










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.83 n.1 Pretoria Jan. 2012
ORIGINAL RESEARCH
Preliminary evaluation of selected minerals in liver samples from springbok (Antidorcas marsupialis) from the National Zoological Gardens of South Africa
Khanyisile R. MbathaI; Emily P. LaneI; Michael LanderII; Adrian S.W. TordiffeI; Sandra CorrIII
IDepartment of Research and Scientific Services, National Zoological Gardens of South Africa, South Africa
IIRoyal Veterinary College, Hertfordshire, United Kingdom
IIIDivision of Surgery, University of Nottingham, United Kingdom
ABSTRACT
Limited information is available on the mineral nutrition of captive antelope in South Africa. Zoo animals are usually offered a very limited array of feeds, which may result in nutritional imbalances. As a pilot study to investigate the presence of myopathy in antelope at the National Zoological Gardens of South Africa (NZG), stored liver samples from six springbok (Antidorcas marsupialis) and seven other antelopes from the NZG, as well as selected food items, were submitted for analysis of selenium, copper, manganese and zinc content by spectrophotometry. Springbok liver levels of copper were 23.07 mg/kg ± 0.72 mg/kg, whilst manganese, selenium and zinc levels were 6.73 mg/kg ± 0.22 mg/kg, 0.14 mg/kg ± 0.05 mg/kg and 135.02 mg/kg ± 1.26 mg/kg, respectively. Liver mineral levels in the other species were very variable. Food item copper levels ranged from 4.00 mg/kg (Eragrostis tef) to 17.38 mg/kg (antelope cubes), lucerne (Medicago sativa) and E. tef contained no detectable selenium. The highest zinc levels were in antelope cubes (147.00 mg/kg) and the lowest were in lucerne (20.80 mg/kg). Interpretation of these results was hampered by the small number of samples and a paucity of information on liver mineral levels in free-ranging and captive antelope; however, results suggested that, in the springbok, whilst copper and manganese intake are likely adequate, selenium nutrition is probably insufficient and may account for the myopathy diagnosed. Zinc liver levels are possibly within the toxic range, perhaps as a result of the high levels of zinc in the antelope cubes. This pilot study highlighted the need to establish baseline mineral nutrition data for captive and free-ranging antelope under South African conditions.
Introduction
In zoo animals, nutritional requirements play an important role in the health of each individual and the overall health of the population (Miller et al. 2001). Although the available energy and protein of feeds are vital to any animal, optimal health is achieved only if there is a sufficient supply of minerals (McDowell 1985). Limited knowledge is available on mineral requirements of wildlife and zoo animals (Johnson, Bleich & Krausman 2007; Zimmerman et al. 2008), complicating the diagnosis of mineral imbalances. Most wildlife and zoo nutrition is extrapolated from research in domestic animals (Robbins 1993), which may not necessarily be appropriate and should therefore be used with caution, because nutritional requirements of domestic animals may not represent the specific needs of zoo animals (Miller et al. 2001). In South Africa, investigations on the mineral status of domestic animals have been conducted since the early 19th century but none have been published regarding zoo animals. In other countries, the mineral status of zoo animals has been evaluated in black and white ruffed lemur (Varecia variegata) (Crawford et al. 2009), black rhinoceros (Diceros bicornis) (Clauss et al. 2007; Dierenfeld et al. 2005), mule and white-tailed deer (Odocoileus hemionus, Odocoileus virginianus) (Zimmerman et al. 2008) and pronghorn (Antilocapra americana) (Miller et al. 2001). The mineral status of free-ranging wildlife has been measured in African buffalo (Syncerus caffer) (Webb et al. 2001), black wildebeest (Connochaetes gnou) (Penrith et al. 1996), blesbok (Damaliscus pygargus phillipsi) (Penrith et al. 1996; Quan 2005; Quan, Mülders & Meltzer 2002), Kafue lechwe (Kobus leche kafuensis) (Syakalima et al. 2000), springbok (Antidorcas marsupialis) (Hoon 2003) and Thompson's gazelle (Eudorcas thomsonii) (Wieckowski, Abraham & Wernery 2008). Evidence suggests that free-ranging species instinctively select forages that will balance some of their mineral requirements (McNaughton 1990). However, zoo animals are usually offered a very limited array of feeds, which, together with regional variations in feed quality, may result in nutritional imbalances (Miller et al. 2001). The diets generally offered to zoo animals include hay or pelleted feed items or fresh produce and browse. Sometimes zoo animals may be fed mineral supplements and, consequently, tissue levels may not reflect the mineral concentrations of the dietary items only. Thus, mineral assays on tissue samples and feedstuffs can be used to establish the mineral nutritional status of zoo animals.
