










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkJournal of the South African Veterinary Association
versão On-line ISSN 2224-9435
versão impressa ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.83 no.1 Pretoria Jan. 2012
HISTORICAL ARTICLES
History of Orbivirus research in South Africa
Daniel W. Verwoerd
Faculty of Veterinary Science, University of Pretoria, South Africa
ABSTRACT
In the early colonial history of South Africa, horses played an important role, both in general transportation and in military operations. Frequent epidemics of African horsesickness (AHS) in the 18th century therefore severely affected the economy. The first scientific research on the disease was carried out by Alexander Edington (1892), the first government bacteriologist of the Cape Colony, who resolved the existing confusion that reigned and established its identity as a separate disease. Bluetongue (BT) was described for the first time by Duncan Hutcheon in 1880, although it was probably always endemic in wild ruminants and only became a problem when highly susceptible Merino sheep were introduced to the Cape in the late 18th century. The filterability of the AHS virus (AHSV) was demonstrated in 1900 by M'Fadyean in London, and that of the BT virus (BTV) in 1905 by Theiler at Onderstepoort, thus proving the viral nature of both agents. Theiler developed the first vaccines for both diseases at Onderstepoort. Both vaccines consisted of infective blood followed by hyper-immune serum, and were used for many years. Subsequent breakthroughs include the adaptation to propagation and attenuation in embryonated eggs in the case of BTV and in mouse brains for AHSV. This was followed by the discovery of multiple serotypes of both viruses, the transmission of both by Culicoides midges and their eventual replication in cell cultures. Molecular studies led to the discovery of the segmented double-stranded RNA genomes, thus proving their genetic relationship and leading to their classification in a genus called Orbivirus. Further work included the molecular cloning of the genes of all the serotypes of both viruses and clarification of their relationship to the viral proteins, which led to much improved diagnostic techniques and eventually to the development of a recombinant vaccine, which unfortunately has so far been unsuitable for mass production.
Introduction
Two diseases caused by orbiviruses have played an important role in the history of veterinary science in South Africa: African horsesickness (AHS) and bluetongue (BT) in sheep. Full descriptions of both diseases are available elsewhere (Coetzer & Tustin 2004), therefore only the historical milestones achieved in South African research will be discussed here.
In the early colonial history of South Africa, horses played an important role, both in general transportation and in military operations. Frequent epidemics of AHS therefore severely affected the economy. An early record, for example, states that 1700 horses, constituting a major proportion of the horse population, died in a single outbreak in the Cape Colony in 1719 (Theiler 1921). An outbreak in 1853 was one of the most severe, when 70 000 horses died, constituting more than 40% of the entire Cape of Good Hope horse population (Bayley 1856).
Alexander Edington, a medical graduate from Edinburgh who was appointed as the first government bacteriologist of the Cape Colony, initiated scientific research on AHS in 1891, and established its identity as a separate disease which previously had often been confused with anthrax and other conditions. Unfortunately, he rather rashly announced the following year (1892) that he had discovered the 'microbe' which caused the disease. Unable to substantiate this claim, he proposed to the Cape Parliament in 1895 that AHS was caused by a fungus, which he promptly named 'oedemamyces', and requested funding from the prime minister, for horses to prove his theory (Edington 1904). After failing again, he announced in 1898 (Edington 1900) that he had isolated an attenuated virus which could be used as a vaccine. When his results could again not be reproduced, his work unfortunately lost all credibility on the local scene although he was still regarded as an AHS 'expert' in the UK for many years, based on several publications there. His research activities were later limited by the Cape government, leading to his resignation in 1905.
Bluetongue was described for the first time by Duncan Hutcheon in 1880. Although probably always endemic in wild ruminants it only became a problem when highly susceptible Merino sheep were introduced to the Cape from Spain via the Netherlands in the late 18th century (Erasmus & Potgieter 2009). The first recorded research on BT was conducted by Spreull (1905).
