










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkJournal of the South African Veterinary Association
versión On-line ISSN 2224-9435
versión impresa ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.84 no.1 Pretoria ene. 2013
ORIGINAL RESEARCH
Effects of the number of people on efficient capture and sample collection: A lion case study
Sam M. FerreiraI; Nkabeng T. MarupingII; Darius SchoultzIII; Travis R. SmitIII
ISANParks Scientific Services, Skukuza, South Africa
IIDepartment of Nature Conservation, Tshwane University of Technology, South Africa
IIIAjubatus Foundation, Hoedspruit Wildlife Estate, South Africa
ABSTRACT
Certain carnivore research projects and approaches depend on successful capture of individuals of interest. The number of people present at a capture site may determine success of a capture. In this study 36 lion capture cases in the Kruger National Park were used to evaluate whether the number of people present at a capture site influenced lion response rates and whether the number of people at a sampling site influenced the time it took to process the collected samples. The analyses suggest that when nine or fewer people were present, lions appeared faster at a call-up locality compared with when there were more than nine people. The number of people, however, did not influence the time it took to process the lions. It is proposed that efficient lion capturing should spatially separate capture and processing sites and minimise the number of people at a capture site.
Introduction
Large carnivore conservationists often face challenges that require the capture of individual animals. Response to transgressing animals occasionally includes capture and translocation to other areas (Trinkel et al. 2008). In addition, many research questions aim to understand large carnivore biology through conducting population surveys (Ferreira & Funston 2010; Smuts, Whyte & Dearlove 1977), defining demographic profiles (Ferreira & Funston 2010; Funston, Hermann & Van Vuuren 2005), defining spatial use (Ogutu & Dublin 2002) and conducting disease screening (Biek, Drummond & Poss 2006). Such approaches generally consist of three components: finding or attracting individuals, capturing them by various means and collecting and processing sample material.
Existing techniques that assist conservationists in attracting or finding individuals include baited cage traps (Bizzarri, Lacrimini & Ragni 2010), snare or leg hold traps (Frank, Simpson & Woodroffe 2003), baited call-ups (Castley et al. 2002), tracking (Stander 1998) and ambushing target animals at sites that individuals often visit (Marnewick & Cilliers 2006). In many cases, individuals need to be chemically immobilised given the generally aggressive nature of large carnivores. In general, sedation and immobilisation techniques for large carnivores are well developed (e.g. Anderson & Richardson 1992; Stander & Morkel 1991), but the influence of environmental factors on induction and recovery is not well understood. Nevertheless, the act of capturing individuals may lead to variances in immobilisation efficiency (i.e. the time taken for an individual to become completely sedated). In addition, factors influencing sample collection from a captured individual may also lead to the unsuccessful release of an animal, such as an individual awakening earlier than expected or dying whilst immobilised.
Capturing lions (Panthera leo) in the Kruger National Park (KNP) served as a case study to evaluate some factors that may influence efficacy of capture or sample collection. Lions in the KNP, which is part of the Great Limpopo Transfrontier Park, are a key conservation unit (Chardonnet 2002). Although the population is relatively large (1684 individuals with a 95% confidence interval of 1617-1751, Ferreira & Funston 2010), threats such as poaching, droughts and diseases may significantly alter the lion population. A long-term population study directed at defining lion temporal and spatial dynamics (SANParks, unpublished records) depends on several captures and provides the opportunity to evaluate capture and sample processing efficiency.
Capturing lions in the KNP makes use of mass capture techniques (Smuts et al. 1977), which comprise a bait station setup and the play back sound of prey in distress. Response rates depend on the effective area that such a call-up setup will sample, the probability that a lion group within that area will respond and the probability that an individual in that group will respond. In the KNP, a call-up sample area of ~ 58 km2 was estimated, within which ~ 73% and ~ 29% of groups without and with cubs respond, respectively. In addition, ~ 96% and ~ 91% of individuals in responding groups without and with cubs, respectively, will respond to a call- up (Ferreira & Funston 2010). However, additional factors may influence response rates, including the terrain where call-ups take place and factors associated with disturbances such as the number of people present at a call-up site. The people assisting at a capture require expertise and a level of efficiency when processing captured individuals and collecting samples. Handling efficiency is dependent on how many lions are captured, availability of equipment and the overall number of experienced capture personnel present. The number of people present at a capture site may thus have significant effects on capture efficiency and specimen processing time. This study investigated (1) how the number of people present at a capture site influences lion response rates and (2) how the number of people at a sampling site influences the time taken for sample collection. Data from captures were assessed to address these two questions and provide recommendations directed at improving the capture of lions.