Between 2003 and 2005, eight antelope from the National Zoological Gardens of South Africa (NZG) died with pathological evidence of skeletal muscle myopathy and in two of these - a scimitar horned oryx (Oryx dammah) and a steenbok (Raphicerus campestris) - the myopathy was considered to be the cause of death. Histological evidence of the cause of the myopathy was absent in all cases, but causes found in the literature include inherited disorders, nutritional myopathy, toxic myopathy and exertional myopathy (Van Vleet & Valentine 2007). Because they are inherited, toxic and exertional myopathies were thought to be inconsistent with the epidemiology of the cases; nutritional myopathy was thought to be the most likely cause.
To investigate mineral nutrition, stored liver samples obtained during the necropsy of antelope that died at the NZG during 2007 and selected food items were submitted for analysis of selected minerals by spectrophotometry. The objective of this article is to report on liver levels of selenium (Se), copper (Cu), manganese (Mn), and zinc (Zn) in six NZG springbok and individuals of seven other species for comparison, as well as the levels of these minerals in food items. Copper, Mn, Se and Zn were selected because nutritional deficiencies in these minerals can be tested for using liver samples (Puls 1994).
Materials and methods
Subjects
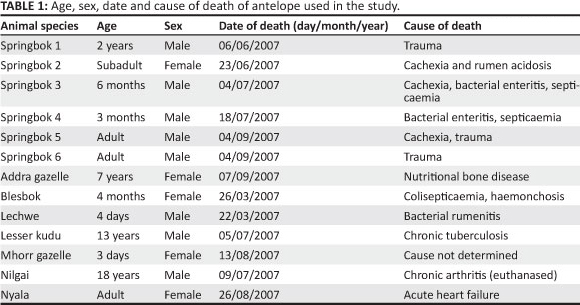
Since 2007, a post-mortem examination has taken place on every animal that dies or is euthanased as a result of disease or injury at the NZG. Liver, kidney and ribs are harvested and stored at -80 °C at the Wildlife Biological Resource Centre (wBRC) at the NZG. Every frozen antelope liver sample submitted to the wBRC between March 2006 and September 2007 was used in this study. The liver samples available included six samples from springbok and one each from seven other antelope (addra gazelle [Nanger dama], blesbok, lechwe, lesser kudu [Ammelaphus imberbis], Mhorr gazelle [Nanger dama mhorr], nilgai [Boselaphus tragocamelus] and nyala [Nyala angasii]) for comparison. Table 1 shows the age, sex, date and cause of death of all 13 antelope.
Diet samples
Diet composition of antelope at the NZG varies seasonally, but consistent food components include antelope cubes (Epol Feeds, Westville, South Africa), Boskos cubes (WES Enterprises, Pty, Ltd, Thabazimbi, South Africa), Eragrostis tef grass hay and Medicago sativa (lucerne or alfalfa) hay. The relative dietary proportions of these components for the eight species in this study are shown in Table 2.
Laboratory analyses
Liver samples from six springbok and seven other antelope were thawed at room temperature for 24 h. Liver mineral concentrations were determined at the University of Pretoria Nutrilab by the standard AOAC technique using a GBC 905AA atomic absorption spectrophotometer (GBC Scientific Equipment, Braeside, Australia) for Cu and Zn, a PerkinElmer 2380 atomic absorption spectrophotometer (PerkinElmer, Waltham, USA) for Se and a Varian Spectra AA 50 atomic absorption spectrophotometer (SpectraLab Scientific Inc., Palo Alto, USA) for Mn (AOAC 2000).
Statistical analyses
Means and standard errors were calculated for dietary parameters of teff, antelope cubes and Boskos cubes and mineral levels for the six springbok. Only one sample from the other seven animal species was available at the time of the study, so no statistical analysis could be conducted on these species.
Ethical considerations
This project was approved by the NZG Ethics and Scientific Committee (P06/19).