Establishment of the viral nature of the aetiological agent
In 1900, the filterability of AHS virus (AHSV) was demonstrated by M'Fadyean in London, using material supplied by Robertson, a former assistant of Edington. Similar results were obtained for BTV in 1905 at Onderstepoort by Theiler, who transmitted the disease to sheep with a bacteria-free filtrate of infective blood. The viral nature of both agents was therefore established in terms of the technology available at the time. The problem of controlling both diseases remained unsolved, however, and AHS remained the main reason why the transport industry depended on oxen rather than horses for traction purposes until rinderpest appeared on the scene. The disease also played havoc amongst the British forces during the Anglo-Boer War, killing thousands of horses, which were mainly imported from British colonies free from the disease and therefore naïve in terms of disease resistance. After the eradication of rinderpest in 1898 and the conclusion of the Anglo-Boer War in 1902, AHS formed an early focus of Arnold Theiler's research and the high incidence of the disease on the farm De Onderstepoort was one of the reasons for its selection in 1908 as the site for a Veterinary Research Institute. Soon afterwards Theiler developed the first effective vaccines for both diseases. These consisted of the injection of infective blood followed by hyper-immune serum in the case of AHS and infective blood containing what he regarded as an 'attenuated' strain for BT. Both were used with reasonable success for several decades (Coetzer & Tustin 2004).
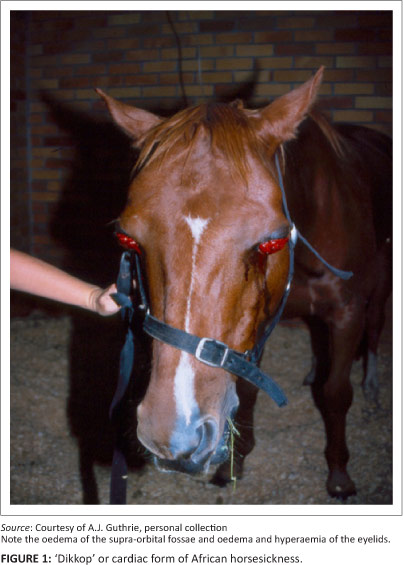
Multiplicity of serotypes
Theiler soon discovered what he called a 'plurality of immunologically distinct strains' of AHSV, based on the observation that immunisation with one isolate of the virus often did not protect against infection with another (Theiler 1921). This hypothesis was confirmed much later by Mcintosh (1958) and by Howell (1962) using a more stringent cross-neutralisation test in mice, leading to the identification of nine serotypes still recognised today.
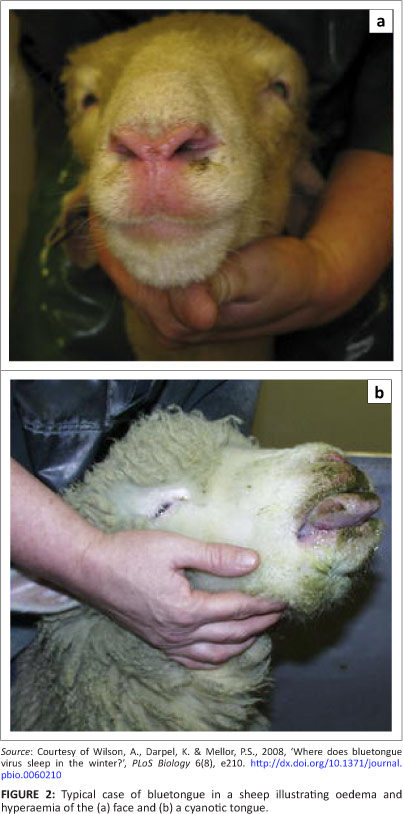
Incomplete protection by the single-strain BT vaccine introduced by Theiler was also observed, but it was only after BT virus (BTV) was adapted to growth in embryonated eggs (Mason, Coles & Alexander 1940), and after the discovery in 1947 that the optimal temperature for the cultivation of BTV in eggs is 33.5 °C and not 37 °C, that scientific evidence for multiple serotypes of this virus was obtained (Alexander 1947). A number of attenuated BTV strains were soon isolated and a trivalent, followed by a quadrivalent, live, attenuated vaccine was produced, replacing the old Theiler vaccine. By 1942, six serotypes had been identified and by the end of the 20th century this number had grown to 24. Immunological studies carried out in 1948 on the available BTV strains clearly demonstrated that immunisation with a particular strain resulted in lifelong solid immunity but little or no immunity to other strains (Neitz 1948). This explained the vaccine failures encountered with Theiler's monovalent AHS vaccine and formed the basis for later serotyping of isolates as well as the development of multivalent vaccines.
