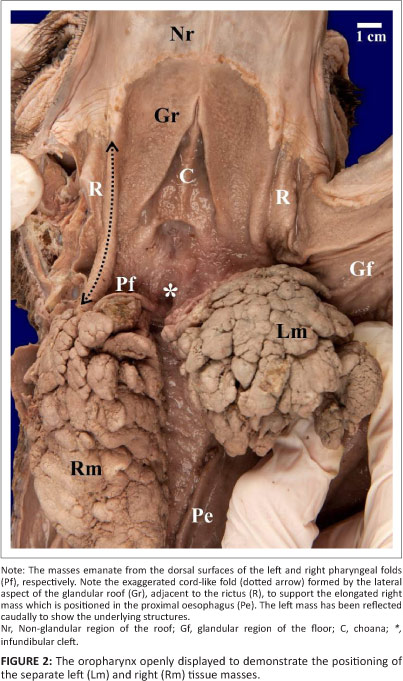
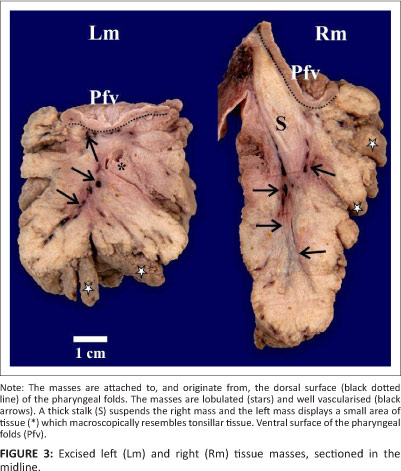
Removal of the tissue masses, together with the pharyngeal folds and proximal oesophagus revealed a highly vascularised area of supporting connective tissue in the retropharyngeal region dorsal to the attachment of the pharyngeal folds. Due to the extent of the vascularisation, this region resembled a haematoma. When sectioned in the midline, the cut surface of the tissue masses clearly demonstrated the vast proliferation of tissue emanating from the dorsal aspect of each pharyngeal fold (Figure 3). The tissue masses were well vascularised, lobulated and cream-coloured (Figure 3). The right tissue mass was suspended by a prominent mucosal fold of the glandular roof (Figure 2) which, when sectioned, was seen to be supported by a thick inner stalk (Figure 3). The left tissue mass contained a small, isolated tract of tissue that macroscopically resembled normal tonsillar tissue (Figure 3). The follicular pharyngeal tonsils (Crole & Soley 2012a) were absent and replaced by the tissue masses.
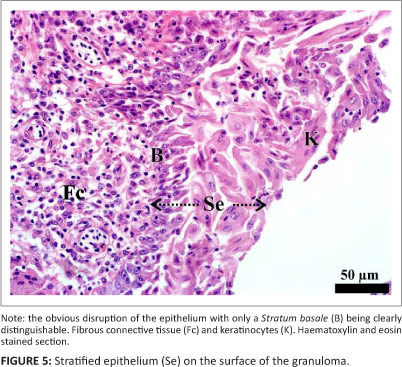
Histopathology and immunohistochemistry
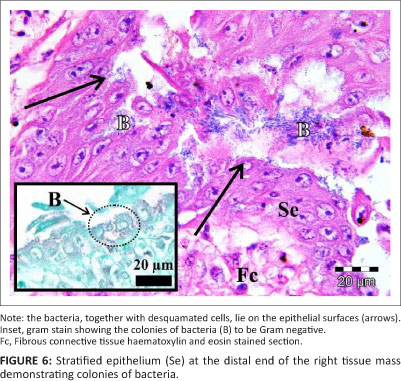
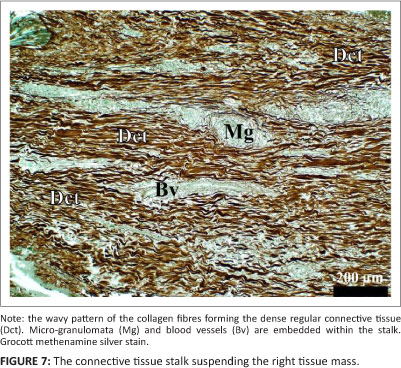
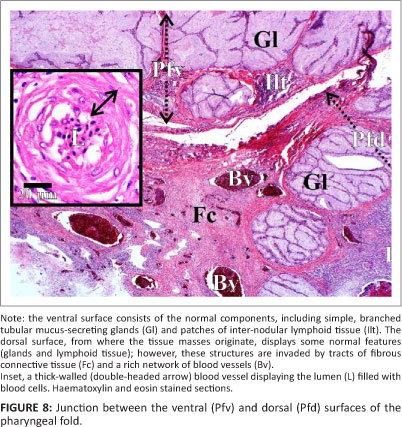
Histology revealed that the tissue masses consisted of confluent micro-granulomata (Figure 4). The surface of each mass was partly covered by a nondescript epithelium (Figure 4), that, when present, appeared to be stratified (Figure 5) and that was extensively eroded and multifocally ulcerated. The epithelium was pushed into blunt papillary folds by the underlying proliferating tissue (Figure 4). In some places, the stratified epithelium displayed signs of keratinisation (Figure 5) evidenced by dead keratinocytes and keratin flakes. Gram-negative bacteria were present, in some regions, on the surface of the epithelium (Figure 6), including areas of ulceration. The right tissue mass was suspended by a thick stalk (Figure 3) composed of dense regular connective tissue with embedded micro-granulomata and blood vessels (Figure 7). Some parts of the stalk displayed more compact regular connective tissue with thick-walled arteries and fewer micro-granulomata than in the surrounding tissue, thus indicating advanced maturity. The tissue masses were both richly supplied with blood vessels of which some were thick-walled (Figure 8).
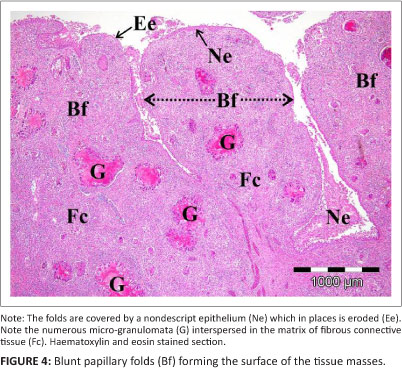
Both tissue masses consisted of numerous variably sized, coalescing micro-granulomata with mature surrounding and intervening fibrous connective tissue (Figure 4). The micro-granulomata were generally characterised by a central area of caseous necrosis with vacuolation and a peripheral rim of multinucleated giant cells (MNGCs), some of which displayed small coalescing intra-cytoplasmic vacuoles and some large, single vacuoles (Figure 9 and Figure 10). Occasional smaller foci were formed by clusters of MNGCs (Figure 10a). Within occasional foci of caseous necrosis were variably sized colonies of bacteria (Figures 10a-c). The bacteria were small, Gram-negative, acid-fast bacilli (Figure 10c) that showed positive IHC labelling for Mycobacterium spp. (Figure 10a and Figure 10b). MNGCs occasionally contained the above-mentioned bacteria or foreign material (Figure 9). Foreign particles were brown, globular, slightly refractive (Figure 9) and stained strongly with GMS (Figure 10d). There was no evidence of fungi (PAS and GMS) (Figure 10d and Figure 10e).