Results
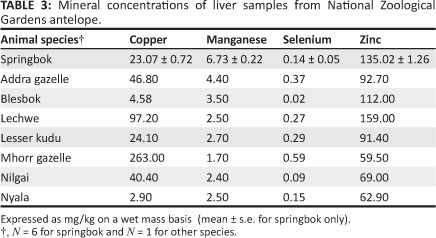
Liver mineral levels are shown in Table 3. Copper levels ranged from 2.90 mg/kg (nyala) to 263.00 mg/kg (Mhorr gazelle), with levels in springbok being 23.07 mg/kg ± 0.72 mg/kg (mean ± s.e.). Manganese levels were the highest in the springbok (6.73 mg/kg ± 0.22 mg/kg), with the lowest Mn (1.70 mg/kg) found in the Mhorr gazelle. The highest Se levels were found in the Mhorr gazelle (0.59 mg/kg), the lowest in the blesbok (0.02 mg/kg) and the levels in the springbok were 0.14 mg/kg ± 0.05 mg/kg. Zinc levels were highest in the lechwe (159.00 mg/kg), lowest in the Mhorr gazelle (59.50 mg/kg) and springbok levels were 135.02 mg/kg ± 1.26 mg/kg.
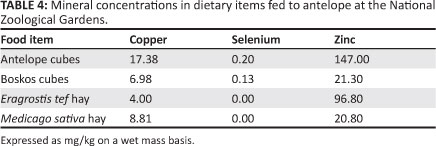
Food item Cu levels ranged from 4.00 mg/kg (£. tef) to 17.38 mg/kg (antelope cubes, Table 4). Medicago sativa and E. tef hay contained no detectable Se. The highest Zn levels were in antelope cubes (147.00 mg/kg) and the lowest were in lucerne (20.80 mg/kg).
Trustworthiness
The Agri-Laboratory Association of Southern Africa recently acknowledged the accurate laboratory analysis undertaken by the University of Pretoria Nutrilab with a prestigious award (http://web.up.ac.za/sitefiles/file/48/Squared%20UP%20May2012_English.pdf). However, because of the small sample numbers available for this pilot study, the results presented here need to be interpreted with caution and studies with larger sample size need to be conducted.
Discussion
In the present study, liver mineral levels were compared with values given in the few published reports. However, it is not always clear in the previous studies whether the animals were captive or free-ranging and methods of sample collection and analysis vary. For example, in the two springbok references used for comparison with this study, one springbok was included as part of a study involving blesbok (Penrith et al. 1996) and a large study of springbok is reported in which sample preparation method was not described (Van Ryssen & Hoon 2010) (Table 5). Domestic animals, such as cattle, sheep and goats, may also be fed mineral supplements and consequently tissue levels may not reflect the position with regard to natural herbage. Therefore, analyses of tissues from free-ranging herbivores, such as the African buffalo, may be valuable in establishing a baseline mineral status of the animals and may be used to monitor changes over time (McDonald et al. 2002). The NZG is in the process of developing a tissue bank from captive and free-ranging animals that could be used for this purpose, as large numbers of antelope are kept commercially in South Africa under captive or semi-captive conditions.
Copper
The liver is the primary Cu storage organ (Brewer 1987) and accounts for approximately 10% of total amount of Cu in the body (Evans 1973). Hepatic Cu concentration increases linearly with Cu intake and is a good measure of the long-term Cu status of the ruminant because of the degradation of the Cu complexes in the rumen facilitating absorption (Du, Hemken & Harmon 1996). Recorded concentrations of springbok liver Cu were 16.02 mg/kg (Penrith et al. 1996) and 35.00 mg/kg (Van Ryssen & Hoon 2010). For comparison, the literature value based on dry matter was divided by 3.3 because it has been assumed that a liver sample has a mean dry matter content of 30% (Long 1961). As referenced, springbok liver mineral levels are scant, they were also compared with the blackbuck (Antílope cervicapra), which inhabit similar environments and have similar feeding and social habits (Burton & Burton 2002). According to Puls (1994), the normal liver Cu concentration is 50.00 mg/kg -100.00 mg/kg for blackbuck, thus the springbok liver Cu concentrations in the present study are probably within the normal range.
The single nilgai (Puls 1994) and blesbok (Penrith et al. 1996; Quan et al. 2002) Cu levels were considerably lower than those recorded in the literature, but the liver Cu level in the addra gazelle was almost identical to that recorded in other captive gazelles (Wieckowski et al. 2008). However, the liver Cu level in the 3-day-old Mhorr gazelle was much higher than that recorded in Thomson's gazelle. This discrepancy could be caused by species differences in Cu metabolism or to the young age of the Mhorr gazelle (Table 1), because liver Cu levels may be higher in neonates (Puls 1994).