The discovery of multiple serotypes in orbiviruses thus played a critically important role in the development of effective vaccines.
Propagation and attenuation
A major milestone in Orbivirus research was the demonstration by Alexander in 1935 that AHSV can be propagated in mouse brains and that serial intra-cerebral passage in mice converts virulent viscerotropic isolates to a neurotropic attenuated virus, with retention of its immunogenic capacity. This work was based on that of Theiler's son Max in the United States of America, on the development of a yellow fever vaccine, for which he was awarded a Nobel Prize. In 1938, it was followed by the successful propagation and attenuation of AHSV in chicken embryos after chorio-allantoic inoculation (Alexander 1938).
During the 1930s, several attempts to repeat the mouse brain results with BTV failed, but in 1940 both growth and attenuation of the virus in embryonated hen's eggs succeeded (Mason et al. 1940). As mentioned above, these discoveries led to the development of new, more effective avianised vaccines for both diseases, containing several serotypes of neurotropic AHSV and chick-attenuated BTV respectively, which were successfully used for more than 30 years.
Discovery of the insect vector
The empirical knowledge that the losses due to both diseases could be limited by keeping animals indoors after sundown, and by avoiding wetland pastures, existed amongst generations of stock farmers, suggesting the involvement of biting insects in their transmission. The possibility of an insect vector for AHS was indeed suggested by Pitchford and Theiler in 1903. Yet it was only in 1944 that the development of a light trap for the catching of large numbers of these tiny insects facilitated the demonstration that both AHSV and BTV are transmitted by the tiny midge Culicoides pallidipennis (prev. imicola) (Du Toit 1944). Much later, other members of the Culicoides genus were also implicated, in various parts of the world. This discovery contributed significantly to the understanding of the epidemiology of both AHS and BTV. An outstanding example is the demonstration that the recent outbreaks of bluetongue in northern Europe followed changes in the distribution patterns of Culicoides caused by climate change.
Replication and vaccine production in cell cultures
The cultivation of egg-adapted BTV in lamb kidney cell cultures was another milestone (Haig, McKercher & Alexander 1956), leading to the discovery of 12 serotypes by means of in vitro neutralisation tests (Howell 1960) and the possibility of producing large volumes of vaccine in vitro, especially after switching to roller flasks of BHK21 cells in 1968. By this time, the number of serotypes in the vaccine had increased to 14 but the protection obtained was unsatisfactory due to interference between and variation in the immunogenicity of different strains. The problem was eventually solved by the plaque purification of each serotype and the selection of the most suitable combinations of strains.

By 1982, 15 such strains had been isolated and incorporated into three pentavalent vaccines which gave good immunity when correctly administered.
The cultivation of mouse brain attenuated AHSV in cell cultures soon followed, initially in monkey kidney cells, then also in BHK21 cultures. A major breakthrough was the discovery in the 1970s that the size of AHSV plaques in cell cultures is a marker for virulence (Erasmus 1978). The selection of low-virulence, large plaque variants from cell culture-attenuated cultures became the method of choice for the selection of vaccine strains, replacing the less satisfactory neurotropic strains previously used.
A polyvalent vaccine consisting of two components, one a trivalent (serotypes 1, 3 and 4) and the other a quadrivalent (serotypes 2, 6, 7 and 8) combination, is presently in use. The two components must be administered 3 weeks apart to be effective. Serotypes 5 and 9 are not included because types 8 and 6 afford adequate cross-protection.
Ultrastructure of the orbiviruses
Bluetongue virus was selected for molecular studies in the 1960s because so little was known at the time about the structure and replication of this economically important virus. Following the establishment of the morphological similarity between BTV and AHSV, most of the molecular work was repeated at a later stage for the latter, confirming a close relationship.