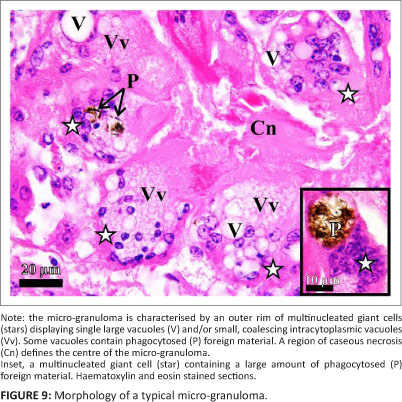
Surrounding the MNGCs was a variable admixture of epithelioid macrophages, numerous plasma cells, lymphocytes and occasional heterophils. Intermingled with the inflammatory cells were fibroblasts, collagen fibres and numerous congested blood vessels, forming peripheral granulation tissue. The fibroblasts and intervening collagen fibres occasionally formed concentric layers around the MNGCs and inflammatory cells, creating an onion-skin effect. Lymph nodules and inter-nodular lymphoid tissue were present nearer the pharyngeal folds (Figure 8); however, this tissue was markedly atrophic.
The ventral surface of the pharyngeal folds presented the normal histological structure (Figure 8) previously described for the ostrich (Crole & Soley 2012a). However, the dorsal surface of the folds was severely affected by the intrusion of micro-granulomata, with only small regions exhibiting a normal structure. The structure of both the follicular and non-follicular tonsil associated with the dorsal aspect of the pharyngeal folds (Crole & Soley 2012a) was obliterated by the proliferating zone of granulomatous inflammation (Figure 8).
Polymerase chain reaction
Polymerase chain reaction (PCR) for the detection of M. tuberculosis complex (M. tuberculosis, M. bovis, M. canettii, M. africanum, M. microti, M. pinnipedii, M. caprae, M. bovis BCG) was negative.
Discussion
This study is the first report in 20 years of AM in a farmed ostrich in South Africa. It is also the first report of AM affecting the pharyngeal tonsils in a ratite species. Mycobacteriosis is a chronic, progressive disease resulting in systemic granulomatous inflammation and in ratites usually presents as unthriftiness, loss of weight and diarrhoea (Shivaprasad 2002). Granulomatous inflammation is characteristic of AM reported in ostriches (Cooper 2005; Doneley et al. 1999; Garcia et al. 2001; Hood 1977; Ševčiková et al. 1999) as well as in other ratite species such as the greater rhea (Sanford et al. 1994), emu (Pocknell et al. 1996; Shane et al. 1993) and kiwi (Davis, Watson & Billing 1984). It was noted that lesions observed in the greater rhea were also typical of those described in other birds (poultry, psittacines and other captive exotic birds) with AM (Sanford et al. 1994). On histopathology, the lesions seen in the present study resembled the typical granulomatous inflammatory response seen in many avian infections (Shivaprasad 2002). In the ostrich, this response has been reported for infectious agents such as Pseudomonas aeruginosa (Momotani et al. 1995), Aspergillus sp. (Fitzgerald & Moisan 1995), Moraxella phenylpyruvica (Gürel, Gülçubuk & Turan 2004) as well as for non-infectious causes, such as chronic granulomatous conjunctivitis (Saroglu, Yucel & Aktas 2000). Although the reported case of granulomatous conjunctivitis was caused by M. phenylpyruvica, the authors noted the resemblance between these lesions and those seen in tuberculosis (Gürel, Gülcubuk & Turan 2004). However, in the current study, there was no evidence of fungi with either PAS or GMS staining, whereas the Gram stain indicated the presence of Gram-negative bacteria. ZN staining subsequently revealed low numbers of acid-fast bacilli, prompting a presumptive diagnosis of AM (Allen 1992; Gyimesi et al. 1999), which was further supported by positive IHC labelling for Mycobacterium spp. Immunostaining with pAbBCG (as in the present study) has proved to be a reliable ancillary method for the detection of mycobacterial organisms (Ulrichs et al. 2005). Crossed immunoelectrophoresis showed that the polyclonal antibody reacted with approximately 100 different BCG antigens, many of which were common to other Mycobacteria species (DakoCytomation information sheet B0124/RUO/OS/13.05.05). It would appear from previous studies on AM in ostriches (Bowes 1993; Doneley et al. 1999; Ševčiková et al. 1999) that low numbers of acid-fast bacilli, as observed in the present study, are typical for this condition. The distribution of bacteria, both free within the central concentration of necrotic debris and locked within the cytoplasm of the MNGCs of the micro-granulomata has also previously been reported (Doneley et al. 1999; Pocknell et al. 1996; Shane et al. 1993).