In the present study, the dietary Cu in feed items fed to the antelope were within the recommended range of 4 mg/kg -15 mg/kg (Pugh 2002; Smith & Sherman 2009; Underwood 1977), except for the antelope cubes, which were slightly higher than recommended (Table 4), perhaps the result of high Cu levels in the ingredients from which the pellets were prepared (Ange et al. 2001).
Dietary concentrations of minerals do not necessarily determine the subsequent tissue concentrations caused by the form of the mineral interactions (such as with tannins or other minerals or vitamins) that prevent absorption and differential organ accumulation of minerals (Penrith et al. 1996). Liver Cu concentration increases linearly with Cu intake, whilst serum and plasma Cu increase at the low range and then remain constant, except if affected by high molybdenum (Mo) and sulphur (S) intakes. The interaction between Cu, Mo and S is of great importance for health and reproduction in ruminants. Copper and Mo are interrelated in animal metabolism and should be considered together (Hennig et al. 1974). In the current study, Mo and S were not ascertained, so no final conclusion about the adequacy of Cu nutrition in the springbok and other NZG antelopes was possible. Zinc levels were high in the antelope cubes, and Zn also antagonises Cu accumulation, therefore reducing availability, and vice versa. Liver Cu levels may also be influenced by dietary protein (Clauss & Dierenfeld 1999), but these were also not evaluated in this study.
Manganese
Most tissues contain traces of Mn and it is important in animals as an activator of many enzymes. The highest concentrations occur in the bones, pituitary gland, kidney, pancreas and liver when ingested in high amounts. Bone Mn is more responsive to dietary intake than other tissues.
Liver Mn concentrations of springbok and other antelope at the NZG were within the normal range (2.0 mg/kg -6.0 mg/kg [Puls 1994], Table 3), except in the juvenile Mhorr gazelle, which had slightly lower liver Mn (1.7 mg/kg). These results suggest that NZG antelope Mn nutrition is likely adequate. Deficiency of Mn in grazing ruminants is rare but causes lameness, shortening and bowing of the legs and enlarged joints in cattle, goats and sheep (McDonald et al. 2002). These conditions were not observed in the animals in the present study, although the addra gazelle died of nutritional bone disease more characteristic of calcium (Ca) and/or phosphorus (P) malnutrition (Table 1). However, it should be emphasised that the Mn concentration in the liver is a poor indicator of the Mn status of the animals, except in situations of abnormally high or low intake (McDonald et al. 2002).
Green plant material contains adequate amounts of Mn. Most forage contains 40 mg/kg - 200 mg/kg dry matter (DM) of Mn and there is a wide margin of safety between the toxic and the normal levels of dietary Mn (McDonald et al. 2002). Excess dietary intake of Ca and P reduces Mn absorption and iron deficiency increases Mn absorption. These minerals were not measured in this study, but Mn levels appear adequate.
Selenium
Selenium is one of the few elements where concentrations in the liver and whole blood are positively correlated with dietary Se intake up to fairly high intakes. Serum Se is considered as an indication of short-term Se intake, whilst whole blood and liver levels reflect long-term Se intake. Selenium has a sparing effect on Vitamin E levels by ensuring normal absorption of the vitamin. Vitamin E and Se assist in the immune system and protect against heavy metal toxicity (McDonald et al. 2002).
Liver Se levels varied considerably between species in the study, with that of the Mhorr gazelle being slightly higher than normal (compared with the Indian blackbuck, Table 3). Springbok, nilgai, blesbok and nyala had liver Se levels below normal; the other species were within the normal range. Normal levels of dietary Se are between 0.1 mg/kg and 0.3 mg/kg DM (McDonald et al. 2002). Lucerne and teff fed to the NZG antelope contained no Se (Table 4), but antelope and Boskos cubes contained normal levels of Se. It is concluded that total dietary Se intake is inadequate, at least in some species. Perhaps this is caused by species differences in food preference or inter-specific competition; species with normal liver Se concentration may be selectively consuming the cubes, which have normal Se content. However, the sample size is too small to make valid conclusions. Since this study, nutritional and parenteral supplementation of NZG antelope with Se and other minerals has been introduced and morbidity and mortality as a result of myopathy has been reduced (data not shown).
