










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkJournal of the South African Veterinary Association
versión On-line ISSN 2224-9435
versión impresa ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.83 no.1 Pretoria ene. 2012
ORIGINAL RESEARCH
Prevalence of pansteatitis in African sharptooth catfish, Clarias gariepinus (Burchell), in the Kruger National Park, South Africa
K. David A. HuchzermeyerI, II
ISterkspruit Veterinary Clinic, Lydenburg, South Africa
IIDepartment of Paraclinical Sciences, University of Pretoria, South Africa
ABSTRACT
Pansteatitis was confirmed in sharptooth catfish, Clarias gariepinus (Burchell), from three main locations within the Kruger National Park (KNP); the Olifants River Gorge, Engelhard Dam on the Letaba River and from the Sabie River in the Sabiepoort. An increasing prevalence of pansteatitis was observed in catfish during repeated samplings from the Olifants Gorge from 2009 to 2011 and co-existence of old and recent lesions indicated on-going incitement of pansteatitis. Only a low prevalence of pansteatitis was observed in catfish sampled from the Olifants River upstream of the Gorge in the KNP and no pansteatitis was observed in catfish sampled from a rain-filled dam not connected to the Olifants River. Common to both the Olifants Gorge and the Sabiepoort is the damming of the rivers in Mozambique to form lakes Massingir and Corumana respectively. Anthropogenic activities resulting in potential pollution of the rivers differ greatly between these two catchments, providing argument against a primary pollution-related aetiology of the pansteatitis found at these two sites. Compared with other sites, analysis of stomach contents of catfish from the Olifants Gorge and the Sabiepoort strongly suggested that consumption of a predominantly fish diet was associated with the development of pansteatitis in these fish. In a farmed population of catfish used as positive control, development of pansteatitis could be ascribed to consumption of rancid fish waste from a trout slaughterhouse. In the Olifants Gorge, alien invasive silver carp, Hypophthalmychthys molitrix (Valenciennes), seasonally migrate upstream out of Lake Massingir to spawn. This schooling species is an obligate phytoplankton feeder with consequent high levels of adipose tissue n-3 polyunsaturated fatty acids. In the Olifants Gorge, at least, this may explain seasonal exposure to levels of polyunsaturated fats in the diets of catfish and crocodiles to which these animals are not adapted. The possible roles of diet, membrane lipid composition and metabolic rate of fish, sediment pollution and seasonal drop in environmental temperature in the pathogenesis of pansteatitis in the catfish are discussed. Further studies are needed to verify some of these speculations.
Introduction
Much attention has been focused on the state of the Olifants River in Mpumalanga Province, South Africa. From its origins on the Highveld plateau the river flows eastwards down the escarpment traversing the Kruger National Park (KNP) and Mozambique before discharging into the Indian Ocean. The catchment has been heavily impacted by human activity including mining, coal-fired electricity generation, industrial and urban wastewater discharges, agricultural practices and water impoundments. As a result, the Olifants River is regarded as one of the most threatened aquatic ecosystems in Mpumalanga (Ashton 2010; De Villiers & Mkwelo 2009; Heath, Coleman & Engelbrecht 2010). Since 2003, increasing Nile crocodile, Crocodylus niloticus Laurenti, mortalities in Lake Loskop, situated in the upper Olifants catchment, have coincided with periodic mass fish mortalities (Botha, Van Hoven & Guillette 2011). In the KNP, an estimated 180 large crocodiles died in the Olifants River Gorge during the winter of 2008 following the raising of the sluice gates of the Lake Massingir dam wall in Mozambique during 2007 (Ferreira & Pienaar 2011; Huchzermeyer et al. 2011). Fewer deaths were recorded in the subsequent two winters. South African National Parks (SANParks) veterinarians established the cause of death as pansteatitis.
Steatitis, the inflammation associated with fat cell necrosis, has been described from many species of warm and cold-blooded animals, including fish. This nutritional disorder is found mainly in farmed and captive animals and rarely in free-living wild animals. Feeding of large amounts of unsaturated fat, particularly if rancid, or diets deficient in vitamin E are known to cause pansteatitis (Goodwin 2006; Herman & Kircheis 1985; Roberts & Agius 2008; Roberts, Richards & Bullock 1979). The condition has been reported in farmed crocodiles fed fish that was no longer fresh (Huchzermeyer 2003; Ladds et al. 1995). In Lake Loskop, large-scale fish mortality was observed as a result of acid mine drainage and may explain the development of pansteatitis in the resident crocodiles after the fish die-off (J. Myburgh and co-workers, University of Pretoria, pers. comm., 2009). It has been proposed that bio-accumulation, via algae, of aluminium and iron within body fat was the cause of yellow discolouration of fat of Mozambique tilapia, Oreochromis mossambicus (Peters), in Lake Loskop and that this bio-accumulation may have provided a trigger for development of pansteatitis in higher trophic level predators (Oberholster et al. 2011). In the Olifants Gorge, significant overt fish mortality has seldom been observed, with the exception of a single localised event affecting almost exclusively large African sharptooth catfish, Clarias gariepinus (Burchell), during the winter of 2009 (D. Pienaar & D. Govender, SANParks, Skukuza, pers. comm., 2009). During the 2011 aerial crocodile survey by SANParks, however, three large dead catfish were observed in the Olifants Gorge and in Lake Massingir (D. Pienaar, SANParks, Skukuza, pers. comm., 2011) suggesting that low level mortality was occurring.
There are numerous references in the literature linking lipid peroxidation to pollutants in the aquatic environment (Bainy et al. 1996; Baker, Martin & Davies 1997; Kelly et al. 1998; Kibanova, Nieto-Camacho & Cervini-Silva 2009), but little has been published linking these effects to pansteatitis, which is regarded as a nutritional disease. Because of a possible link between pansteatitis in catfish and crocodiles, the study of pansteatitis in catfish was initiated as part of the multidisciplinary investigation into the crocodile mortality in the KNP under the auspices of the Consortium for the Restoration of the Olifants Catchment (CROC).