A definitive diagnosis of AM requires culture of the mycobacterium. However, this method is both slow and difficult (Gyimesi et al. 1999) and was not possible in the present study as no fresh tissue was available. The identification of Mycobacterium spp. by PCR has been used on formalin-fixed tissue in cattle and elk, where M. bovis was the causative organism (Miller et al. 1997). This technique has also been used for the diagnosis of M. avium in a duck (Thornton et al. 1999). However, this method holds its own challenges, especially if the period of formalin fixation of the tissue is lengthy (Gyimesi et al. 1999). The negative result of the PCR for the M. tuberculosis complex on the formalin-fixed tissue in the present study must be interpreted with caution due to the lengthy fixation of the tissue. Therefore, although a notifiable species was not detected, one cannot be excluded as the causative agent and nor can any of the organisms of the M. avium complex.
The association of the twin tissue masses with the pharyngeal folds raises the question of their origin and the source of inflammation. A recent study of the pharyngeal folds (Plica pharyngea) and associated lymphoid tissue in the ostrich revealed the presence of follicular pharyngeal tonsils (Tonsilla pharyngea) (located retropharyngeally), small non-follicular pharyngeal tonsils (on the dorsal surface of the pharyngeal folds) and pharyngeal lymph nodules (Lymphonoduli pharyngeales) (present on the dorsal and ventral surfaces of the pharyngeal folds) (Crole & Soley 2012a). Based on this information, it was evident that the tissue masses originated specifically from the follicular pharyngeal tonsils due to their location as well as by the almost complete obliteration of the tonsillar tissue. The ventral surface of the pharyngeal folds remained structurally normal whereas the dorsal surface and associated non-follicular pharyngeal tonsils were partially involved in the granulomatous reaction, most likely due to their proximity to the follicular pharyngeal tonsils. The location of the tissue masses raises an interesting question concerning the ability of the ostrich to ingest food and water. The feeding method of paleognathous birds has been described as cranioinertial (Bonga Tomlinson 2000) or 'catch-and-throw' (Gussekloo & Bout 2005) whereby food is accelerated from the bill tips through the oropharynx to land caudal to the laryngeal mound, in the proximal oesophagus. Simultaneously the linguo-laryngeal apparatus (Crole & Soley 2012b) would prevent food from entering the glottis. Although the bird in question would have been able to feed in this manner (Bonga Tomlinson 2000; Gussekloo & Bout 2005) while the granulomas were small, it would not have been possible near the time of slaughter due to the position and size of the granulomas, especially taking into consideration that the left granuloma occupied almost the entire caudal oropharynx and the right granuloma the proximal oesophagus. There is little doubt that the twin granulomas, had they continued to grow, would eventually have inhibited feeding, drinking and respiration.
A report of a mycotic (Aspergillus) granuloma of the nasal passages in an ostrich suggested that the inciting cause was local damage due to the inhalation of plant material (Fitzgerald & Moisan 1995). Similarly, the presence of pollen grains in the Mycobacterium-positive granulomatous reaction in the conjunctiva of an ostrich brought into question which factor (pollen or Mycobacterium) was the inciting cause and it could not be determined which factor was responsible for the initial insult (Ševčíková et al. 1999). The inciting cause of the granulomatous inflammation of the tonsils in the present study is also ambiguous as it could not be determined whether the initial reaction was caused by the impaction of feed or the mycobacterium. The tonsillar crypts of the ostrich are deep and branching (Crole & Soley 2012a), which may predispose them to trapping fine particulate matter. The foreign material had the appearance of plant matter and stained strongly with GMS, similar to that reported for AM in another ostrich (Ševčíková et al. 1999). Circumstantial evidence (foreign material and acid-fast bacilli within the micro-granulomata) points to the possible role of contaminated feed. An unusual feature in the present study was the bilateral nature of the granulomatous inflammation. The feed may have carried a high load of infection and both tonsils may have become infected due to direct exposure.