Zinc
Zinc is distributed widely in body tissues but liver levels increase more rapidly after oral or intravenous Zn administration in normal or Zn deficient cattle than bone, red blood cells, hair and muscle levels (Pond et al. 2005). Plasma and serum values lack certainty and sensitivity as diagnostic criteria. Individual variability is high and Zn availability may be reduced by decreased Ca and increased P levels in the diet (McDonald et al. 2002). High dietary intake of Ca may be antagonistic to Zn utilisation and excess or deficiency of S reduces Zn availability.
Dietary Zn is required to mobilise and utilise Vitamin A stored in liver and to convert beta carotenes to Vitamin A in the intestinal wall. Liver Zn levels may increase with bacterial infection (Puls 1994). Zinc concentration in the liver and kidney increases at high levels of dietary Zn intake (Henry, Littell & Ammerman 1997). The single lechwe in the study had the highest Zn liver levels (159.00 mg/kg), followed by springbok liver samples (135.20 mg/kg ± 1.26 mg/kg). Both were considerably higher than recorded in the literature and within the toxic range for cattle and goats (Puls 1994). Therefore, these Zn levels may compromise Cu metabolism in these species. However, apart from interfering with Cu metabolism, Zn toxicity is rare (Haschek, Rousseaux & Wallig 2010). Individual liver Zn level variability is high within species and concentrations in tissues are very susceptible to stress and microbial infection (Underwood & Suttle 1999). The lechwe died of bacterial rumenitis, and two of the springbok died of bacterial enteritis and septicaemia, which may be relevant to mineral nutrition but are also related to feeding practices and hygiene.
According to Underwood and Suttle (1999), the determination of Zn in the diet can be helpful in diagnosing Zn deficiency. The Zn levels of lucerne and Boskos cubes were below the dietary recommended concentration of 45 mg/kg -100 mg/kg (Penrith et al. 1996; Puls 1994; Smith & Sherman 2009). The Zn concentration of antelope cubes was very high (14 700 mg/kg). Liver concentrations of Zn in all other animals in the study were within the normal range, suggesting that Zn nutrition is adequate for most species, most likely the result of the high Zn levels in the antelope cubes. Strangely, neither the springbok nor the lechwe had low liver Cu levels. For many of the elements, high to toxic intakes will not be reflected in a tissue until some threshold value is exceeded, whereupon it will increase dramatically (Henry et al. 1997).
Given the commercial importance of certain antelope in South Africa, too little is known about the mineral requirements of most free-ranging and captive antelope. The complexities associated with mineral nutrition and metabolism can confound the diagnosis of a mineral-related disease, because the manifestations of a mineral deficiency can differ amongst animals of different ages, sex and species, and can also be affected by the severity of the deficiency and the length of time over which the deficiency has occurred (Puls 1994; Underwood & Suttle 1999). Although clinical signs usually indicate the presence of a mineral imbalance, symptoms are rarely disease-specific (Underwood & Suttle 1999). For example, muscle necrosis that can be diagnosed readily at necropsy (if sufficient dystrophic mineralisation is present) and histologically may be caused by exertional myopathy, nutritional myopathy because of absolute or relative Se or vitamin E deficiency, ischaemic damage, intoxication with ionophores and various plant species, and hypokalaemia in ruminants (Van Vleet & Valentine 2007). Consequently, the diagnosis of a mineral-related disease is accomplished habitually through a mixture of clinical and biochemical analyses.
Mineral deficiencies may develop in captive animals as a result of inappropriate feeding, but also because of suboptimal environmental conditions. Free-ranging springbok prefer arid environments, dry grassy flats, Karoo scrub, salty pans, dune pathways, dry river beds and semi-desert shrub land with high mineral content. Their natural diet has high mineral content and they can utilise highly mineralised water (Skinner & Chimimba 2005). As small ruminants, they select food items with high nutritional value. Deficiencies may be supplemented by their habit of licking soil or rocks that are high in minerals (Van der Merwe & Perold 1967). This is not possible in most captive situations. Unfortunately, concentrations of minerals in the game block were not measured in this study. High levels of minerals are often added to commercially available lick blocks, which may cause toxicity if consumed in large amounts. Efforts to minimise the cost of mineral supplementation require knowledge of the supply and availability of mineral nutrients in feed and forages (Dost et al. 1990). A combination of factors makes it difficult to determine the actual mineral status of animals and their need and degree of supplementation. Detailed studies on mineral nutrition in healthy free-ranging or captive antelope are rarely published. In addition, disagreement exists on what concentrations indicate deficient, marginal or adequate intakes.
