










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.83 n.1 Pretoria Jan. 2012
CLINICAL COMMUNICATION
Doramectin toxicity in a group of lions (Panthera leo)
Remo G. LobettiI; Peter CaldwellII
IBryanston Veterinary Hospital, Bryanston, South Africa
IIOld Chapel Veterinary Clinic, Totiusdal, South Africa
ABSTRACT
Ten lions (Panthera leo) that were treated with a single injection of doramectin at a dose ranging between 0.2 mg/kg and 0.5 mg/kg showed clinical signs consistent with avermectin toxicity, namely ataxia, hallucinations, and mydriasis. Two subsequently died whereas the other eight lions recovered after 4-5 days with symptomatic therapy. Post-mortem examinations of the two that died showed cyanosis, severe pulmonary oedema, pleural effusion, and pericardial effusion, with histopathology not revealing any abnormalities. In both these lions, doramectin brain and liver tissue concentrations were elevated. Although doramectin is regularly used in wild felids, to date there have been no reports of avermectin toxicity in the literature. This article highlights the potential for doramectin toxicity in this species.
Introduction
Doramectin is a novel avermectin that is produced by mutational biosynthesis and differs from ivermectin by having a cyclohexyl group in the C25 position of the avermectin ring (Goudie et al. 1993; Reinemeyer & Courtney 2001). Doramectin has a broad range of activity against endoparasites and ectoparasites, and although only approved for use in cattle, sheep, goats and swine (Plumb 1999; Reinemeyer & Courtney 2001), off-label use has been widely used and reported in other species (Yas-Natan et al. 2003). In lions, it is used extra-label, both as an anthelmintic against intestinal round worms and systemic filaroides, as well as an ectoparasitic against ticks, blow fly strike, mites, and ear mites.
Like other macrocyclic lactones, doramectin has two modes of action. The primary mode of action is binding of the doramectin molecule to postsynaptic glutamate-gated chloride ion channels in the synapses between inhibitory interneurons and excitatory motor neurons in nematodes, and in myoneural junctions in arthropods (Plumb 1999; Reinemeyer & Courtney 2001). Avermectins also enhance the release of gamma-aminobutyric-acid (GABA) in presynaptic neurons, which, in turn, open postsynaptic GABA-gated chloride channels (Hopper, Aldrich & Haskins 2002; Plumb 1999; Reinemeyer & Courtney 2001). In either case, the influx of chloride ions reduces cell membrane resistance, which prevents the potential hyperpolarisation of neural stimuli to muscles, resulting in flaccid paralysis and death (Plumb 1999; Reinemeyer & Courtney 2001).
The avermectins are highly toxic to invertebrates and can be toxic to mammals, with clinical signs of toxicity being mydriasis, vomiting, seizure activity, and/or tremors, coma, and death from respiratory failure (Plumb 1999; Reinemeyer & Courtney 2001). Ivermectin toxicity in collies and other dog breeds has been well documented and has been associated with diffuse cerebral and cerebellar dysfunction. Signs of toxicosis are related to the penetration of the drug into the central nervous system (CNS) (Yas-Natan et al. 2003).
Doramectin has a plasma half-life of approximately twice that of ivermectin in cattle (Reinemeyer & Courtney 2001). The difference between the two drugs can be explained by the greater non-polarity of doramectin and its slower metabolism due to the cyclohexyl group (Goudie et al. 1993, Perez et al. 2002; Toutain et al. 1997). Thus, the clinical course of doramectin toxicity would be expected to be longer when compared with ivermectin toxicity.
Although regularly used in wild felids as an endo-and ectoparasitic drug , to date there have been no reports of avermectin toxicity in the literature. This article describes doramectin toxicity in a group of lions.
Case history
Ten captive lions (four tawny and six white) were treated with a single injection of doramectin at a dose ranging between 0.2 mg/kg and 0.5 mg/kg, given either subcutaneously or intramuscularly with a pole syringe, depending on the angle of the pole syringe and accessibility.