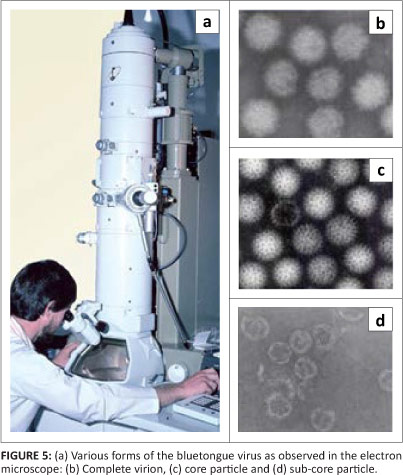
Mass production and purification of the virus, aided by the development of a plaque assay, was the first step before morphological studies and biochemical analysis could start. Surprisingly, electron microscopy revealed two differently sized particles, respectively 54 nm and 70 nm in diameter, which were first thought to represent two viruses.
Further studies revealed the explanation: the presence of a double-layered protein capsid, one of which can be lost during the processing for electron microscopy (Els & Verwoerd 1969; Verwoerd, Huismans & Erasmus 1979). One or both of the layers can also be selectively removed enzymatically, in the latter case giving rise to a highly unstable third 'sub core' particle (Huismans, Van Dijk & Els 1987). The inner capsid has icosahedral symmetry.
The double capsid, unique at the time, led to the description of a new genus, later named Orbivirus after the prominent ringlike capsomeres of the inner layer which are characteristic of both BTV and AHSV as well as other members of the genus discovered later (Verwoerd et al. 1972).
Molecular structure and genetics of the orbiviruses
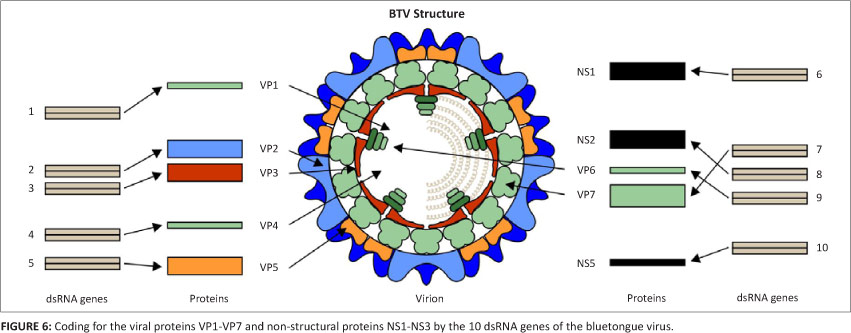
Even more surprising were the results of the chemical and physico-chemical analyses. At the time, all known viruses consisted of a protein capsid containing a genome which was either double-stranded or single-stranded DNA or single-stranded RNA. Chemical analysis clearly indicated an RNA genome but physico-chemical characteristics, including melting curves, sedimentation rate in the ultra-centrifuge, base composition and a heterochromatic staining reaction were closer to those of DNA (Verwoerd 1970). The conclusion that the virus has a double-stranded RNA genome was rather revolutionary and not easily accepted at first. In addition, it was found that the genome consists of 10 segments and the capsid of 7 protein components. It was therefore clear that we were dealing with a uniquely structured virus. Almost simultaneously it had been discovered that reoviruses also possess a segmented double-stranded RNA genome but differ in having a single-layered capsid and in not being transmitted by insects. Consequently, the orbiviruses were classified as a genus in the family Reoviridae.