Some of the antelope in the present study died as the result of acute trauma, whilst others only died after a more chronic disease had taken its course. Diseased animals are likely to reduce their total food intake and thus alter their mineral intake and tissue mineral concentrations. Few studies have evaluated the effects of disease on the mineral status of either wild or domestic animals and, in most cases, it is difficult to determine whether the variation in mineral levels between healthy and diseased animals is a cause rather than an effect of the disease. In one study, differences between selected mineral concentrations in apparently healthy mule deer and those affected by chronic wasting disease were, in most cases, not significantly different (Wolfe et al. 2010).
Conclusion
This study has limitations. It was retrospective, using stored samples of which only limited numbers were available at the time of the study. Owing to the limited sample size, only tentative conclusions can be drawn regarding the Cu, Mn, Se and Zn nutritional status of the springbok in this study and therefore further investigation of stored samples from captive and free-ranging antelope would be necessary to confirm these findings. However, these preliminary results indicate possible significant mineral imbalances in both the antelope and their food items. Springbok liver concentrations of Cu and Mn were adequate, but Zn levels were higher than normal. Zinc may be interfering with Cu metabolism, although liver Cu levels were within normal range. In contrast, this study confirmed that Se nutrition in the springbok is inadequate, presumably because of the absence of Se in lucerne and teff. Although Se levels in the cubes were adequate, this apparently did not compensate for the absence of Se in the forage, because total dietary Se was deficient. Supplementation of Se is therefore indicated when antelope are fed a diet of lucerne and/or teff, even if they are fed cubes, particularly in multispecies enclosures, where smaller antelope may not have adequate access to more palatable feeds. This study has indicated that mineral malnutrition may be a feature in captive antelope in South Africa and highlighted the urgent need for further research on antelope nutrition. To this end, a detailed prospective study on mineral nutrition of antelope is planned at the NZG, using samples stored since 2007.
Acknowledgements
Excellent technical assistance was provided by the University of Pretoria Nutrilab. Financial assistance was obtained from the Royal Veterinary College (RVC) for Michael Lander, then a veterinary student at the RVC, London. The wBRC of the NZG of South Africa provided storage for the tissue samples.
Competing interests
The authors declare that they have no financial or personal relationship(s) which may have inappropriately influenced them in writing this paper.
Authors' contributions
K.R.M. (National Zoological Gardens of South Africa) wrote the manuscript, made conceptual contributions and conducted the statistical analysis. E.P.L. (National Zoological Gardens of South Africa) was responsible for project design, student supervision, collected the samples and assisted in writing the manuscript, whilst M.L. (Royal Veterinary College) prepared the samples, performed the experiments and assisted in the writing of the manuscript, A.S.W.T. (National Zoological Gardens of South Africa) assisted in the writing of the manuscript and made conceptual contributions and S.C. (University of Nottingham) was co-responsible for project design, student supervision and writing the manuscript.