Conclusion
True pharyngeal tonsils have recently been identified in birds and their structure has been described in the ostrich and emu (Crole & Soley 2012a). Tonsils have been identified in mammals as very important tissues for diagnostic sampling for infectious diseases (Liebler-Tenorio & Reinhard 2006) and in birds the pharyngeal tonsil is prone to the formation of micro-abscesses after infection (Casteleyn et al. 2010). The significance and role of the pharyngeal tonsils of ratites in diseases such as AM warrant specific attention. It is noteworthy that although AM in ratite species is considered to be rare, many cases, like the present study, do not present as typical AM (Bowes 1993; García et al. 2001; Hood 1977; Nouvel & Leclerc-Cassan 1970; Pocknell et al. 1996; Ševčíková et al. 1999). In the present study, there were presumably no noticeable signs of loss of body condition, as the ostrich was deemed fit for slaughter. Such an asymptomatic adult bird may become a source of infection to the remainder of the flock (Bowes 1993; Doneley et al. 1999). Similarly, other ratite species (Bowes 1993; Garcia et al. 2001; Hood 1977; Pocknell et al. 1996; Ševčíková et al. 1999; Shane et al. 1993) have presented with AM infections with apparently no visible loss in body condition (Pocknell et al. 1996). This is an important consideration when investigating unusual masses in apparently healthy ratite species.
Acknowledgements
Tiny Hlokwe from the Tuberculosis Laboratory, Zoonotic Diseases Section, Onderstepoort Veterinary Institute, South Africa; Mrs Charmaine Vermeulen for the macroscopic photography; the Department of Paraclinical Sciences, Faculty of Veterinary Science, University of Pretoria for funding the histopathology and special stains. Publication of this article was sponsored by the Faculty of Veterinary Science, University of Pretoria.
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Authors' contributions
M.R.C. (University of Pretoria) located the specimen and drafted the manuscript. S.J.C. (University of Pretoria) carried out the histophathology and IHC preparation, staining, analysis and interpretation. J.T.S. (University of Pretoria) and M.R.C. assisted in the interpretation of the material. J.T.S. supervised the study, participated in the design and was responsible for the final editing. All authors read and approved the final manuscript.
References
Allen, J.L., 1992, 'A modified Ziehl-Neelsen stain for mycobacteria', Medical Laboratory Sciences 49, 99-102. PMid:1283196 [ Links ]
Anon, 1985, 'Tuberkulose onder volstruise', Maand Verslag, RSA Departement van Veterinêre Dienste, Wes Kaap. [ Links ]
Bonga Tomlinson, C.A., 2000, 'Feeding in paleognathus birds', in K. Schwenk (ed.), Feeding: Form, function, and evolution in tetrapod vertebrates, pp. 359-394, Academic Press, San Diego. [ Links ]
Bowes, V., 1993, 'Avian tuberculosis in ostriches', Canadian Veterinary Journal 34, 758. PMid:17424343 PMCid:1686642 [ Links ]
Casteleyn, C., Doom, M., Lambrechts, E., Van den Broeck, W., Simoens, P. & Cornillie, P., 2010, 'Locations of gut-associated lymphoid tissue in the 3-month-old chicken: A review', Avian Pathology 39, 143-150. http://dx.doi.org/10.1080/03079451003786105, PMid:20544418 [ Links ]