Materials and methods
Study area
The study took place in the KNP, South Africa, with an area of 19 485 km2. Mozambique and Zimbabwe abut the KNP in the east and north, respectively. The KNP is located in the low-lying savannas of the eastern parts of the Limpopo and Mpumalanga provinces, receiving annual rainfall ranging from 450 mm in the north to 750 mm in the south between October and March (Gertenbach 1980; Mucina & Rutherford 2006). The geology dichotomises into granite and gneiss soils in the west and nutrient rich basalt soils in the east of the park (Schutte 1986). The southern basalts are covered largely by wooded savanna, with Sclerocarya caffra and Acacia nigrescens dominating the tree canopy. Mixed Combretum spp. and Acacia spp. dominate the southern granites. Colophospermum mopane dominates both the basalt and granite substrates in the north (Gertenbach 1983; Mucina & Rutherford 2006).
Data collection
A total of 145 individual lions were included in the study, of which 85 were captured in the southern parts of the KNP (n = 17 capture occasions) between February 2010 and November 2010, and 60 in the north (n = 19) during August 2010. Call-ups were conducted at different times of the year depending on staff availability. In addition, the northern and southern parts of the KNP were sampled as independent groups because of varying disease prevalences, landscapes and prey biomass (Ferreira & Funston 2010). Locations of call-ups were in areas with optimal sound dispersal; that is, elevated and away from rivers.
Captures typically involved setting up a call-up station in close proximity to where lions were noted during the day. This removed the variance in responses associated with distance (see Ferreira & Funston 2010). Information from boards recording sightings by tourists, information from rangers and having members of the capture team drive out during the day to locate individuals assisted in gathering information about a potential pride for capture. Obtaining knowledge on pride size beforehand is ideal, but on most occasions it was unrealistic owing to logistics and time constraints. Therefore, using information after captures helped to identify pride size and therefore the response rate of the whole pride. Fresh buffalo (Syncerus caffer), zebra (Equus zebra), wildebeest (Connochaetes taurinus) or impala (Aepyceros melampus) carcasses secured to a tree or pole driven into the ground were used as bait. Bait selection was determined by availability and known prey preference in the region. Larger prey species were selected to allow for longer feeding periods and thereby increase the time available to dart lions.
A recording of a combination of calls consisting of a buffalo calf in distress and pig squeals were played repeatedly for varying periods of time on an LG MF-FM12 MP3 player (LG Electronics, Korea). Sound was amplified through a 12 V, 60 W Jedia Mobile 60 Mixing Power Amplifier (Jedia, Korea) connected to two Show 40 M 4 Ω horn speakers (Show Co., Korea; diameter = 40 cm) with Show TU-35 M4 40 W driver units (Show Co., Korea) connected in series and facing in a 180° direction from each other. The vocalisations were broadcast at full volume from the speakers mounted on a steel platform (height = 2.5 m) that was pegged to the ground. Call-ups started 30 min after sunset and were completed before midnight.
A caravan was parked 20 m from the bait and a light delivery vehicle with an open load bed was used as recovery vehicle after lions were immobilised. In some instances only the light delivery vehicle was present, with the speaker system mounted on the roof. This enhanced mobility when several localities needed to be targeted in one night. For each call-up the numbers of motorised and non-motorised (caravans and trailers) vehicles, as well as people present at the site were recorded. The time from the start of a call-up until lions appeared and the number of lions that responded were then noted. The vehicles present were parked in a circle, away from the bait. A game guard or ranger with a rifle was always present and a spotlight was used to scan the area at approximately 10 min intervals.