During the 1970s, our research concentrated on genetic aspects such as the relationship between the genome segments and the viral proteins. It was shown that seven of the segments act as genes, coding for structural proteins in the viral particles, the other three for non-structural proteins found in the infected cell (Verwoerd et al. 1979). At a later stage, the functions of most of these macromolecules were determined. The molecular weights of all the components were determined, as well as the relative copy number of each viral protein (VP) in the viral particle (Huismans & Van Dijk 1990). Of great importance for later work was the assignment of each protein to a genome segment, which was numbered in order of decreasing size, and the identification of VP2 and VP5 as the constituents of the outer capsid, VP1, VP3, VP4, VP6 and VP7 of the core particle and NS1-NS3 as constituents in the infected cell. VP2 is the major component of the outer capsid, responsible for serotype specificity and infectivity, whereas VP7 forms the ring-like capsomere surface layer of the core particle and is the dominant group-specific antigen (Huismans & Erasmus 1981). This was achieved by the in vitro transcription and translation of the mRNA from all 10 genome segments (Van Dijk & Huismans 1988).
The 1980s was also the decade during which many new techniques such as cloning and sequencing of nucleic acids were developed, which collectively became known as biotechnology. These techniques were applied to both BTV and AHSV and all segments of all the serotypes were eventually cloned and sequenced, leading inter alia to the development of much improved new diagnostic tests. Cloned cDNA was first used as probes in hybridisation studies to investigate the genetic relationship between various BTV serotypes. Cross-hybridisation of these DNA clones suggested that segment 6, coding for non-structural protein NS1, is the most suitable group-specific probe, as it is highly conserved in all serotypes, whereas segment 2, coding for an outer protein, is best for serotype-specific probes (Huismans & Cloete 1987). In 1991, a series of sequencing studies on BTV isolates from various countries led to the discovery of 'topotypes', or geographical variants within serotypes that can be used to determine the origin of an outbreak. Improving diagnostic tests started to dominate the scene during this period.
Probes initially also replaced the traditional virus isolation and complement fixation (CF) tests for the identification of orbiviruses or Orbivirus serotypes in field samples but were later in turn replaced by various applications of the polymerase chain reaction (PCR), which excel in sensitivity and speed. Similarly, various enzyme-linked immunosorbent assay (ELISA) techniques have largely replaced the classical CF tests for the detection of Orbivirus antibodies. These techniques also allowed studies of the molecular epidemiology of both diseases and eventually to research aimed at the development of new generation, improved vaccines.
Recombinant subunit vaccines
The multivalent live attenuated vaccines developed and used for many decades in South Africa to combat both BT and AHS were highly successful in a country where both diseases are endemic.
There are, however, a number of problems associated with the use of a multivalent live vaccine, both in an endemic situation and especially in countries where the disease occurs sporadically. The phenomenon of interference between the different serotypes in a multivalent vaccine, which necessitates multiple and repeated vaccinations to achieve full protection, has already been mentioned. There is also always the danger of genetic re-assortment between different serotypes leading to new strains which may escape control, although this has not been observed in this country. Both of these problems can be solved by the use of a monovalent vaccine in the case of a new outbreak, but the danger of the vaccine strain becoming endemic, and possibly reverting to virulence, remains. Another problem is the inability to distinguish by serological means between vaccinated and infected animals. This impacted severely on the racehorse industry in South Africa, where the export of horses to most countries was not allowed for many years and is still strictly controlled to prevent the spread of AHS.
All of these problems could be solved by the development of a recombinant subunit vaccine. Considerable excitement was therefore created when the immunogenicity of a single outer capsid protein (VP2) of BTV was demonstrated in 1987, supporting the feasibility of developing recombinant vaccines (Huismans et al. 1987) Genome libraries were constructed and the first recombinant subunit vaccines developed in collaboration with researchers at Oxford, using the baculovirus of insects as vector (Roy & Erasmus 1992). Various combinations were shown to be effective as monovalent vaccines but, unfortunately, problems encountered in upscaling production have so far precluded their commercial application. The most promising candidate at present is a recombinant AHS vaccine using the canarypox virus as vector, which has been developed by an international consortium, including the Onderstepoort Faculty.
In conclusion, being involved in Orbivirus research for more than two decades was personally a highly satisfying experience and I wish to dedicate this article to all those workers, too many to mention by name, who contributed to the discoveries made over more than a century.
Acknowledgements
Competing interests
The author declares that he has no financial or personal relationship(s) which may have inappropriately influenced him in writing this paper.