Cooper, R.G., 2005, 'Bacterial, fungal and parasitic infections in the ostrich (Struthio camelus var. domesticus), Animal Science Journal 76, 97-106. http://dx.doi.org/10.1111/j.1740-0929.2005.00243.x [ Links ]
Crole, M.R. & Soley, J.T., 2010, 'Gross morphology of the intra-oral rhamphotheca, oropharynx and proximal oesophagus of the emu (Dromaius novaehollandiae), Anatomia Histologia Embryologia 39, 207-218. http://dx.doi.org/10.1111/j.1439-0264.2010.00998.x, PMid:20353488 [ Links ]
Crole, M.R. & Soley, J.T., 2012a, 'Evidence of a true pharyngeal tonsil in birds: A novel lymphoid organ in Dromaius novaehollandiae and Struthio camelus (Palaeognathae)', Frontiers in Zoology 9, 21. http://dx.doi.org/10.1186/1742-9994-9-21, PMid:22909013 PMCid:3502113 [ Links ]
Crole, M.R. & Soley, J.T., 2012b, 'What prevents Struthio camelus and Dromaius novaehollandiae (Palaeognathae) from choking? A novel anatomical mechanism in ratites, the linguo-laryngeal apparatus', Frontiers in Zoology 9, 11. http://dx.doi.org/10.1186/1742-9994-9-11, PMid:22650266 PMCid:3404984 [ Links ]
Cromie, R.L., Ash, N.J., Brown, M.J. & Stanford, J.L., 2000, 'Avian immune responses to Mycobacterium avium: The wildfowl example', Developmental and Comparative Immunology 24, 169-185. http://dx.doi.org/10.1016/S0145-305X(99)00071-3 [ Links ]
Davis, G.B., Watson, P.R. & Billing, A.E., 1984, 'Tuberculosis in a kiwi (Apteryx mantelli)', New Zealand Veterinary Journal 32, 30. http://dx.doi.org/10.1080/00480169.1984.35052, PMid:16031035 [ Links ]
Doneley, R.J.T., Gibson, J.A., Thorne, D. & Cousins, D.V., 1999, 'Mycobacterial infection in an ostrich', Australian Veterinary Journal 77, 368-370. http://dx.doi.org/10.1111/j.1751-0813.1999.tb10305.x, PMid:10812400 [ Links ]
Fitzgerald, S.D. & Moisan, P.G., 1995, 'Mycotic rhinitis in an ostrich', Avian Diseases 39, 194-196. http://dx.doi.org/10.2307/1592003, PMid:7794183 [ Links ]
Garcia, A., LeClear, C.T. & Gaskin, J.M., 2001, 'Mycobacterium avium infection in an ostrich (Struthio camelus), Journal of Zoo and Wildlife Medicine 32, 96-100. PMid:12790402 [ Links ]
Gerlach, H., 1994, 'Bacteria', in B.W. Ritchie, G.J. Harrison & L.R. Harrison (eds.), Avian medicine: Principles and application, pp. 971-975, Wingers Publishing, Lake Worth. [ Links ]
Gürel, A., Gülcubuk, A. & Turan, N., 2004, 'A granulomatous conjunctivitis associated with Morexella phenylpyruvica in an ostrich (Struthio camelus)', Avian Pathology 33, 196-199. http://dx.doi.org/10.1080/03079450310001652095, PMid:15276987 [ Links ]
Gussekloo, S.W.S. & Bout, G.R., 2005, 'The kinematics of feeding and drinking in palaeognathous birds in relation to cranial morphology', Journal of Experimental Biology 208, 3395-3407. http://dx.doi.org/10.1242/jeb.01769, PMid:16109899 [ Links ]
Gyimesi, Z.S., Stalin, I.H., Miller, J.M. & Thoen, C.O., 1999, 'Detection of Mycobacterium avium subspecies avium in formalin-fixed, paraffin-embedded tissues of captive exotic birds using polymerase chain reaction', Journal of Zoo and Wildlife Medicine 30, 348-353. PMid:10572856 [ Links ]
Hood, H.B., 1977, 'Eye pathology in an adult male ostrich (Struthio camelus)', Proceedings of the American Association of Zoo Veterinarians 17, 54-58. [ Links ]