Prior to the start of a call-up, the capture coordinator would instruct all those present as to the protocol during a capture and give directions to people when tasks needed to be carried out during the capture. Capture operations were predominantly carried out by experienced personnel who were aware of the capture protocol. On numerous occasions, however, inexperienced guests were allowed to join the capture, which increased the number of people and the number of vehicles at a capture site. This increase in the total number of people and vehicles could potentially have affected response rates.
In addition, the presence of additional people limited processing space within the circle of vehicles and increased visibility of humans and noise. People involved in the capture were requested either to remain in the vehicles or to be within the protective circle of vehicles. The use of unnecessary lights was prohibited and voices had to be kept to a minimum. The role of participants was to set up the bait, immobilise and retrieve the lions, gather samples from the caught individuals and scan the surrounding area with spotlights as a safety precaution.
Once lions were feeding on the bait, veterinarians used formalised drug combinations (SANParks, unpubished data) according to an individual's age and sex for darting. Most lions were darted using a combination of medetomidine and Zoletil (a combination of tiletamine and zolazepam) delivered from the caravan or vehicle with a Dan-Inject dart gun. Lions were darted and processed simultaneously. Processing involves a team of four people retrieving an animal from the bait and transferring it to the processing site within the protective vehicle circle using the light delivery vehicle.
Sample collection varied depending on the objectives of the study for which a capture was undertaken. For each lion, veterinarians collected blood samples, performed body measurements, conducted body assessments, assigned ages (Whitman & Packer 2006) and collected tissue samples to define disease status. In addition, researchers fitted one standard mammalian tracking collar on selected prides (n = 20 across the KNP). Efficiency of these tasks can vary as a result of the number of experienced people assisting. The lions were then taken back to the capture site near the bait and the anaesthetic was reversed following antidote application of atipamezole (Antisedan). The time was noted when a lion was first sedated with the combination drugs from the dart gun, as well as when it had fully recovered and was mobile after sedation had been reversed. The number of people present was also noted. During recovery all people present remained within the protective car circle or in vehicles and did not have direct contact with the animals.
Data analyses
It was argued that a response index, independent of distance effects (Ferreira & Funston 2010), is a function of the number of lions responding and the response time. In a group lions tend to be more adventurous in exploiting potential food resources than as individuals (Funston, Mills & Biggs 2001). A standardised response index (It) was calculated according to:

Where n is the number of lions responding at call-up, i is the call-up and t is the response time of those lions to call-up. T denotes total. This index scales between 0 and 1.
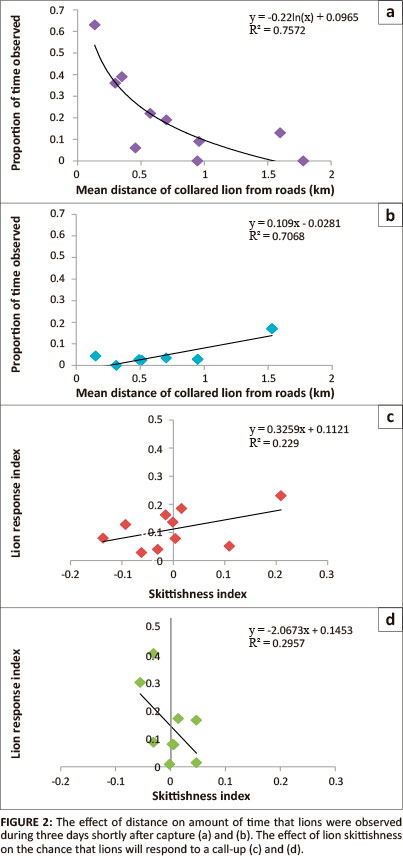
Behavioural differences between lions were determined by previous type and frequency of exposures to human activity. Lions habituate easily to people and vehicles when regularly exposed to tourist vehicles and open game drive vehicles (Okello, Manka & D'Amour 2008). Exposure to vehicles is likely to vary across the KNP because of tourist road densities and the number of tourists. Each pride was followed continuously for three days within two months of a collar being placed on a pride. Visibility as well as the time the animal was seen was noted and the percentage of times seen was used as an index of skittishness. Such an index is sensitive to the distance of a pride from roads or vehicle access. Distance was measured using MapWindowGIS, where distances were measured from fixed global positioning system points along tourist tar roads. It was thus necessary to correct the skittishness index for the mean distance a pride was away from roads as an index of the influence of vehicle access, vegetation density and terrain during the three days of them being followed. The skittishness index was plotted against mean distance from tourist tar roads, a linear regression was fitted (Sokal & Rohlf 1995) and residuals were derived from this regression if a significant relationship existed between these variables.