Materials and methods
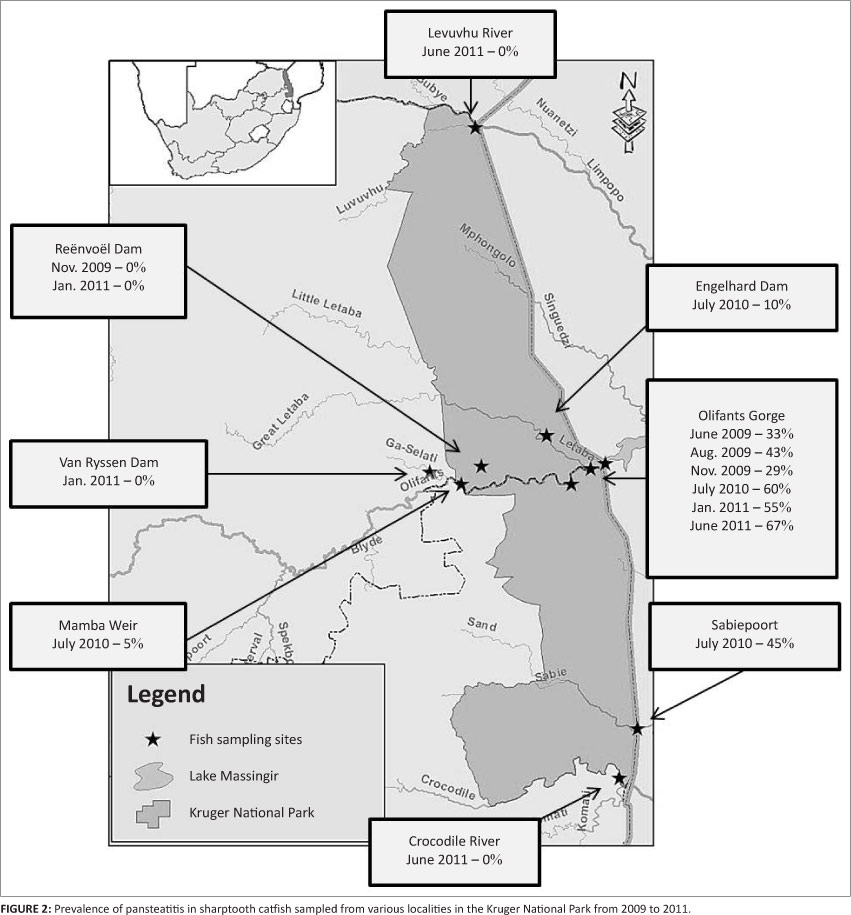
Sharptooth catfish were collected during the winter and summer months from various localities within and outside the KNP, including two negative reference populations, namely Reënvoël Dam (23°58'37.2"S 31°19'38.4"E) that has its entire catchment within the KNP, and Van Ryssen Dam (24°00'13.6"S 31°05'36.9"E) at the FOSKOR phosphate mine in Phalaborwa just west of the KNP. A farmed population of sharptooth catfish at Lunsklip Fisheries near Lydenburg, Mpumalanga, served as a positive reference population. Subject to catch success, up to 20 fish were collected on each sampling occasion (Table 1). From 2009, fish were sampled repeatedly from the Olifants Gorge. This section of river stretches eastwards from the confluence of the Olifants and Letaba rivers (23°59'21.8"S 31°49'35.6"E), through a 9 km long gorge in the Lebombo Mountains, to where it enters Lake Massingir on the Mozambique border (23°57'48"S 31°52'97"E). Further samplings took place from the Olifants River at Mamba Weir (24°03'32"S 31°14'14"E), where the Olifants River enters the western boundary of the KNP, from Engelhard Dam (23°50'19"S 31°28'28"E) on the Letaba River, and further south in the KNP from the Sabiepoort (25°10'25.41"S 32°02'23.42"E), where the Sabie River enters Lake Corumana on the Mozambique border. In the north of the KNP, fish were sampled from the Levuvhu River (22°25'51.0"S 31°18'04.4"E) and on the southern boundary of the Park from the Crocodile River (25°23'57.1"S 31°57'29.9"E). Fish were caught by baited hook and line and by netting. During the June 2011 sampling in the Olifants Gorge, 21 tiger fish, Hydrocynus vittatus Castelnau, were also sampled from the confluence of the Olifants and Letaba rivers.
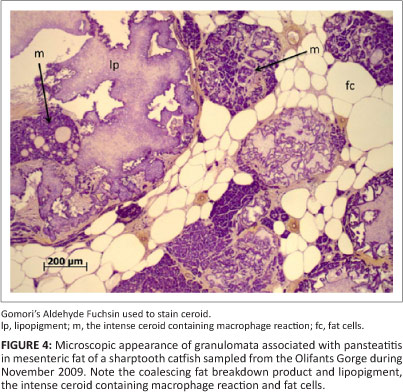
All fish were anaesthetised using benzocaine hydrochloride (Kyron Laboratories, Johannesburg, South Africa) and subjected to body mass and length measurements, body condition scoring and sexing. Body condition was scored on a scale ranging from 1 (emaciated) to 5 (obese). Blood was collected from the vessels ventral to the vertebral column in the tail region. Each fish was euthanized with an overdose of benzocaine hydrochloride and a detailed autopsy was performed. Positive pansteatitis cases were identified by presence of grossly observable lesions in the fat. Tissue specimens from organs and abdominal and subcutaneous adipose tissues were collected in 10% buffered formalin from all sampled fish and subjected to standard histological techniques. All tissue sections were stained with haematoxylin and eosin and examined by light microscopy. Selected sections were stained with Gomori's Aldehyde Fuchsin to demonstrate presence of ceroid within the adipose tissues and in macrophages associated with necrotic lesions in the fat. The stomach contents of each fish were examined and recorded. The sagittal otoliths were removed from all sampled catfish, embedded in resin and sectioned transversely at 0.4 mm thickness by microtome according to the method of Weyl and Booth (2008). Mounted sections were examined under the light microscope and growth zones were counted for age determination.
Ethical considerations
The project was approved by the Animal Use and Care Committee of the University of Pretoria under Protocol VO13/10.