References
Ange, K., Crissey, S.D., Doyle, C., Lance, K. & Hintz, H., 2001, 'A survey of African and Asian elephant diets and measured body dimensions compared to their estimated nutrient requirements', Proceedings of the Nutrition Advisory Group (NAG) 4, 5-14. [ Links ]
AOAC, 2000, Official methods of analysis, vol. 1, 17th edn., AOAC, Gaithersburg. [ Links ]
Brewer, N.R., 1987, 'Comparative metabolism of copper', Journal of the American Veterinary Medical Association 190, 654-656. PMid:3553117 [ Links ]
Burton, M. & Burton, R., 2002, International wildlife encyclopedia, 3rd edn., Marshall Cavendish, New York. [ Links ]
Clauss, M. & Dierenfeld, E.S., 1999, 'Susceptibility of yak (Bos grunniens) to copper deficiency', Veterinary Record 145, 436-437. http://dx.doi.org/10.1136/vr.145.15.436, PMid:10755592 [ Links ]
Clauss, M., Firzlaff, U., Castell, J.C., Kiefer, B., Streich, W.J. & Liesegang, A., 2007, 'Effect of captivity and mineral supplementation on body composition and mineral status of mustached bats (Pteronotus parnellii rubiginosus), Journal of Animal Physiology and Animal Nutrition 91, 187-192. http://dx.doi.org/10.1111/j.1439-0396.2007.00691.x, PMid:17516939 [ Links ]
Crawford, G.C., Puschner, B., Dierenfeld, E.S. & Dunker, F., 2009, 'Survey of minerals and fat-soluble vitamins in captive black and white ruffed lemurs (Varecia variegata)', Journal of Zoo and Wildlife Medicine 40, 632-638. http://dx.doi.org/10.1638/2008-0014.1, PMid:20063808 [ Links ]
Dierenfeld, E.S., Atkinson, S., Craig, A.M., Walker, K.C., Streich, W.J. & Clauss, M., 2005, 'Mineral concentrations in serum/plasma and liver tissue of captive and free-ranging rhinoceros species', Zoo Biology 24, 51-72. http://dx.doi.org/10.1002/zoo.20043 [ Links ]
Dost, M., Hussain, A., Khan, S. & Bhatti, M.B., 1990, 'Locational differences in forage yield and quality of maize cultivars', Pakistan Journal of Science and Industrial Research 33, 454-456. [ Links ]
Du, Z., Hemken, R.W. & Harmon, R.J., 1996, 'Copper metabolism of Holstein and Jersey cows and heifers fed diets high in cupric sulfate or copper proteinate', Journal of Dairy Science 79, 1873-1880. http://dx.doi.org/10.3168/jds.S0022-0302(96)76555-4 [ Links ]
Evans, G.W., 1973, 'Copper homeostasis in mammalian systems', Physiology Review 53, 353-570. [ Links ]
Haschek, W.M., Rousseaux, C.G. & Wallig, M.A., 2010, Fundamentals of toxicologic pathology, 2nd edn., Elsevier, London. [ Links ]
Hennig, A., Anke, M., Groppel, B. & Luedke, H., 1974, 'Secondary copper deficiency in ruminants', in W.G. Hoekstra, J.W. Suttie, H.E. Ganther & W. Mertz (eds.), Proceedings of International Symposium: Trace elements metabolism in animals-2, pp. 726-729, University Park Press, Baltimore. [ Links ]
Henry, P.R., Littell, R.C. & Ammerman, C.B., 1997, 'Bioavailability of cobalt sources for ruminants, 1: Effects of time and dietary cobalt concentration on tissue cobalt concentration', Nutrition Research 17, 947-955. http://dx.doi.org/10.1016/S0271-5317(97)00060-2 [ Links ]
Hoon, J.H., 2003, Mineral status of springbok (Antidorcas marsupialis) in different areas of South Africa, viewed 23 August 2010, from http://gadi.agric.za/articles/Hoon_JH/mineral-status-of-springbok.php [ Links ]
Johnson, H.E., Bleich, V.C. & Krausman, P.R., 2007, 'Mineral deficiencies in tule elk, Owens Valley, California', Journal of Wildlife Diseases 43, 61-74. PMid:17347394 [ Links ]
Long, C., 1961, Biochemists handbook, F.N. Spon Ltd, London. [ Links ]
McDonald, P., Edwards, R., Greenhalgh, J. & Morgan, C., 2002, Animal nutrition, 5th edn., Prentice Hall, Harlow. [ Links ]
McDowell, L., 1985, Nutrition of grazing ruminants in warm climates, Academic Press, New York. [ Links ]
McNaughton, S., 1990, 'Mineral nutrition and seasonal movements of African migratory ungulates', Nature 345, 613-615. http://dx.doi.org/10.1038/345613a0 [ Links ]