It is acknowledged that skittishness as defined here could be influenced by how individual lions experienced stress and compromised well-being in response to people at captures. However, post-capture observation of individual lion interactions suggested that it was unlikely and no capture-related mortalities or aberrant behaviour was recorded.
To define how people may influence the response rate of lions the data were arranged sequentially, from smallest to greatest number of people present (ni). Paired response curves were then constructed progressively, one for call-ups when ni< n*,j, and one for call-ups when ni > n*,j, where n*,j was a critical splitting value defined by the number of people present (people split). A response curve was defined by calculating, at 5 min intervals, the proportion of call-ups where lions responded in the sample classified by n*,j. We then used a one tailed paired t-test (Sokal & Rohlf 1995) to evaluate whether the response curve for ni< n*,j predicted a faster response than that for ni> n*, j. By progressively increasing n*,j a value n* was defined until the two response curves were no longer different; that is, where ni< n*,j = ni > n*,j. We plotted a probabilities from paired t-tests against the people split and fitted the function log y = β1 + β2log x to this data using linear regression (Sokal & Rohlf 1995).
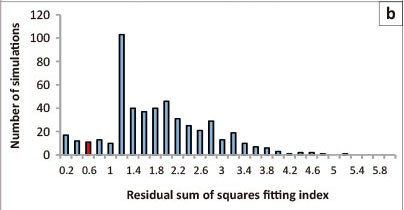
There was concern that the results obtained might be a common consequence of such an analytical approach primarily because it was noticed that there was a differential response in the initial data analyses. Best fit for a probabilities from paired t-tests against the people split was achieved when n*,j < 15 and β2> 0, whilst n*,j> 15 and β < 0. We thus randomly re-arranged the response time data amongst the number of people present and recalculated response curves and paired t-tests for the equivalent series of n*,j values in the same way as before. The residual sum of squares was then used to which 1 was added if n*,j < 15 and β2 < 0 and 1 again if n*,j> 15 and β2 > 0. We termed this the residual sum of square fitting index (RSSfi), which reflects whether similar types of models fitted to observed data did better when fitted to rearranged data. To check this we repeated this simulation 500 times and constructed a frequency distribution of RSSfi. It was concluded that the observed pattern is most likely unique at the 90% confidence level if it falls within the lower 10% of the distribution of RSSfi.
To evaluate the second study question, the average time that lions were immobilised at each capture was used as an index of processing time (tp) to collect samples. It was anticipated that the more lions captured, the longer the processing time would be. We thus fitted a multivariate linear regression model that included the number of lions captured at occasion i (Li) as well as the number of people present (ni) (Sokal & Rohlf 1995). The t-statistics for the coefficients estimated for each variable were used to evaluate the relative ability of a variable to explain the variance we noted in processing time.
Ethical considerations
All protocols to ensure the safety of the people involved at the captures was undertaken by the Veterinary Wildlife Services of South African National Parks (SANParks). The welfare and safety of the animals during capture events were also the responsibility of the Veterinary Wildlife Services (standard operating procedures for the capture, transportation and maintenance in holding facilities of wildlife).
Captures were undertaken by staff members, researchers, employed affiliates of the project and some observers. Safety procedures were discussed prior to all capture events by a Veterinary Wildlife Services employee. All researchers, employed affiliates and observers completed and submitted official indemnity forms.
Only personnel affiliated with or approved by SANParks participated in data collection. Once collected, the data remained with the researchers and the employed affiliates.