Results
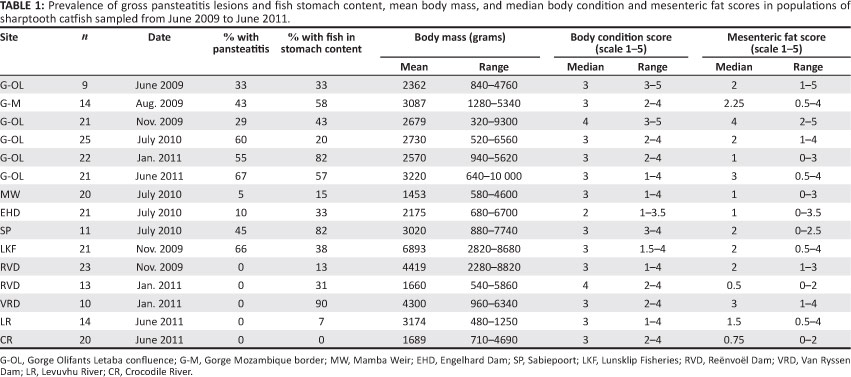
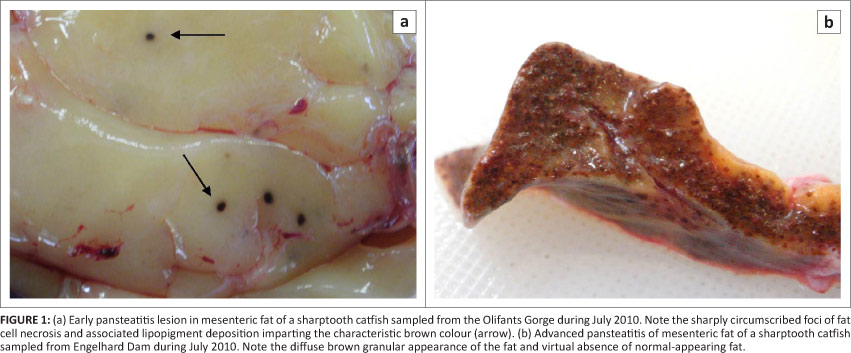
During the period from June 2009 to June 2011, 265 specimens of C. gariepinus were examined during 15 sampling episodes from the various localities within and outside of KNP, including the positive and negative control populations (Table 1). Pansteatitis was observed in mesenteric adipose tissues of fish from the Olifants Gorge, Sabiepoort, Engelhard Dam, Mamba Weir and Lunsklip Fisheries and differed little except in degree of severity. Gross lesions consisted of small focally disseminated to coalescing granulomata up to 5 mm in diameter characterised by a brown colour, sometimes with an orange-coloured centre (Figure 1). Affected fat in severe cases had a rubbery consistency. In the adipose tissues, gross lesions of pansteatitis were confirmed histologically by the presence of foci of adipocyte necrosis with extracellular lipopigment surrounded by an intense infiltration of ceroid-containing macrophages. The detailed pathology and histopathology of the organs and the specific lesions associated with fat necrosis in fish from the Olifants Gorge have been published elsewhere (Huchzermeyer et al. 2011). Pansteatitis was not found in the 21 tiger fish collected from the Olifants Gorge during June 2011.
Mesenteric fat reserves varied from sparse to prominent in catfish represented in samplings from most sites but mesenteric fat stores of most catfish from the Olifants Gorge were larger than those of catfish sampled from other localities in KNP. Catfish from Reënvoël Dam, Engelhard Dam, the Crocodile River and Mamba Weir were leaner. Most fish from Lunsklip Fisheries had prominent mesenteric fat reserves.
Catfish sampled from the Olifants Gorge and from Reënvoël Dam ranged in age from 1 to 19 years. In the Olifants Gorge, pansteatitis was detected in catfish ranging from 3 to 19 years with both sexes equally affected. The main focus of the study was on the Olifants River Gorge, where an increase in prevalence of pansteatitis in sampled catfish was detected since structured sampling began in 2009 (Figure 2). Presence of gross lesions was used to determine prevalence of pansteatitis (Figures 2 and 3). Microscopic examination of histological sections of fat from sampled fish confirmed the macroscopic diagnosis of pansteatitis (Figure 4). Co-existence of coalescing granulomata, scarring of the adipose tissues, and more recent lesions characterised by small foci of brown discolouration of the fat suggested on-going incitement of fat necrosis and attempts at healing in catfish from the Olifants Gorge.
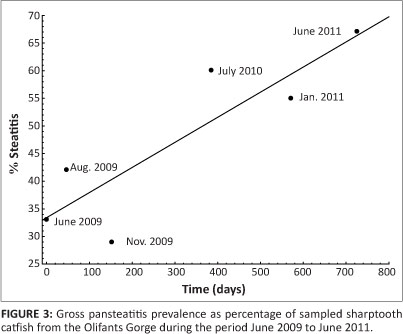
Catfish from Lunsklip Fisheries were fed almost exclusively on untreated waste, rich in polyunsaturated fat, from the trout slaughterhouse on this farm. This waste, consisting largely of fat rich innards, was dumped into the catfish pond where it decomposed until consumed by the fish. These fish showed a high prevalence of pansteatitis as expected under such nutritional conditions, and provided the study with a positive reference population (Table 1). In the Olifants Gorge, analysis of stomach contents showed that catfish fed predominantly on fish as well as insects and small reptiles that had been washed into the river during flood conditions. On the Mozambique border where the Olifants River flows into Lake Massingir and where the sand-bottomed pools and rapids have been inundated with clay deposits, sampled catfish appeared to be feeding off the surface of the clay, and ingesta consisted of algal detritus and clay. In the Sabie River, the Sabiepoort has also been partially inundated with clay sediments as a result of Lake Corumana in Mozambique damming this gorge. A single sampling revealed a similar prevalence of pansteatitis in catfish to that found in the Olifants Gorge (Figure 2). Several of these fish appeared to have been feeding off a dead crocodile with pansteatitis. In the Olifants Gorge both crocodiles and catfish have been observed feeding off the carcasses of dead crocodiles (D. Pienaar, SANParks, Skukuza, pers. comm., 2009). Fish from Mamba Weir were lean and had fed heavily on the fruit of sycamore fig trees, Ficus sycomorus, which overhang the river embankment. Prevalence of pansteatitis was significantly lower than that in the Olifants Gorge (Figure 2). Fish sampled from Van Ryssen Dam showed no lesions of pansteatitis (Table 1). They appeared to have fed exclusively on Mozambique tilapia. The stomachs of the majority of fish sampled from the Crocodile River during June 2011 were distended with ingested filamentous algae. Microscopic examination of fluid expressed from stomach contents revealed large numbers of diatoms together with filamentous algae. Fish sampled from the Levuvhu River during June 2011 had been feeding off algae and sycamore figs. No pansteatitis was detected in fish from the Crocodile and Levuvhu rivers (Table 1). Fish remnants, vegetation, invertebrate and algal detritus were equally represented in stomach contents of fish from Reënvoël Dam. Gross steatitis was not detected (Table 1). Histological examination, however, revealed small numbers of lipopigment-containing macrophages within the mesenteric adipose tissue in one fish (3.6% of sampled fish [n = 41]) from Reënvoël Dam. The associated small focus of lipopigment-containing necrotic cell remnants was in the proximity of a large parasitic cyst. Stomach contents of fish from Engelhard Dam included fish remnants, plant and algal detritus. Pansteatitis prevalence was 10% (n = 21) (Figure 2).