The animals' body weights were estimated, which may have been inaccurate, and they were thus possibly given a much higher dose of doramectin. The lions' weights were estimated on average age weights, as sub-adult and young adults generally range in weight between 150 kg and 200 kg. Two days later, all ten animals were fed a horse carcass. The horse had been treated with doramectin, butylscopolamine, and flunixin meglumine in the neck muscles two days before being shot. Three to 5 days later all ten lions started to show clinical signs of ataxia, hallucinations, and mydriasis. The lions appeared to be seeing objects that did not exist, as they would follow an imaginary object with their eyes and paw at it and would also sit and stare into the distance without being aware of their surroundings. It was also noted that the white lions showed more severe clinical signs, namely severe disorientation, loss of balance, ataxia of all four limbs, and falling over, whereas the tawny lions were less disorientated and ataxic. Two subsequently died, five days after ingestion of the carcass, both of which were white lions. One was an adult (18 months) and the other a sub-adult (5 months). The other eight lions recovered after 4-5 days with symptomatic treatment, which included intravenous and subcutaneous fluids, short-acting corticosteroids, broadspectrum antibiotics, omeprazole, sucralfate, butaphosphate as an appetite stimulant, and vitamin B1 and A injections. Husbandry management was maintained by keeping them in a warm, protected environment.
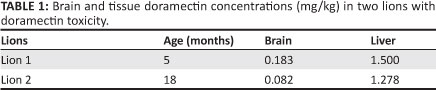
Post-mortem examinations of the two lions that died revealed cyanosis, severe pulmonary oedema, pleural effusion, and mild pericardial effusion, with histopathology not revealing any abnormalities. Brain and liver tissue from both these lions showed elevated doramectin concentrations (Table 1).
Discussion
Ivermectin at a therapeutic dose of 0.2 mg/kg can result in toxicosis in predisposed dogs (Hopper et al. 2002; Tranquilli, Paul & Seward 1989). In contrast, dosages of 2 mg/kg can be administered to beagles and non ivermectin-sensitive Collies without evidence of drug toxicity (Pulliam et al. 1985). The dose of doramectin used in this group of lions ranged between 0.2 mg/kg and 0.5 mg/kg, which is what has been used in dogs (Borst & Oude-Elferink 2002). However, as the lion's body weights were estimated, this could have been inaccurate, resulting in a higher than recommended dose of doramectin being administered. In addition, the lions were fed a horse carcass and although speculative, the neck muscles may have been contaminated with doramectin as the horse had been treated with doramectin prior to death. Other lions from the same camp did not show any clinical signs of doramectin toxicity. As these lions had not been treated with doramectin, it could mean that either they were not susceptible to doramectin or that they did not ingest any contaminated horsemeat. It was also observed that some lions fed on the neck and others on the rump.
Reported signs of acute ivermectin toxicity in predisposed and overdosed dogs include drooling, mydriasis, bradycardia, hyperthermia, ataxia, apparent blindness, decreased menace response, muscle tremors and disorientation, which may progress to weakness, recumbency, unresponsiveness, stupor and coma (Hopkins, Marcella & Strecker 1990; Hopper et al. 2002; Paul & Tranquilli 1989; Reinemeyer & Courtney 2001).
Doramectin toxicity has been reported in a Border collie (Yas-Natan et al. 2003). The diagnosis was based on the neurological signs, which were similar to those previously reported for ivermectin toxicity, namely ataxia, disorientation, head pressing, hypersalivation, and bilateral lack of menace response, with normal cranial nerves, pupillary size, light response, spinal reflexes, and postural reactions. Similar clinical signs, namely ataxia, hallucinations, and mydriasis were present in these lions. Unfortunately, neurological examinations were not feasible. As death from respiratory failure has been reported with avermectin toxicity (Paul & Tranquilli 1989; Plumb 1999; Reinemeyer & Courtney 2001), the two lions that died could have died from respiratory failure, as both showed cyanosis on post-mortem examination, without any obvious other aetiology.