References
Alexander, R.A., 1935, 'Studies on the neurotropic virus of horsesickness III: The intracerebral protection test and its application to the study of immunity', Onderstepoort Journal of Veterinary Science and Animal Industry 4, 349-377. [ Links ]
Alexander, R.A., 1938, 'Studies on the neurotropic virus of horsesickness VI: Propagation in the developing chick embryo', Onderstepoort Journal of Veterinary Science and Animal Industry 11, 9-19. [ Links ]
Alexander, R.A., 1947, 'The propagation of bluetongue virus in the developing chick embryo with particular reference to the temperature of incubation', Onderstepoort Journal of Veterinary Science and Animal Industry 22, 7-26. PMid:18915581 [ Links ]
Bayley, T.B., 1856, Notes on the horse sickness at the Cape of Good Hope in 1854-55, Saul Solomons & Co., Cape Town. [ Links ]
Coetzer, J.A.W. & Tustin, R.C. (eds.), 2004, Infectious diseases of livestock, 2nd edn., Oxford University Press, Cape Town. [ Links ]
Du Toit, R.M., 1944, 'The transmission of bluetongue and horsesickness by Culicoides', Onderstepoort Journal of Veterinary Science and Animal Industry 19, 7-16. [ Links ]
Edington, A., 1892, 'Horse-sickness microbe', Grahamstown Journal August 11, 3. [ Links ]
Edington, A., 1900, 'South African horse sickness: Its pathology and methods of protective inoculation', Journal of Comparative Pathology and Therapeutics 18, 142-155. [ Links ]
Edington, A., 1904, Report of the Director of the Colonial Bacteriological Institute 1894-1904. [ Links ]
Els, H.J. & Verwoerd, D.W., 1969, 'Morphology of bluetongue virus', Virology 38, 213219. http://dx.doi.org/10.1016/0042-6822(69)90362-6 [ Links ]
Erasmus, B.J., 1978, 'A new approach to polyvalent immunization against African horsesickness', in J.T. Bryans & H. Gerber (eds.), Equine infectious diseases, Proceedings of the Fourth International Conference on Equine Infectious Diseases, Veterinary Publications Inc., Princeton, New Jersey, September 24-27, 1976, pp. 401-403. [ Links ]
Erasmus, B.J. & Potgieter, A.C., 2009, 'The history of bluetongue', in P. Mertens, M. Bayliss & P. Mellor (eds.), Bluetongue, pp. 7-21, Elsevier, Academic Press, London. http://dx.doi.org/10.1016/B978-012369368-6.50006-X [ Links ]
Haig, D.A., McKercher, D.G. & Alexander, R.A., 1956, 'The cytopathogenic action of bluetongue virus on tissue cultures and its application to the detection of antibodies in the serum of sheep', Onderstepoort Journal of Veterinary Research 27, 171-177. [ Links ]
Howell, P.G., 1960, 'A preliminary antigenic classification of strains of bluetongue virus', Onderstepoort Journal of Veterinary Research 28, 357-363. [ Links ]
Howell, P.G., 1962, 'The isolation and identification of further antigenic types of African horsesickness virus', Onderstepoort Journal of Veterinary Research 29, 139-149. [ Links ]
Huismans, H. & Erasmus, B.J., 1981, 'Identification of the serotype specific and group-specific antigens of bluetongue virus', Onderstepoort Journal of Veterinary Research 48, 51-58. PMid:6273773 [ Links ]
Huismans, H. & Cloete, M., 1987, 'A comparison of different cloned bluetongue virus segments as probes for the detection of virus-specified RNA', Virology 158, 373380. http://dx.doi.org/10.1016/0042-6822(87)90209-1 [ Links ]
Huismans, H., Van Dijk, A.A. & Els, H.J., 1987, 'Uncoating of parental bluetongue virus to core and subcore particles in infected L-cells', Virology 157, 180-188. http://dx.doi.org/10.1016/0042-6822(87)90327-8 [ Links ]