Huchzermeyer, F.W., 1997, 'Public health risks of ostrich and crocodile meat', Revue Scientifique et Technique, Office International des Épizooties 16, 599-604. [ Links ]
Kazda, J., Pavlik, I. & Hruska, K., 2009, 'Chapter 6. The occurrence of pathogenic and potentially pathogenic mycobacteria in animals and the role of the environment in the spread of infection', in J. Kazda, I. Pavlik, J.O. Falkinham III & K. Hruska, (eds.), The ecology of mycobacteria: Impact on animal's and human's health, pp. 199-281, Springer, Dordrecht. http://dx.doi.org/10.1007/978-1-4020-9413-2 [ Links ]
Kincade, G.F., 1959, 'Avian tuberculosis in the human', Canadian Medical Association Journal 80, 32-34. [ Links ]
Krishna Mohan Roa, M.R. & Chowdary, C., 1980, 'Tuberculosis in an emu (Dromiceius novoeholandies)', Indian Veterinary Journal 57, 169. [ Links ]
Liebler-Tenorio, E.M. & Reinhard, P., 2006, 'MALT structure and function in farm animals', Veterinary Research 37, 257-280. http://dx.doi.org/10.1051/vetres:2006001, PMid:16611547 [ Links ]
Levkut, M., Ševčíková, Z., Ledecký, V. & Pauer, T., 1998, 'Neobvykla manifestacia tuberkulozy u pstrosa dvojprsteho', Veterinarstvi 48, 380. [ Links ]
McManus, J.F.A., 1946, 'Histological demonstration of mucin after periodic acid', Nature (London) 158, 202. http://dx.doi.org/10.1038/158202a0, PMid:20995486 [ Links ]
Miller, J., Jenny, A., Rhyan, J., Saari, D. & Suarez, D., 1997, 'Detection of Mycobacterium bovis in formalin-fixed, paraffin-embedded tissues of cattle and elk by PCR amplification of an IS6110 sequence specific for Mycobacterium tuberculosis complex organisms', Journal of Veterinary Diagnostic Investigation 9, 244-249. http://dx.doi.org/10.1177/104063879700900304, PMid:9249162 [ Links ]
Momotani, E., Kiryu, M., Ohshiro, M., Murakami, M., Ashida, Y., Watanabe, S. & Matsubara, Y., 1995, 'Granulomatous lesions caused by Pseudomonas aeruginosa in the ostrich (Struthio camelus)', Journal of Comparative Pathology 112, 273282. http://dx.doi.org/10.1016/S0021-9975(05)80080-5 [ Links ]
Montali, R.J., Bush, M., Thoen, C.O. & Smith, E., 1976, 'Tuberculosis in captive exotic birds', Journal of the American Veterinary Medical Association 169, 920-927. PMid:977462 [ Links ]
Nouvel, J. & Leclerc-Cassan, M., 1970, 'Un cas de tuberculose du penis chez l'autruche', Recueil de Médecine Vétérinaire 7, 879-880. [ Links ]
Pavlik, I., Machackova, M., Yayo Ayele, W., Lamka, J., Parmova, I., Melicharek, I., et al.,2002, 'Incidence of bovine tuberculosis in wild and domestic animals other than cattle in six Central European countries during 1990-1999', Veterinarni Medicina 47, 122-131. [ Links ]
Pavlik, I., Svastova, P., Bartl, J., Dvorska, L. & Rychlik, I., 2000, 'Relationship between IS901 in the Mycobacterium avium Complex strains isolated from birds, animals, humans, and the environment and virulence for poultry', Clinical and Diagnostic Laboratory Immunology 7, 212-217. PMid:10702495 PMCid:95851 [ Links ]
Pocknell, A.M., Miller, B.J., Neufeld, J.L. & Grahn, B.H., 1996, 'Conjunctival mycobacteriosis in two emus (Dromaius novaehollandiae)', Veterinary Pathology 33, 346-348. http://dx.doi.org/10.1177/030098589603300314, PMid:8740711 [ Links ]