Discussion
Poikilothermic animals such as fish require inclusion of highly polyunsaturated fatty acids within biological membranes to maintain membrane fluidity necessary for normal metabolism at the relatively colder temperatures at which these animals function (Hulbert 2003). In rainbow trout, Oncorhynchus mykiss (Walbaum), it has been shown that the relative proportions of polyunsaturated fatty acids in membranes change with cold acclimation, with particularly the n-3 fatty acids increasing (Hazel 1979). The greater the degree of unsaturation of fatty acids, the more vulnerable they are to oxidative breakdown and intact antioxidant protective mechanisms, particularly presence of adequate vitamin E, are required to prevent in vivo autoxidation in bio-membranes (Niki et al. 1989). Oxidative stress associated with intense intake of polyunsaturated fat in fresh fish consumed over a short period, or due to ingestion of rancid fish remains, is likely to deplete vitamin E reserves and to result in pansteatitis in catfish as it is known to do in other animals. Vitamin E levels in rainbow trout liver were found to be inversely proportional to dietary level of lipid unsaturation, showing a higher utilisation of vitamin E associated with unsaturated lipid intake, and feeding of such diets may induce apparent vitamin E deficiency symptoms (Watanabe et al. 1981). Under conditions of dietary oxidant overload, depletion of vitamin E has also been shown to occur in muscle, liver and plasma of sharptooth catfish (Baker & Davies 1996, 1997). Where vitamin E is insufficient to provide adequate protection against the peroxidation of unsaturated lipids, necrosis and inflammation of the adipose tissues ensues, giving rise to the clinical picture of pansteatitis. Steatitis is the lesion that follows on an oxidative insult to the adipose tissues and the author has demonstrated (unpublished data) that in catfish such lesions can persist for protracted periods of time. From field measurements done by the author (unpublished data) of serum vitamin E values in catfish with pansteatitis in the Olifants Gorge, it would appear that many but not all of these fish had normal serum vitamin E values at the time of sampling, indicating that the oxidative stress exposure was not continuous. Where oxidation of lipids is not currently taking place in the adipose tissues, vitamin E levels may return to normal in animals chronically affected by pansteatitis.
Pansteatitis was confirmed in catfish from three main locations within the KNP: the Olifants Gorge and lower Letaba River at the confluence with the Olifants River, Engelhard Dam on the Letaba River upstream of the Olifants-Letaba confluence, and the Sabiepoort. Pansteatitis in crocodiles was first reported from the Olifants Gorge when large numbers of crocodiles died in this section of the Olifants River in the winter of 2008 (Ferreira & Pienaar 2011). During subsequent samplings from 2009 to 2011 an increasing prevalence of pansteatitis was observed in sharptooth catfish in the Olifants River gorge, yet only a low prevalence was found in catfish inhabiting the Olifants River upstream of the gorge, with none in catfish from a rain-filled dam not connected to the Olifants River. Common to both the Olifants Gorge and the Sabiepoort is the damming of the rivers in Mozambique to form lakes Massingir and Corumana respectively. The inlets of both lakes extend westwards back into the KNP, flooding the respective gorges where these rivers previously traversed the Lebombo Mountains as fast flowing rapids. During 2009, SANParks staff found several dead crocodiles in the Sabie River in the vicinity of the Sabiepoort with pansteatitis also found to be the cause of death (D. Govender, SANParks, Skukuza, South Africa, pers. comm., 2010). A single sampling of catfish from the Sabiepoort, where the Sabie River enters Lake Corumana on the Mozambique border, revealed pansteatitis prevalence similar to that found in the Olifants Gorge, suggesting that the aquatic environment in the two rivers is similar. As in the case of Lake Massingir, Lake Corumana supports a freshwater fishery in Mozambique. Catfish with pansteatitis may have migrated from the Olifants Gorge upstream to Engelhard Dam and this may be one explanation for the presence of pansteatitis-affected fish at this site, but the Sabiepoort has an entirely separate catchment. The anthropogenic activities resulting in potential pollution of the rivers differ greatly between these two catchments, providing argument against primary pollution-related aetiology of pansteatitis at these two sites.
Crocodile mortalities have declined in the Olifants Gorge since the mass die-off of 2008 but co-existence of old and recent lesions in sampled catfish point to an on-going incitement of pansteatitis. Analysis of catfish stomach contents in KNP strongly suggested that consumption of a predominantly fish diet within the Olifants Gorge was associated with the high prevalence of pansteatitis in catfish in this location. Catfish are primarily omnivorous benthic scavengers with wide dietary options but may form schools that actively hunt fish (Skelton 2001) and are known to feed on fish elsewhere (Spataru, Viveen & Gophen 1987). Under natural circumstances, a diet of fish may be well tolerated by sharptooth catfish (Uys 1988), but an intense intake of polyunsaturated fat over a short period is likely to deplete vitamin E reserves and to result in pansteatitis, especially if combined with other oxidative stressors. The Van Ryssen Dam collects treated wastewater from a phosphate mine, and fish from this dam showed no signs of pansteatitis despite a high prevalence of Mozambique tilapia in the stomach contents. The captive population of catfish of Lunsklip Fisheries was fed an excess of fish waste that was observed rotting in the water before being consumed. Under such conditions, pansteatitis could be expected as a result of rancidity of ingested fats leading to oxidative stress and depletion of vitamin E stores.
Tiger fish included in the preliminary samplings of fish from the Olifants Gorge during the winter of 2008, soon after the initial crocodile deaths were discovered, did not have pansteatitis. This finding was repeated during a sampling in June 2011 at a time when prevalence of pansteatitis in catfish from the gorge was high. Tiger fish, being obligate piscivorous predators, possibly evolved more effective antioxidant mechanisms to deal with a higher polyunsaturated fat intake. Being near the top of the aquatic food chain, they would be expected to develop pansteatitis if an inciting agent was bio-accumulated via the fish they kill and eat, or was assimilated from the water. The development of pansteatitis in sharptooth catfish must therefore be linked rather to their benthic scavenging habits or to a change in food source and/ or other simultaneous environmental factors to which they are not adequately adapted.
Pansteatitis in free ranging wild animals has rarely been reported except from aquatic birds. Recently an outbreak of steatitis in wild egrets and herons was reported from a reservoir in Japan (Neagari et al. 2011). However, steatitis has been reported in wild marine fish, the common dab, Limanda limanda (L.), with a suspected pollution-related aetiology (Begg, Bruno & McVicar 2000). Concern about the KNP Olifants River crocodile demise stems from the possible influence of anthropogenic effects on the Olifants River catchment, which covers some 74 500 km2 and is home to about 8% of South Africa's population (Ashton 2010). Approximately 90% of the country's saleable coal is mined in this catchment and is used to generate 55% of South Africa's electricity, resulting in serious pollution concerns (Coetzee, Du Preez & Van Vuuren 2002; De Villiers & Mkwelo 2009). The area contains numerous dams, including 38 major dams, as well as the country's second largest irrigation scheme (Anon 2001). In addition, large areas of the landscape have been changed by afforestation and agriculture. Huge increases in urban wastewater discharge and on-going high nutrient run-off from agricultural practices raise added concerns of eutrophication (Heath et al. 2010). A large phosphate mine is situated near the town of Phalaborwa just east of the KNP near the western entry point of the Olifants River into the KNP. For a number of years prior to 2004, and once in 2008, abnormally high phosphate levels were recorded in the Olifants River within the KNP (J. Venter, SANParks, Skukuza, South Africa, pers. comm., 2012). These were ascribed to the discharge of tailings from the phosphate mine into the Selati River, a tributary of the Olifants River, and to municipal sewerage discharges from the town of Phalaborwa. Such phosphate discharges would have contributed to the inorganic nutrient load trapped in sediments of Lake Massingir. Dissolved phosphate is often the limiting nutrient governing phytoplankton growth in fresh water and phosphates released from sediments will continue to drive the nutrient cycle of the lake. This seasonal stimulus for phytoplankton growth may have contributed to an increasing biomass of fish in the lake.