Results
The time until lions appeared at a call-up station varied from 2 min to 8 min, with between 1 and 13 lions arriving. There were between 1 and 33 people, and between 1 and 9 motorised and non-motorised vehicles at call-up stations. On average, more lions (t27 = 2.19, p = 0.04) appeared in the south after a call-up started (5.41 ± 3.71; n = 17) than in the north (3.11 ± 2.38; n = 19), but they did so in relatively similar times (south: 36.11 min ± 33.80 min, n = 17; north: 27.05 min ± 19.33 min, n = 19; t27 = 0.93, p = 0.36).
Exploratory analyses suggested that vehicles were not a factor. Response indices were bimodal in relation to the number of people at a call-up station (Figure 1). When the northern and southern sites were considered separately we noted differences. The response index was 0.12 ± 0.09 (mean ± s.d., n = 16) in the south, which was significantly higher than the 0.05 ± 0.05 (n = 19) noted in the north of the park (t34 = 2.44, p = 0.02). It was concluded that a better lion response could be expected when the number of people present at a call-up is smaller than n, predicted at α = 0.05.

The time that lions were visible during three continuous days of observation shortly after initial capture was significantly influenced by the distance they were away from tourist tar roads, both in southern (t8 = 7.36, p < 0.01) and northern KNP (t8 = 4.11, p < 0.01), as shown in Figure 2. When the effect of distance was removed by calculating residuals, the negative of which reflects skittishness, lion response rates were only weakly associated with skittishness in both the southern (t8 = 1.54, p = 0.16) and the northern (tg = 1.72, p = 0.13) parts of the KNP. In the south, lions responded better when the skittishness index increased, whilst the opposite was noted in the north.
Paired t-test probabilities for paired lion response curves to the number of people split at a series of n*,j values predicted that for fewer than nine people the response of lions was quicker than if there were more people (Figure 3a). However, it also predicted that when 14 or more people were present the response of lions was also quicker. This pattern was relatively unique as only 8.7% of the simulated RSSfi values were smaller than those that were noted for the observed data in this study (Figure 3b). In this case, all call-ups had lions responding within 60 min if fewer than nine people were present (n = 14). Conversely, after 110 min 93% of call- ups had lions responding when nine or more people were present (n = 21) (Figure 3c).


We could only weakly explain processing time with a multivariate model including the number of lions and the number of people (tp = 1.18 n1 + 4.11 Li + 47.67, R2= 0.21, F2,23 = 3.12, p = 0.06). Most of the variance that could be explained was associated with the number of lions captured (t23 = 2.02, p = 0.05), rather than with the number of people present (t23 = 1.34, p = 0.19). The average number of people present at captures differed significantly (t35 = 3.73, p < 0.01), with the number in the south (16.88 ± 7.02, n = 17) being nearly double that in the north (8.74 ± 6.04, n = 19). Processing time was different between the northern and the southern sites (t26 = 5.77, p < 0.01), with processing taking longer in the south (108.93 min ± 22.00 min, n = 60) than in the north (62.67 min ± 34.35 min, n = 21).
Discussion
Capturing large carnivores presents challenges (Linnell et al. 1997), some of which can be controlled by capture teams. It was noted in this study that the number of people present affects lion responses to a call-up at a capture site, but not the time it took to extract samples and data from immobilised lions. Some anomalies were also noted. Response indices that accommodated response times as well as the number of lions responding was higher in the southern parts of the KNP than noted for the northern KNP. The number of people present at capture sites was, however, significantly greater in the south compared with the north.
A primary concern is that the assumption of negligible influences of landscape features (Ferreira & Funston 2010) and intra- as well as interspecific competition (Owen-Smith & Mills 2008) on lion response were not fully accounted for. Of the 36 incidences of captures included in this study, only 5 involved cases of no confirmed lion sightings before the call-up site was selected. All of these were in the northern part of the park and non-responses were recorded in only two of these cases. The distance capacity of the call-up station is 4.5 km. Although wind and landscape were not accounted for in the analysis, we are confident that the variable response rates associated with the distance of the lions from a capture site at the start of a call-up (Ferreira & Funston 2010) had a negligible influence on the results. Influences of other species responding to call-ups, specifically spotted hyaenas (Crocuta crocuta) (Graf et al. 2009), was not formally assessed in this or other studies.