Miller, M., Amsel, S., Boehm, J. & Gonzales, B., 2001, 'Presumptive copper deficiency in hand-reared captive pronghorn (Antilocapra americana) fawns', Journal of Zoo and Wildlife Medicine 32, 373-378. PMid:12785688 [ Links ]
Penrith, M., Tustin, R., Thornton, D. & Burdett, P., 1996, 'Swayback in a blesbok (Damaliscus dorcas phillipsi) and a black wildebeest (Connochaetes gnou)', Journal of the South African Veterinary Association 67, 93-96. PMid:8765071 [ Links ]
Pond, W.G., Pond, K., Schoknecht, P.A. & Church, D.B., 2005, Basic animal nutrition and feeding, 5th edn., John Wiley & Sons Inc., New York. [ Links ]
Pugh, D.G., 2002, Sheep and goat medicine, W.B. Saunders Company, Philadelphia. [ Links ]
Puls, R., 1994, Mineral levels in animal health: Diagnostic data, 2nd edn., Sherpa International, Clearbrook. [ Links ]
Quan, M., Mulders, M.S.G. & Meltzer, D.G.A., 2002, 'The effect of storage conditions on samples for the evaluation of copper status in blesbok (Damaliscus pygargus phillipsi)', Journal of the South African Veterinary Association 73, 111-114. PMid:12515296 [ Links ]
Quan, M., 2005, 'Copper deficiency in blesbok (Damaliscus pygargus phillipsi) from the Karoo Nature Reserve', MSc thesis, Department of Veterinary Wildlife Unit, University of Pretoria. [ Links ]
Robbins, C.T., 1993, Wildlife feeding and nutrition, Academic Press, San Diego. [ Links ]
Skinner, J.D. & Chimimba, C.T., 2005, The mammals of the southern African subregion, Cambridge University Press, Cambridge. [ Links ]
Smith, M.C. & Sherman, D.M., 2009, Goat medicine, 2nd edn., Wiley Blackwell, New York. [ Links ]
Syakalima, M., Choongo, K., Nakazato, Y., Onauma, M., Sugimoto, C. & Tsubota, T., 2000, 'An investigation of heavy metal exposure and risks to wildlife in the Kafue Flats of Zambia', Journal of Veterinary Medical Science 63, 315-318. http://dx.doi.org/10.1292/jvms.63.315, PMid:11307933 [ Links ]
Underwood, E., 1977, Trace elements in human and animal nutrition, Academic Press, New York. [ Links ]
Underwood, E. & Suttle, N., 1999, The mineral nutrition of livestock, 3rd edn., CABI Publishing, New York. http://dx.doi.org/10.1079/9780851991283.0000 [ Links ]
Van der Merwe, F.J. & Perold, I.S., 1967, 'Trace elements in natural pastures', Journal of the South African Veterinary Association 38, 355-363. [ Links ]
Van Ryssen, J.B.J. & Hoon, J.H., 2010, Tissue mineral analysis of wildlife as indication of the mineral nutrition situation in regions, viewed 23 August 2010, from http://0-gadi.agric.za.innopac.up.ac.za/articles/Agric/tissuemineral.php [ Links ]
Van Vleet, J.F. & Valentine, B.A., 2007, 'Muscle and tendon', in M.G. Maxie (ed.), Jubb, Kennedy, and Palmer's pathology of domestic animals, vol. 1, pp. 185-280, Saunders Elsevier, Edinburgh. [ Links ]
Wieckowski, T., Abraham, A.A. & Wernery, U., 2008, 'Liver copper concentration in captive Thomson gazelles (Gazella thomsoni), in the united Arab Emirates', Proceedings of the European Association of Zoo and Wildlife Veterinarians 7, 287-292. [ Links ]
Webb, E., Van Ryssen, J., Erasmus, M. & McCrindle, C., 2001, 'Copper, manganese, cobalt and selenium concentrations in liver samples from African buffalo (Syncerus caffer) in the Kruger National Park', Journal of Environmental Monitoring 3, 583585. http://dx.doi.org/10.1039/b106307n, PMid:11785630 [ Links ]
Wolfe, L.L., Conner, M.M., Bedwell, C.L., Lukacs, P.M. & Miller, M.W., 2010, 'Select tissue mineral concentrations and chronic wasting disease status in mule deer from North-central Colorado', Journal of Wildlife Diseases 46, 1029-1034. PMid:20688718 [ Links ]
Zimmerman, T.J., Jenks, J.A., Leslie, D.M. & Neiger, R.D., 2008, 'Hepatic minerals of white-tailed deer and mule deer in the southern Black Hills, South Dakota', Journal of Wildlife Diseases 44, 341-350. PMid:18436666 [ Links ]
 Correspondence to:
Correspondence to:
Khanyisile Mbatha
Postal address: PO Box 754
Pretoria 0001
South Africa
Email: khanyi@nzg.ac.za
Received: 07 Sept. 2011
Accepted: 21 Apr. 2012
Published: 11 Sept. 2012


{kind=link}
{kind=link}