Contamination of surface waters in the catchment, with accumulation of heavy metals within sediments through adsorption and precipitation processes, has long been recognised as a serious pollution concern, and site specific bio-accumulation of metals has been demonstrated in sharptooth catfish in the upper catchment of the Olifants River (Coetzee et al. 2002). Oberholster et al. (2011) have suggested that aluminium and iron bio-accumulation by Mozambique tilapia in Lake Loskop may have induced the yellow fat observed from fish at that site. Baker et al. (1997) have, however, proposed that sharptooth catfish efficiently regulate iron status and are able to prevent tissue assimilation of dietary iron intake. This is an important adaptation to their benthic habitat, in which they are likely to consume sediment burrowing organisms with inadvertent ingestion of sediment. Large dams in the catchment act as traps for sediments, nitrates, phosphates and heavy metals and are regarded as the epicentre of recent mortalities of fish and crocodiles (Heath et al. 2010). Over time, changing water quality may cause sediment-bound contaminants to become bio-available and result in bio-accumulation in fish tissues.
Redox cycling of iron is known to be an initiator of lipid peroxidation (Baker & Davies 1997; Demopoulos 1973; Minotti & Aust 1992; Tappel 1973) and depletion of tissue vitamin E levels by high dietary iron intake may render polyunsaturated fats in the tissues of the fish vulnerable to peroxidation (Baker & Davies 1997). In another study (Huchzermeyer et al. 2011), pansteatitis-affected catfish caught during September of 2008 and from January to November of 2009 in the lower Letaba and Olifants Rivers were shown by special histological staining to have accumulated large amounts of iron in the form of haemosiderin in the melanomacrophages of the liver, spleen, ovary and to a lesser extent in the kidney. However, haemosiderin was not detected in the intense macrophage reaction associated with pansteatitis in the adipose tissues. Blood smears of many of these fish showed an abundance of immature erythrocytes as well as irregular erythrocyte shapes as described in cases of vitamin E deficiency in fish (Murai & Andrews 1974; Post 1993; Smith 1979) and increased erythrocyte turnover may have been the source for the increased haemosiderin carried by macrophages. Stomach contents of fish caught in the gorge at the confluence yielded mainly remnants of large fish, whereas specimens caught directly over the silt deposits on the Mozambique border only contained detritus and silt. These findings suggested bio-accumulated iron as an additional oxidative trigger possibly ingested in polluted sediment. As stated before, however, catfish are able to prevent tissue assimilation of dietary iron (Baker et al. 1997). This and the absence of haemosiderin in macrophages associated with pansteatitis in the adipose tissues of catfish from the Olifants Gorge thus suggest that the role of iron in the aetiology of pansteatitis in catfish in the Olifants Gorge remains speculative.
As a consequence of raising the dam of Lake Massingir, the heavy silt load of the Olifants River has been deposited in flooded parts of the narrow gorge extending into the KNP. The now inundated sand-bottomed pools between previously fast flowing rapids once formed a favoured habitat for large crocodiles. This altered habitat in the Olifants Gorge may have favoured access to certain fish species not normally consumed in large numbers by crocodiles and catfish. Such species may provide higher levels of polyunsaturated fatty acids than those to which the animals are adapted. An increase in dietary polyunsaturated fat intake has been reported to result in pansteatitis in various animals. Wallach and Hoessle (1968) concluded that a change in diet from smelt (6.7% fat) to mackerel (29.9% fat) was the precipitating cause of pansteatitis in captive American alligators, Alligator mississippiensis (Daudin). Goodwin (2006) stressed the dangers of using diets high in fish oils for inappropriate species, and a change from Baltic and Mediterranean clupeids to Moroccan Atlantic pilchards was suspected to have been the cause of pansteatitis in northern bluefin tuna, Thunnus thynnus (L.), reported by Roberts and Agius (2008). Similarly pansteatitis could be induced in cats by feeding an oil-rich fish-based diet (Fytianou et al. 2006).
Silver carp, Hypophthalmichthys molitrix (Valenciennes), an alien invasive schooling species from East Asia (Kolar et al. 2005), were introduced into Mozambique from Cuba and also escaped into the Olifants River from South Africa. They are known to occur in Lake Massingir (Skelton 2001) and have been observed on occasion in the Olifants River in large numbers (J. Venter, SANParks, Skukuza, South Africa, pers. comm., 2012). This fish is a specialised plankton feeder that by preference feeds off phytoplankton and is an important consumer of cyanobacterial blooms (Kolar et al. 2005), a niche no indigenous South African fish species occupies (P. Skelton, South African Institute of Aquatic Biodiversity, Grahamstown, South Africa, pers. comm., 2012). Such blooms have been observed near the inlet to Lake Massingir (D. Pienaar, SANParks, Skukuza, South Africa, pers. comm., 2009). Phytoplankton naturally contains large quantities of α-linolenic acid and other n-3 polyunsaturated fatty acids, in particular eicosapentaenoic acid C20:5n-3 (EPA) and docosahexaenoic acid C22:6n-3 (DHA) (Steffens 1997). Intake of these fatty acids is reflected in the adipose tissues of silver carp. In one study, these two fatty acids were found to constitute up to 5.28% and 3.41% of body fat triacylglycerols respectively (Buchtová & Jezek 2011). The n-6 and n-3 fatty acids derived from linoleic and α-linolenic acids respectively are essential fatty acids that cannot be synthesised by animals (Steffens 1997). The relative abundance of these fatty acids in the diet of animals is reflected in the composition of their fat tissues (Hoffman & Prinsloo 1995; Steffens 1997). Compared with the fat of farmed crocodiles, a much higher intake of n-3 fatty acids was reflected in the fat of crocodiles in the Olifants Gorge (Osthoff et al. 2010). In another study, Huchzermeyer et al. (in press) demonstrated that the mesenteric fat of catfish with pansteatitis from the Olifants Gorge showed a similarly high inclusion of n-3 fatty acids, whereas mesenteric fat of healthy catfish from the same site reflected a lower inclusion of n-3 fatty acids.