Normal and toxic concentrations of doramectin in the brain and liver of Felidae have to date not been reported; however, concentrations have been reported in cattle. In a herd of Murray Grey cattle, 8 out of 312 cattle treated with a therapeutic dose of ivermectin showed clinical signs of hypersensitivity, with a mean brain concentration of 0.056 mg/kg in affected animals and 0.004 mg/kg in unaffected cattle (Seaman et al. 1987). Thus the levels of 0.183 mg/kg and 0.083 mg/kg reported in these two lions would be very toxic. In addition, the clinical signs and relatively insignificant post-mortem findings would also support this conclusion.
Mammals, unlike nematodes and arthropods, have GABAmediated inter-neuronal inhibitors only in the CNS (Hopkinsv et al. 1990; Hopper et al. 2002; Plumb 1989). It is believed that the mammalian blood brain barrier (BBB) is impermeable to avermectins and therefore toxicity in mammals ensues at a much higher avermectin concentration compared with nematodes and arthropods (Hopper et al. 2002). An important component of the BBB is the P-glycoprotein. P-glycoprotein is an ATP-driven efflux transporter (ABC transporter), encoded by the multidrug-resistance gene MDR1/ABCB1 (Dean, Rzhetsky & Allikmets 2001). It is known that MDR1 P-glycoprotein transports a wide range of structurally unrelated lipophilic and amphipathic drugs, toxins, and xenobiotics, including many commonly used veterinary drugs (Geyer et al. 2007). The MDR1 efflux machinery protects the organism from exposure to drugs and environmental xenobiotics by decreasing their absorption in the intestinal tract and promoting their excretion into bile and urine. Additionally, MDR1 P-glycoprotein in the blood-brain barrier highly restricts the entry of drugs and xenobiotics into the central nervous system (Borst & Oude Elferink 2002; Fromm 2000). A mutation in the MDRI gene has been described in ivermectin-sensitive collies (Mealey, Bentjen & Waiting 2002). The product of the MDRI gene, P-glycoprotein, is an important component of the blood-brain barrier, and is a drug transporter expressed in the luminal membrane of brain capillary endothelial cells (Mealey et al. 2002). Its function in the brain is to transport certain substrates, including ivermectin, from the brain tissue back into the capillary lumen, thus lowering the concentrations of these drugs in the CNS. The frequency of the homozygous mutant genotype in a population of collies in Washington and Idaho was 35%, which is similar to the estimated prevalence of ivermectin sensitivity in collies (Mealey et al. 2002). Determination of the MDR1 genotype of the affected lions was not performed; however, it is reasonable to assume that the affected lions were homozygous for the mutant allele, and that this was more prevalent in white lions, as they showed more severe clinical signs than the others.
Conclusion and recommendations
In conclusion, the very high brain and liver concentrations of doramectin in two lions, typical clinical signs, and relatively insignificant post-mortem findings all support the diagnosis of doramectin toxicity in the lions. Thus, doramectin should be used with care in this species and future studies looking at the presence or absence of the MDR1 gene in lions are also warranted.
Acknowledgements
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this paper.
Authors' contributions
R.G.L. (Bryanston Veterinary Hospital), consultation on the case, literature search, writing of the article. P.C. (Old Chapel Veterinary Clinic) data collection, case consultation, writing of the article.