Huismans, H., Van der Walt, N.T., Cloete, M. & Erasmus, B.J., 1987, 'Isolation of a capsid protein of bluetongue virus that induces a protective immune response in sheep', Virology 157, 172-179. http://dx.doi.org/10.1016/0042-6822(87)90326-6 [ Links ]
Huismans, H. & Van Dijk, A.A., 1990, 'Bluetongue virus structural components', Current topics in Microbiology and Immunology 162, 2141. http://dx.doi.org/10.1007/978-3-642-75247-6_2 [ Links ]
Hutcheon, D., 1880, 'Fever or epizootic catarrh', p. 12, Report of the Colonial Veterinary Surgeon for 1880. [ Links ]
Mason, J.H., Coles, J.D.W.A. & Alexander, R.A., 1940, 'Cultivation of bluetongue virus in fertile eggs produced on a vitamin deficient diet', Nature 145, 1022-1023. http://dx.doi.org/10.1038/1451022a0 [ Links ]
M'Fadyean, J., 1900, 'African horse-sickness', Journal of Comparative Pathology 13,1-20. [ Links ]
McIntosh, B.M., 1958, 'Immunological types of horsesickness virus and their significance in immunization', Onderstepoort Journal of Veterinary Research 27, 465-539. [ Links ]
Neitz, W.O., 1948, 'Immunological studies on bluetongue in sheep', Onderstepoort Journal of Veterinary Science and Animal Industry 23, 93-136. PMid:18863435 [ Links ]
Pitchford, N. & Theiler, A., 1903, 'Investigations into the nature and causes of horsesickness', Agricultural Journal of the Cape of Good Hope 23, 153-156. [ Links ]
Roy, P. & Erasmus, B.J., 1992, 'Second generation candidate vaccine for bluetongue disease', in T.L. Barber & M.M. Jochim (eds.), Bluetongue and related orbiviruses, pp. 856-867, Alan R. Liss, Inc., New York. [ Links ]
Spreull, J., 1905, 'Malarial catarrhal fever (bluetongue) of sheep in South Africa', Journal of Comparative Pathology 18, 321-337. [ Links ]
Theiler, A., 1905, 'Horsesickness - introduction', Report of the Government Veterinary Bacteriologist, 131-162. [ Links ]
Theiler, A., 1908, 'Further notes on immunity in horsesickness', Report of the Government Veterinary Bacteriologist, 89-161. [ Links ]
Theiler, A., 1921, 'African horsesickness (pestis equorum)', Scientific Bulletin 19, 1-29. [ Links ]
Van Dijk, A.A. & Huismans, H., 1988, 'The in vitro activation and further characterization of bluetongue virus mRNA', Journal of General Virology 69, 573-581. http://dx.doi.org/10.1099/0022-1317-69-3-573, PMid:2832524 [ Links ]
Verwoerd, D.W., 1969, 'Purification and characterization of bluetongue virus', Virology 38, 203-212. http://dx.doi.org/10.1016/0042-6822(69)90361-4 [ Links ]
Verwoerd, D.W., 1970, 'Diplornaviruses: A newly recognized group of double-stranded RNA viruses', Progress in Medical Virology 12, 192-210. [ Links ]
Verwoerd, D.W., Els, H.J., De Villiers, E.M. & Huismans, H., 1972, 'Structure of the bluetongue virus capsid', Journal of Virology 10, 783-794. PMid:4117349 [ Links ]
Verwoerd, D.W., Huismans, H. & Erasmus, B.J., 1979, 'Orbiviruses', in H. Fraenkel-Conrat & R.R. Wagner (eds.), Comprehensive Virology, pp. 285-345, Plenum Press, New York. [ Links ]
Wilson, A., Darpel, K. & Mellor, P.S., 2008, 'Where does bluetongue virus sleep in the winter?', PLoS Biology 6(8), e210. http://dx.doi.org/10.1371/journal.pbio.0060210 [ Links ]
 Correspondence to:
Correspondence to:
Daniel Verwoerd
PO Box X04
Onderstepoort 0110, South Africa
daan.verwoerd@up.ac.za
Received: 25 June 2012
Accepted: 28 June 2012
Published: 13 Nov. 2012
Paper given at the 30th World Veterinary Congress, October 2011, Cape Town, South Africa.