Seasonal spawning migration of silver carp out of still water bodies into fast flowing rivers occurs over an 8-10 week period once rivers reach their peak summer flows (Kolar et al. 2005). During this time, the fish congregate in large numbers and become easy prey for crocodiles and catfish. In the Olifants Gorge this would occur from January to March, a time when fish in the river are difficult to monitor (A. Deacon, SANParks, Skukuza, South Africa, pers. comm., 2012). This may provide an explanation for intense dietary exposure to polyunsaturated fats that could have led to development of obesity and pansteatitis in crocodiles in the Olifants Gorge during the winter of 2008 and to a lesser extent during subsequent winters and to the on-going prevalence of pansteatitis in sharptooth catfish.
The only recorded catfish die-off in the Olifants Gorge was observed during the winter of 2009. The major crocodile die-offs in the Olifants Gorge occurred in 2008 and 2009 respectively and to a lesser extent in subsequent years, after the first really cold weather was experienced around the end of May each year. In many aquatic poikilotherms, especially fish, acclimation to colder water temperatures in winter involves an increase in membrane polyunsaturated fatty acids, in particular DHA (Hazel 1979; Hulbert 2003).
Where antioxidant mechanisms have been depleted through oxidative stress, such as that produced by intense dietary polyunsaturated fat intake, compromised bio-membranes in critical tissues might result in impairing adaptation to cold. In an unpublished study the author has demonstrated, however, that catfish severely affected by pansteatitis have survived protracted periods at water temperatures as low as 12 °C with no undue effect. This is well below the lowest water temperature experienced in the Olifants Gorge during winter. Pansteatitis is largely a foreign-body type reaction following on breakdown of fat cells, and presence of the lesion may imply either continuous or preceding oxidative stress. The effect of pansteatitis on cold tolerance may therefore differ depending on the time that has lapsed since the oxidative insult took place.
Conclusion
The deaths of a large number of Nile crocodiles, a keystone species in aquatic conservation, in the Olifants Gorge in the KNP, has raised serious questions about the consequences of anthropogenic activity resulting in altered hydrodynamics and pollution in the catchment of the Olifants River. The objective of this study was to investigate the occurrence of pansteatitis in catfish inhabiting the same waters in the KNP where crocodiles had died of pansteatitis. Catfish were sampled repeatedly over a two-year period from the Olifants Gorge as well as from other sites in and around KNP. In the KNP, pansteatitis in both catfish and crocodiles has been observed in areas where the natural habitat has been drastically altered as a result of damming of rivers, and where the associated deposition of large clay deposits is a potential source of pollution. The increasing prevalence of pansteatitis in catfish in the Olifants Gorge, and the accumulation of lesions over time, points to periodic or seasonal episodes of dietary oxidative stress in these animals. Pansteatitis has also been identified in catfish at other sites in the KNP. The catchment areas feeding these sites differ from that of the Olifants River, providing argument against a primary pollution-related oxidative stress. To date, the only factor common to both the Olifants Gorge and the Sabiepoort, both epicentres of crocodile mortality, and both sites where pansteatitis has been identified in catfish, is the extension of man-made lake inlets into areas favoured by crocodiles. Increasing phosphate levels from anthropogenic activities upstream and in the catchment area of the Olifants River have led to an increase in phytoplankton blooms in Lake Massingir. Whereas this suggests that hydrodynamic change and pollution are the main drivers of this condition in the Olifants Gorge, the presence of large schools of the invasive alien silver carp, benefiting from the nutrient-rich raised water level of Lake Massingir, and known for its high content of n-3 polyunsaturated fatty acids, likely formed much of the seasonal diet of the catfish and crocodiles, either alive or as dead remains, and are thus proposed as the cause of the obesity and a pansteatitis-initiating factor in these animals. The role of bioaccumulation of iron from polluted sediments in initiating lipid autoxidation in catfish is speculative and needs further investigation. The effects on poikilotherm membranes and metabolic rate of a sudden seasonal drop in environmental temperature may have contributed to the die-off of the pansteatitis-affected crocodiles, yet catfish survive cold even when severely affected by pansteatitis. It is not yet clear whether silver carp occur in Lake Corumana and further studies are needed to verify these proposals and answer outstanding questions.
Acknowledgements
The assistance of SANParks and in particular Mr D.J. Pienaar, Drs D. Govender and A.R. Deacon as well as Mr J. Venter and the SANParks rangers and fishermen is gratefully acknowledged. Mss P. Shikwambana and N. Tyaqana, and Mr K. Deenadayalan of SANParks are thanked for help with the field laboratory. Messrs C., R., and N. Huchzermeyer are thanked for assistance with catching and handling fish and with field logistics. Mr J. Muller and IDEXX laboratory and Drs J. Myburgh and J. Steyl from the Department of Paraclinical Sciences at the Faculty of Veterinary Science, University of Pretoria are thanked for assistance with sampling. Dr Steyl is thanked for preparing histological sections and special stains. Mr T. Boikhutlo and Ms G. Taylor from the Department of Ichthyology and Fisheries Science, Rhodes University are thanked for preparing otolith sections. Prof. J. Lawrence is thanked for scientific and editorial comments. Dr P.A. Colly is thanked for help with sampling and logistic, editing and financial support. The assistance by Lunsklip Fisheries is gratefully acknowledged. Mr M. Lotter is thanked for preparing the map of the sampling sites. Colleagues from Sterkspruit Veterinary Clinic are thanked for standing in during the author's absence with fieldwork.
Financial assistance for this study was provided by WRC consultancy contract K8/948 and by the South African Veterinary Foundation. The project was approved by SANParks and the Animal Use and Care Committee of the University of Pretoria under Protocol VO13/10.
Competing interests
The author declares that he has no financial or personal relationship(s) which may have inappropriately influenced him in writing this paper.