Porchescu, G., 2007, 'Comparative morphology of the digestive tract of the black African ostrich, hen and turkey', PhD thesis, Ministry of Agriculture and Food Industry of the Republic of Moldova, Moldova Agrarian State University. [ Links ]
Ryo, I., Hiroaki, K. & Yoshihiro, W., 2006, 'An outbreak of mycobacterial infection in a farm of ostriches and efficacy of fecal cultivation as an ante mortem diagnosis', Journal of Livestock Medicine 520, 607-611. [ Links ]
Sanford, S.E., Rehmtulla, A.J. & Josephson, G.K.A., 1994, 'Tuberculosis in farmed rheas (Rhea americana), Avian Diseases 38, 193-196. http://dx.doi.org/10.2307/1591857, PMid:8002893 [ Links ]
Saroglu, M., Yucel, R. & Aktas, M., 2003, 'Granulomatous conjunctivitis in an ostrich', Veterinary Opthalmology 6, 337-339. http://dx.doi.org/10.1111/j.1463-5224.2003.00305.x, PMid:14641833 [ Links ]
Ševčíková, Z., Ledecký, V., Capík, I. & Levkut, M., 1999, 'Unusual manifestation of tuberculosis in an ostrich (Struthio camelus)', Veterinary Record 145, 708. PMid:10638801
Shane, S.M., Camus, A., Strain, M.G., Thoen, C.O. & Tully, T.N., 1993, 'Tuberculosis in commercial emus (Dromaius novaehollandiae)', Avian Diseases 37, 1172-1176. http://dx.doi.org/10.2307/1591933, PMid:8141752 [ Links ]
Shivaprasad, H.L., 2002, Pathology of birds - An overview, AFIP, Washington. PMCid:139663 [ Links ]
Tacconi, G. & Valente, C., 1981, 'Su di un caso di tubercolosi del nandu (Rhea americana)', Rivista di zootecnia e veterinaria 9, 234-236. [ Links ]
Tadjalli, M., Mansouri, S.H. & Poostpasand, A., 2008, 'Gross anatomy of the oropharyngeal cavity in the ostrich (Struthio camelus)', Iranian Journal of Veterinary Research 9, 316-323. [ Links ]
Tell, L.A., Woods, L. & Cromie, R.L., 2001, 'Mycobacteriosis in birds', Revue Scientifique et Technique, Office International des Épizooties 20, 180-203. [ Links ]
Thornton, C.G., Cranfield, M.R., MacLellan, K.M., Brink, T.L., Strandberg, J.D., Carlin, E.A., et al., 1999, 'Processing postmortem specimens with Cl8-carboxypropylbetaine and analysis by PCR to develop an antemortem test for Mycobaterium avium infections in ducks', Journal of Zoo and Wildlife Medicine 30, 11-24. PMid:10367639 [ Links ]
Tivane, C., 2008, 'A morphological study of the oropharynx and oesophagus of the ostrich (Struthio camelus), MSc dissertation, Anatomy and Physiology Department, University of Pretoria. [ Links ]
Tully, T.N. & Shane, S.M., 1996, 'Husbandry practices as related to infectious and parasitic diseases of farmed ratites', Revue Scientifique et Technique, Office International des Épizooties 15, 73-89. [ Links ]
Ulrichs, T., Lefmann, M., Reich, M., Morawietz, L., Roth, A., Brinkmann, V., et al., 2005, 'Modified immunohistological staining allows detection of Ziehl-Neelsen-negative Mycobacterium tuberculosis organisms and their precise localization in human tissue', Journal of Pathology 205, 633-640. http://dx.doi.org/10.1002/path.1728, PMid:15776475 [ Links ]
Wozikowski, R., 2001, 'Gruzlica u fermowych strusi afrykanskich' [Control of enzootic bovine, bovine tuberculosis, brucellosis in cattle], Życie Weterynaryjne 76, 664-665. [ Links ]
 Correspondence:
Correspondence:
Martina Crole
Private Bag X04
Onderstepoort 0110, South Africa
Email: martina.crole@up.ac.za
Received: 20 Nov. 2012
Accepted: 19 Apr. 2013
Published: 30 Oct. 2013