Intraspecific effects, namely response rates when other lions have already responded, is also anticipated to be negligible although it was noted previously that prides with cubs responded differently from those without cubs (Ferreira & Funston 2010). The approach of identifying lion sightings without cub information and focusing on prides when setting up call-up stations most likely negated this potential confounding factor.
Lions can be skittish and reluctant to respond to distress calls of prey for other reasons than those already noted. Most noteworthy would be experiences with humans, which may have been either non-detrimental (e.g. ecotourism experiences [Okello et al. 2008]) or harmful (e.g. conflict with humans, usually associated with livestock raiding [Inskip & Zimmermann 2009]). It was noted that weak relationships between lion response indices and indices of skittishness were present. These, however, were different between the northern (decreased response index with increased skittishness index) and the southern areas (increased response index with increased skittishness index). The northern part of the KNP has far fewer tourists visiting the park than the southern KNP (Kruger & Saayman 2010). In addition, human-lion conflict is almost solely restricted to the north-western boundary of the KNP. Lions most likely do not associate vehicles with danger related to humans, particularly when these involve lion friendly activities such as ecotourism (Okello et al. 2008). The reality is that although lions in the northern KNP may have different histories of experiences with humans than those in the southern KNP, skittishness may not be a confounding factor, simply because relationships were weak at best.
A final confounding factor is associated with lion densities and, perhaps, social structure. In the south, lions live at densities ranging from 6.9 n.100 km-2 to 14.9 n.100 km-2 compared with 4.1 n.100 km-2 to 9.7 n. 100 km-2 in the north (Ferreira & Funston 2010). More lions responded per call-up in the southern parts of the KNP than in the northern KNP. The number of lions responding per call-up in the south was ~ 70% higher than in the north; densities are ~ 60% higher in the south than in the north. Responses per group thus reflect population densities to such an extent that densities do not impose a confounding factor on how lions responded to call-ups in this study.
It should be noted that the time it took for lions to appear at a call-up had a similar statistical distribution in the northern and southern KNP. The response indices may thus be confounded by factors influencing individual lions' responses, leading to the non-relationship noted between the response index and the number of people.
Data were pooled because response times were statistically similar in the northern and southern parts. When response curves were constructed by calculating, at five-minute intervals, the proportion of call-ups where lions responded in a sample of call-ups arranged from lowest to highest number of people present, the analyses revealed key splits in response curves when fewer than 10 people were present. One assumption is that the actual visibility of more people and vehicles could cause lions to be more tentative towards going to the bait. Lions appeared consistently faster at call-up stations with fewer than 10 people present than where there were more people. It was also noted that when more than 14 people were present lions' response times were faster.
The reason for this can be only speculative. It has already been pointed out that lions most likely do not associate danger from humans with humans in vehicles. There may, however, be an introduced element unrelated to skittishness associated with habituation to vehicles (Okello et al. 2008) or previous experience with humans (Inskip & Zimmermann 2009). Harmful experiences such as hunting or torment through human-wildlife conflict may lead to lions avoiding voices of people, but in cases when lions have killed and eaten humans, they may respond inquisitively to human voices. It is unlikely that lions, specifically in the southern KNP, are habituated to people as a result of their human-eating habits because incidences in the KNP are extremely low (~ 21 in 40 years, SANParks, unpublished data) compared with other places in Africa (Packer et al. 2005). Instead, it is proposed that lions are inquisitive and, when presented with novel or neutral elements in their environment in addition to the prey response induced by the call-up procedure, are likely to investigate. A large fraction of the relatively quick lion responses noted when there were a large number of people present was in the southern KNP, the most likely area for lions not to have experienced any regular detrimental interactions with humans.
These results suggest that the number of people present at a capture site influenced the rate at which lions responded. This effect, however, is not evident when considering the time needed for collecting samples and data from individual lions that were successfully captured and immobilised. In fact, in the south sampling and data collection time was longer when many people were present, but more lions were also caught per capture occasion. Processing time was associated with the number of lions caught rather than the number of people present, primarily because of expertise required for certain procedures and constraints associated with limited availability of specialised equipment.