References
Anon, 2001, State of the rivers report: Crocodile, Sabie-Sand and Olifants River systems, 2001, viewed 2011, from http://www.dwaf.gov.za/iwqs/rhp/state_of_ rivers/crocsabieolif_01_toc.html [ Links ]
Ashton, P.J., 2010, 'The demise of the Nile crocodile (Crocodylus niloticus) as a keystone species for aquatic ecosystem conservation in South Africa: The case of the Olifants River', Aquatic Conservation: Marine and Freshwater Ecosystems 20, 489-493. http://dx.doi.org/10.1002/aqc.1132 [ Links ]
Baker, R.T.M. & Davies, S.J., 1996, 'Oxidative nutritional stress associated with feeding rancid oils to African sharptooth catfish, Clarias gariepinus (Burchell) and the protective role of α-tocopherol', Aquaculture Research 27, 795-803.http://dx.doi.org/10.1046/j.1365-2109.1996.t01-1-00814.x [ Links ]
Baker, R.T.M. & Davies, S.J., 1997, 'Muscle and hepatic fatty acid profiles and α-tocopherol status in African catfish (Clarias gariepinus) given diets varying in oxidative state and vitamin E inclusion level', Animal Science 64, 187-195. http://dx.doi.org/10.1017/S1357729800015708 [ Links ]
Baker, R.T.M., Martin, P. & Davies, S.J., 1997, 'Ingestion of sub-lethal levels of iron sulphate by African catfish affects growth and tissue lipid peroxidation', Aquatic Toxicology 40, 51-61. http://dx.doi.org/10.1016/S0166-445X(97)00047-7 [ Links ]
Bainy, A.C.D., Saito, E., Carvalho, P.S.M. & Junqueira, V.B.C., 1996, 'Oxidative stress in gill, erythrocytes, liver and kidney of Nile tilapia {Oreochromis niloticus) from a polluted site', Aquatic Toxicology 34, 151-162. http://dx.doi.org/10.1016/0166-445X{95)00036-4 [ Links ]
Begg, G.S., Bruno, D.W. & McVicar, A.H., 2000, 'The histopathology and ultrastructure of steatitis affecting common dab Limanda limanda', Diseases of Aquatic Organisms 41, 123-133. http://dx.doi.org/10.3354/dao041123, PMid:10918980 [ Links ]
Botha, H., Van Hoven, W. & Guillette, L.J., 2011, 'The decline of the Nile crocodile population in Loskop Dam, Olifants River, South Africa', Water SA 37, 103-108. http://dx.doi.org/10.4314/wsa.v37i1.64109 [ Links ]
Buchtová, H. & Jezek, F., 2011, 'A new look at the assessment of silver carp {Hypophthalmichthys molitrix Val.) as a food fish', Czech Journal of Food Science 29, 487-497. [ Links ]
Coetzee, L., Du Preez, H.H. & Van Vuuren, J.H.J., 2002, 'Metal concentrations in Clarias gariepinus and Labeo umbratus from the Olifants and Klein Olifants River, Mpumalanga, South Africa: Zinc, copper, manganese, lead, chromium, nickel, aluminium and iron', Water SA 28, 443-448. http://dx.doi.org/10.4314/wsa.v28i4.4917 [ Links ]
Demopoulos, H.B., 1973, 'Control of free radicals in biologic systems', Federation Proceedings 32, 1903-1908. PMid:4718907 [ Links ]
De Villiers, S. & Mkwelo, S.T., 2009, 'Has monitoring failed the Olifants River, Mpumalanga?', Water SA 35, 671-676. http://dx.doi.org/10.4314/wsa.v35i5.49193 [ Links ]
Ferreira, S.M. & Pienaar, D., 2011, 'Degradation of the crocodile population of Kruger National Park, South Africa', Aquatic Conservation: Marine and Freshwater Ecosystems 21, 155-164. http://dx.doi.org/10.1002/aqc.1175 [ Links ]
Fytianou, A., Koutinas, A.F., Saridomichelakis, M.N. & Koutinas, C.K., 2006, 'Blood α-tocopherol, selenium, and glutathione peroxidase changes and adipose tissue fatty acid changes in kittens with experimental steatitis {yellow fat disease): A comparative study between the domestic shorthaired and Siamese breed', Biological Trace Element Research 11, 131-143. http://dx.doi.org/10.1385/BTER:112:2:131 [ Links ]
Goodwin, A.E., 2006, 'Steatitis, fin loss and skin ulcers of channel catfish, Ictalurus punctatus (Rafinesque), fingerlings fed salmonid diets', Journal of Fish Diseases 29, 61-64. http://dx.doi.org/10.1111/j.1365-2761.2005.00686.x, PMid:16351699 [ Links ]
Hazel, J.R., 1979, ' Influence of thermal acclimation on membrane lipid composition of rainbow trout liver', American Journal of Physiology - Regulatory, Integrative and Comparative Physiology 236, 91-101. [ Links ]
Heath, R., Coleman, T. & Engelbrecht, J., 2010, 'Water quality overview and literature review of the ecology of the Olifants River', WRC Report No. TT452/10. [ Links ]
Herman, R.L. & Kircheis, F.W., 1985, 'Steatitis in Sunapee trout, Salvelinus alpinus oquassa Girard', Journal of Fish Diseases 8, 237-239. http://dx.doi.org/10.1111/j.1365-2761.1985.tb01219.x [ Links ]
Hoffman, L.C. & Prinsloo, J.F., 1995, 'Genetic and nutritional influence on the total lipid fatty acid profile of Clarias gariepinus muscle', Aquatic Living Resources 8, 415-121. http://dx.doi.org/10.1051/alr:1995049 [ Links ]
Huchzermeyer, F.W., 2003, Crocodiles: Biology, husbandry and diseases, CABI Publishing, Wallingford. http://dx.doi.org/10.1079/9780851996561.0000 [ Links ]
Huchzermeyer, K.D.A., Govender, D., Pienaar, D.J. & Deacon, A.R., 2011, 'Steatitis in wild sharptooth catfish, Clarias gariepinus (Burchell), in the Olifants and Lower Letaba Rivers in the Kruger National Park, South Africa', Journal of Fish Diseases 34, 389-398. http://dx.doi.org/10.1111/j.1365-2761.2011.01267.x, PMid:21675995 [ Links ]
Huchzermeyer, K.D.A., Osthoff, G., Hugo, A. & Govender, D., in press, 'Comparison of the lipid properties of healthy and pansteatitis-affected African sharptooth catfish, Clarias gariepinus (Burchell), and the role of diet in pansteatitis outbreaks in the Olifants River in the Kruger National Park, South Africa', Journal of Fish Diseases, (accepted for publication 31 August 2012). [ Links ]
Hulbert, A.J., 2003, 'Life, death and membrane bilayers', Journal of Experimental Biology 206, 2303-2311. http://dx.doi.org/10.1242/jeb.00399, PMid:12796449 [ Links ]