References
Borst, P. & Oude-Elferink, R., 2002, 'Mammalian ABC transporters in health and disease', Annual Review of Biochemistry 71, 537-592. http://dx.doi.org/10.1146/annurev.biochem.71.102301.093055, PMid:12045106 [ Links ]
Dean, M., Rzhetsky, A. & Allikmets, R., 2001, 'The human ATP-binding cassette (ABC) transporter superfamily', Genome Research 11, 1156-1166. http://dx.doi.org/10.1101/gr.GR-1649R, PMid:11435397 [ Links ]
Fromm, M.F., 2000, 'P-glycoprotein: A defense mechanism limiting oral bioavailability and CNS accumulation of drugs', International Journal of Clinical Pharmacology and Therapeutics 38, 69-74. PMid:10706193 [ Links ]
Geyer, J., Klintzsch, S., Meerkamp, K., Wohlke, A., Distl, O., Moritz, A. et al., 2007, 'Detection of the nt230(del4) MDR1 mutation in White Swiss Shepherd dogs: Case reports of doramectin toxicosis, breed predisposition, and microsatellite analysis', Journal of Veterinary Pharmacology and Therapy 30, 482-485. http://dx.doi.org/10.1111/j.1365-2885.2007.00885.x, PMid:17803743 [ Links ]
Goudie, A.C., Evans, N.A., Gration, K.A.F., Bishop, B.F., Gibson, S.P., Holdom, K.S. et al., 1993, 'Doramectin - A potent novel endectocide', Veterinary Parasitology 49, 5-15. http://dx.doi.org/10.1016/0304-4017(93)90218-C [ Links ]
Hopkins, K.D., Marcella, K.L. & Strecker, A.E., 1990, 'Ivermectin toxicosis in a dog', Journal of the American Veterinary Medical Association 197, 93-94. PMid:2370228 [ Links ]
Hopper, K., Aldrich, J. & Haskins, S.C., 2002, 'Ivermectin toxicity in 17 collies', Journal of Veterinary Internal Medicine 16, 89-94. http://dx.doi.org/10.1111/j.1939-1676.2002.tb01611.x, PMid:11822811 [ Links ]
Mealey, K.L., Bentjen, S.A. & Waiting, D.K., 2002, 'Frequency of the mutant MDR1 allele associated with ivermectin sensitivity in a sample population of Collies from the northwestern United States', American Journal of Veterinary Research 63, 479-481. http://dx.doi.org/10.2460/ajvr.2002.63.479, PMid:11939306 [ Links ]
Paul, A. & Tranquilli, W., 1989, 'Ivermectin', in R.W. Kirk & J.D. Bonagura (eds.), Current veterinary therapy X: Small animal practice, pp. 140-142, WB Saunders, Philadelphia. [ Links ]
Perez, R., Cabezas, I., Godoy, C., Rubilar, L., Munoz, L., Arboix, M. et al., 2002, 'Pharmacokinetics of doramectin and ivermectin after oral administration in horses', Veterinary Journal 163, 161-167. http://dx.doi.org/10.1053/tvjl.2001.0624 [ Links ]
Plumb, D.C., 1999, Veterinary drug handbook, 3rd edn., Iowa State University Press, Ames. [ Links ]
Pulliam, J.D., Seward, R.L., Henry, R.T. & Steinberg, S.A., 1985, 'Investigating ivermectin toxicity in Collies', Veterinary Medicine 80, 33-40. [ Links ]
Reinemeyer, C.R. & Courtney, C.H., 2001, 'Antinematodal drugs', in H.R. Adams (ed.), Veterinary pharmacology and therapeutics, 8th edn., pp. 963-969, Iowa State University Press, Ames. [ Links ]
Seaman, J.T., Eagleson, J.S., Carrigan, M.J. & Webb, R.F., 1987, 'Avermectin B toxicity in a herd of Murray Grey cattle', Australian Veterinary Journal 64, 284-285. http://dx.doi.org/10.1111/j.1751-0813.1987.tb15963.x, PMid:3426470 [ Links ]
Toutain, P.L., Upson, D.W., Terhune, T.N. & Mckenzie, M.E., 1997, 'Comparative pharmacokinetics of doramectin and ivermectin in cattle', Veterinary Parasitology 72, 3-8. http://dx.doi.org/10.1016/S0304-4017(97)00070-8 [ Links ]
Tranquilli, W.J., Paul, A.J. & Seward, R.L., 1989, 'Ivermectin plasma concentrations in collies sensitive to ivermectin-induced toxicosis', American Journal of Veterinary Research 50, 769-770. PMid:2729723 [ Links ]
Yas-Natan, E., Shamir, M., Kleinbart, S. & Aroch, I., 2003, 'Doramectin toxicity in a collie', Veterinary Record 153, 718-720. PMid:14690080 [ Links ]
 Correspondence to:
Correspondence to:
Remo Lobetti
PO Box 67092
Bryanston 2021
South Africa
rlobetti@mweb.co.za
Received: 24 Feb. 2012
Accepted: 12 June 2012
Published: 15 Oct. 2012