Conclusion
The results illustrate that, (1) the number of people present at a capture site influenced lion response rates, but (2) that the number of people at a sampling site did not influence the time it took to extract samples and collect data from individual lions. This suggests that for research that requires the capture of large carnivores such as lions, researchers can achieve efficient capture and data collection and processing by separating capture sites from sites where samples and data are collected from immobilised individuals. It is suggested that the minimum number of people at a capture site is preferable. A maximum of nine people at a capture occasion, however, will optimise lion capture success and processing efficiency.
Acknowledgements
We would like to thank the Ajubatus Foundation for providing financial and logistical support. We are grateful to the Veterinary Wildlife Services of SANParks for assistance with capturing and logistical arrangements.
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this article.
Authors' contributions
S.M.F. (SANParks) is the large-mammal ecologist based in the KNP. As the project leader S.M.F. was responsible for project design, coordination and primary analysis. N.T.M (Tshwane University of Technology) collected, sorted and proofed the data supplied with some analysis. D.S. (Ajubatus Foundation) and T.R.S (Ajubatus Foundation) collected and provided field data.
References
Anderson, M.D. & Richardson, P.R.K., 1992, 'Immobilization of the aardwolf', South African Journal of Wildlife Research 22, 26-28. [ Links ]
Biek, R., Drummond, A.J. & Poss, M., 2006, 'A virus reveals population structure and recent demographic history of its carnivore host', Science 311, 538-541. http://dx.doi.org/10.1126/science.1121360, PMid:16439664 [ Links ]
Bizzarri, L., Lacrimini, M. & Ragni, B., 2010, 'Live capture and handling of the European wildcat in central Italy', Italian Journal of Mammalogy 21, 73-82. [ Links ]
Castley, J.G., Knight, M.H., Mills, M.G.L. & Thouless, C., 2002, 'Estimation of the lion (Panthera leo) population in the southeastern Kgalagadi Transfrontier Park using a capture-recapture survey', African Zoology 37, 27-34. [ Links ]
Chardonnet, P., 2002, Conservation of the African lion: Contribution to a status survey. International Foundation for the Conservation of Wildlife, Conservation Force, Paris. [ Links ]
Ferreira, S.M. & Funston, P.J., 2010, 'Estimating lion population variables: Prey and disease effects in Kruger National Park, South Africa', Wildlife Research 37, 194-206. http://dx.doi.org/10.1071/WR09030 [ Links ]
Frank, L., Simpson, D. & Woodroffe, R., 2003, 'Footsnares: an effective method for capturing African lions', Wildlife Society Bulletin 31, 309-314. [ Links ]
Funston, P.J., Herrmann, E. & Van Vuuren, J. H., 2005, 'Lions in the Kgalagadi Transfrontier Park: modelling the effect of human-caused mortality', International Transactions in Operational Research 12, 145-171. http://dx.doi.org/10.1111/j.1475-3995.2005.00495.x [ Links ]
Funston, P.J., Mills, M.G.L. & Biggs, H.C., 2001, Factors affecting the hunting success of male and female lions in the Kruger National Park, Journal of Zoology 253, 419-431. http://dx.doi.org/10.1017/S0952836901000395 [ Links ]
Gertenbach, W.P.D., 1980, 'Rainfall patterns in the Kruger National Park', Koedoe 23, 35-43. http://dx.doi.org/10.4102/koedoe.v23i1.634 [ Links ]
Gertenbach, W.P.D., 1983, 'Landscape of the Kruger National Park', Koedoe 26, 9-121. http://dx.doi.org/10.4102/koedoe.v26i1.591 [ Links ]