Kelly, S.A., Havrilla, C.M., Brady, T.C., Abramo, K.H. & Levin, E.D., 1998, 'Oxidative stress in toxicology: Established mammalian and emerging piscine model systems', Environmental Health Perspectives 106, 375-384. http://dx.doi.org/10.1289/ehp.98106375, PMid:9637794 [ Links ]
Kibanova, D., Nieto-Camacho, A. & Cervini-Silva, J., 2009, 'Lipid peroxidation induced by expandable clay minerals', Environmental Science and Technology 43, 75507555. http://dx.doi.org/10.1021/es9007917 [ Links ]
Kolar, C.S., Chapman, D.C., Courtenay, Jr. W.R., Housel, C.M., Williams, J.D. & Jennings, D.P., 2005, 'Asian carps of the genus Hypophthalmychthys (Pisces, Cyprinidae) -A biological synopsis and environmental risk assessment', Report to US Fish and Wildlife Service per Interagency Agreement 94400-3-0128, viewed 2012, from http://www.fws.gov/contaminants/OtherDocuments/ACBSRAFinalReport2005.pdf [ Links ]
Ladds, P.W., Mangunwirjo, H., Sebayang, D. & Daniels, P.W., 1995, 'Diseases in young farmed crocodiles in Irian Jaya', Veterinary Record 136, 121-124. http://dx.doi.org/10.1136/vr.136.5.121, PMid:7740731 [ Links ]
Minotti, G. & Aust, S.D., 1992, 'Redox cycling of iron and lipid peroxidation', Lipids 27, 219-226. http://dx.doi.org/10.1007/BF02536182, PMid:1326072 [ Links ]
Murai, T. & Andrews, J.W., 1974, 'Interactions of dietary α-tocopherol, oxidized menhaden oil and ethoxyquin on channel catfish (Ictalurus punctatus}', Journal of Nutrition 104, 1416-1431. PMid:4423571 [ Links ]
Neagari, Y., Arii, S., Udagawa, M., Onuma, M., Odaya, Y., Kawasaki, T. et al., 2011, 'Steatitis in egrets and herons from Japan', Journal of Wildlife Diseases 47, 49-55. PMid:21269996 [ Links ]
Niki, E., Yamamoto, Y., Takahashi, M., Komuru, E. & Miyama, Y., 1989, 'Inhibition of oxidation of biomembranes by tocopherol', in A.T. Diplock, L.J. Machlin, L. Packer & W.A. Pryor (eds.), Vitamin E: Biochemistry and health implications, pp. 23-31, Annals of the New York Academy of Sciences 570. [ Links ]
Oberholster, P.J., Myburgh, J.G., Ashton, P.J., Coetzee, J. & Botha, A.-M., 2011, 'Bioaccumulation of aluminium and iron in the food chain of Lake Loskop, South Africa', Ecotoxicology and Environmental Safety 75, 134-141. http://dx.doi.org/10.1016/j.ecoenv.2011.08.018, PMid:21924494 [ Links ]
Osthoff, G., Hugo, A., Bouwman, H., Buss, P., Govender, D., Joubert, C.C. et al., 2010, 'Comparison of the lipid properties of captive, healthy wild, and pansteatitis affected wild Nile crocodiles (Crocodylus niloticus}', Comparative Biochemistry and Physiology Part A: Molecular and Integrative Physiology 155, 64-69. http://dx.doi.org/10.1016/j.cbpa.2009.09.025 [ Links ]
Post, G.W., 1993, 'Nutrition and nutritional diseases of salmonids', in M.K. Stoskopf (ed.), Fish Medicine, pp. 343-358, W.B. Saunders Company, Harcourt Brace Jovanovich, Philadelphia, London, Toronto, Montreal, Sydney, Tokyo. [ Links ]
Roberts, R.J. & Agius, C., 2008, 'Pan-steatitis in farmed northern bluefin tuna, Thunnus thynnus (L.), in the eastern Adriatic', Journal of Fish Diseases 31, 83-88. http://dx.doi.org/10.1111/j.1365-2761.2007.00901.x, PMid:18234015 [ Links ]
Roberts, R.J., Richards, R.H. & Bullock, A.M., 1979, 'Pansteatitis in rainbow trout Salmo gaidneri Richardson: A clinical and histological study', Journal of Fish Diseases 2, 85-92. http://dx.doi.org/10.1111/j.1365-2761.1979.tb00145.x [ Links ]
Skelton, P., 2001, A complete guide to the freshwater fishes of Southern Africa, Struik Publishers, Cape Town. [ Links ]
Spataru, P., Viveen, W.J.A.R. & Gophen, M., 1987, 'Food composition of Clarias gariepinus (= C. lazera) (Cypriniformes, Clariidae) in Lake Kinneret (Israel)', Hydrobiologica 144, 77-82. http://dx.doi.org/10.1007/BF00008053 [ Links ]
Smith, C.E., 1979, 'The prevention of liver lipoid degeneration (ceroidosis) and microcytic anaemia in rainbow trout Salmo gairdneri Richardson fed rancid diets: A preliminary report', Journal of Fish Diseases 2, 429-437. http://dx.doi.org/10.1111/j.1365-2761.1979.tb00394.x [ Links ]
Steffens, W., 1997, 'Effects of variation in essential fatty acids in fish feeds on nutritive value of freshwater fish for humans', Aquaculture 151, 97-119. http://dx.doi.org/10.1016/S0044-8486(96)01493-7 [ Links ]
Tappel, A.L., 1973, 'Lipid peroxidation damage to cell components', Federation Proceedings 32, 1870-1874. PMid:4352451 [ Links ]
Uys, W., 1988, 'Nutrition', in T. Hecht, W. Uys & P.J. Britz (eds.), The culture of sharptooth catfish, Clarias gariepinus in southern Africa, South African National Scientific Programmes Report no. 153, pp. 47-61, CSIR, Pretoria. [ Links ]
Wallach, J.D. & Hoessle, C., 1968, 'Steatitis in captive crocodilians', Journal of the American Veterinary Medical Association 153, 845-847. PMid:5692919 [ Links ]
Watanabe, T., Takeuchi, T., Wada, M. & Uehara, R., 1981, 'The relationship between dietary lipid levels and α-tocopherol requirement of rainbow trout', Bulletin of the Japanese Society of Scientific Fisheries 47, 1463-1471.http://dx.doi.org/10.2331/suisan.47.1463 [ Links ]
Weyl, O.L.F. & Booth A.J., 2008, 'Validation of annulus formation in otoliths of a temperate population of adult African sharptooth catfish Clarias gariepinus using fluorochrome marking of wild fish', Journal of Fish Biology 73, 1033-1038. http://dx.doi.org/10.1111/j.1095-8649.2008.01975.x [ Links ]
 Correspondence to:
Correspondence to:
David Huchzermeyer
PO Box 951
Lydenburg 1120, South Africa
aquavet@telkomsa.net
Received: 19 Jan. 2012
Accepted: 08 Aug. 2012
Published: 09 Nov. 2012


{kind=link}
{kind=link}
{kind=link}