Graf, J.A., Somers, M.J., Szykman-Gunther, M. & Slowtow, R., 2009, 'Heterogeneity in the density of spotted hyaenas in Hluhluwe-iMfolozi Park, South Africa' Acta Theriologica 54, 333-343. http://dx.doi.org/10.4098/j.at.0001-7051.095.2008 [ Links ]
Inskip, C. & Zimmermann, A., 2009, 'Human-felid conflict: A review of patterns and priorities worldwide', Oryx 43, 18-34. http://dx.doi.org/10.1017/S003060530899030X [ Links ]
Kruger, M. & Saayman, M., 2010, 'Travel motivation of tourists to Kruger and Tsitsikamma National Parks: a comparative study', South African Journal of Wildlife Research 40, 93-102. http://dx.doi.org/10.3957/056.040.0106 [ Links ]
Linnell, J., Aanes, R., Swenson, J., Odden, J. & Smith, M., 1997, 'Translocation of carnivores as a method for managing problem animals: A review', Biodiversity Conservation 6, 1245-1257. http://dx.doi.org/10.1023/B:BIOC.0000034011.05412.cd [ Links ]
Marnewick, K. & Cilliers, D., 2006, 'Range use of two coalitions of male cheetahs, Acinonyx jubatus, in the Thabazimbi district of the Limpopo Province, South Africa', South African Journal of Wildlife Research 36, 147-151. [ Links ]
Mucina L. & Rutherford, M.C. (eds), 2006, The vegetation of South Africa, Lesotho and Swaziland, Strelitzia 19, 540-567. [ Links ]
Ogutu, J.O. & Dublin, H.T., 2002, 'Demography of lions in relation to prey and habitat in the Maasai Mara National Reserve, Kenya', African Journal of Ecology 40, 120129. http://dx.doi.org/10.1046/j.1365-2028.2002.00343.x [ Links ]
Okello, M.M., Manka, S.G. & D'Amour, D.E., 2008, 'The relative importance of large mammal species for tourism in Amboseli National Park, Kenya', Tourism Management 29, 751-760. http://dx.doi.org/10.1016/j.tourman.2007.08.003 [ Links ]
Owen-Smith, N, & Mills, M.G.L., 2008, 'Predator-prey size relationships in an African large-mammal food web', Journal of Animal Ecology 77, 173-183. http://dx.doi.org/10.1111/j.1365-2656.2007.01314.x, PMid:18177336 [ Links ]
Packer, C., Ikanda, D., Kissui, B. & Kushnir, H., 2005, 'Lion attacks on humans in Tanzania - understanding the timing and distribution of attacks on rural communities will help to prevent them', Nature 436, 927-928. http://dx.doi.org/10.1038/436927a, PMid:16107828 [ Links ]
Schutte, I. C., 1986, 'The general geology of the Kruger National Park', Koedoe 29, 13-38. http://dx.doi.org/10.4102/koedoe.v29i1.517 [ Links ]
Smuts, G.L., Whyte, I.J. & Dearlove, T.W., 1977, 'A mass capture technique for lions', East African Wildlife Journal 15, 81-87. [ Links ]
Sokal, R.R. & Rohlf, F.J., 1995, Biometry: The principles and practice of statistics in biological research, W.H. Freeman, New York. [ Links ]
Stander, P.E., 1998, 'Spoor counts as indices of large carnivore population: the relationship between spoor frequency, sampling effort and true density', Journal of Applied Ecology 35, 378-385. http://dx.doi.org/10.1046/j.1365-2664.1998.00313.x [ Links ]
Stander, P.E. & Morkel, P., 1991, 'Field immobilization of lions using disassociative anaesthetics in combination with sedatives', African Journal of Ecology 29, 137-148. http://dx.doi.org/10.1111/j.1365-2028.1991.tb00994.x [ Links ]
Trinkel, M., Ferguson, N., Reid, A., Somers, M., Turelli, L., Graf, J. et al., 2008, 'Translocating lions into an inbred lion population in the Hluhluwe-iMfolozi Park, South Africa', Animal Conservation 11, 138-143. http://dx.doi.org/10.1111/j.1469-1795.2008.00163.x [ Links ]
Whitman, K.L. & Packer, C., 2006, A practical guide to aging African lions, 1st edn., International Foundation for the Conservation of Wildlife, Conservation Force, Paris. [ Links ]
 Correspondence:
Correspondence:
Sam Ferreira
SANParks Scientific Services
Skukuza 1350, South Africa
sam.ferreira@sanparks.org
Received: 14 May 2012
Accepted: 02 Mar. 2013
Published: 24 May 2013

