










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.82 n.4 Pretoria Dec. 2011
SHORT COMMUNICATION KORT MEDEDELING
A case study of rabies diagnosis from formalin-fixed brain material
J CoertseI; L H NelI; C T SabetaII; J WeyerI,III; A GroblerIV; J WaltersIV; W MarkotterI,*
IDepartment of Microbiology and Plant Pathology, Faculty of Natural and Agricultural Sciences, University of Pretoria, Pretoria, 0001 South Africa
IIAgricultural Research Council, Onderstepoort Veterinary Institute, Pretoria, 0001 South Africa
IIINational Institute of Communicable Diseases, National Health Laboratory Services, Sandringham, 2192 South Africa
IVGauteng Department of Agriculture, Conservation and Environment, Johannesburg, 2000 South Africa
ABSTRACT
Rabies is caused by several Lyssavirus species, a group of negative sense RNA viruses. Although rabies is preventable, it is often neglected particularly in developing countries in the face of many competing public and veterinary health priorities. Epidemiological information based on laboratory-based surveillance data is critical to adequately strategise control and prevention plans. In this regard the fluorescent antibody test for rabies virus antigen in brain tissues is still considered the basic requirement for laboratory confirmation of animal cases. Occasionally brain tissues from suspected rabid animals are still submitted in formalin, although this has been discouraged for a number of years. Immunohistochemical testing or a modified fluorescent antibody technique can be performed on such samples. However, this method is cumbersome and cannot distinguish between different Lyssavirus species. Owing to RNA degradation in formalin-fixed tissues, conventional RT-PCR methodologies have also been proven to be unreliable. This report is concerned with a rabies case in a domestic dog from an area in South Africa where rabies is not common. Typing of the virus involved was therefore important, but the only available sample was submitted as a formalin-fixed specimen. A real-time RT-PCR method was therefore applied and it was possible to confirm rabies and obtain phylogenetic information that indicated a close relationship between this virus and the canid rabies virus variants from another province (KwaZulu-Natal) in South Africa.
Keywords: epidemiology, formalin, Lyssavirus, rabies, real-time PCR, South Africa.
Rabies is a preventable disease but is nevertheless estimated to have an annual human death toll of at least 55 000 in canine rabies-endemic regions of the world - 40 % of these in Africa6,21.Itis caused by single-stranded, negative-sense RNA viruses of the Lyssavirus genus in the Rhabdoviridae family (order Mononegavirales). This genus, of which rabies virus (RABV) is the prototype, currently consists of 11 species all capable of causing rabies8. In the developing world, RABV infections contribute most significantly to the public and veterinary health burden of rabies. In southern Africa 2 variants of RABV circulate, namely the canid variant infecting members of the Canidae family and the mongoose variant infecting members of the Herpestidae13,19. Spillover infections of the mongoose variant in canid species are infrequently noted. In addition, 4 rabies-related lyssaviruses have been reported exclusively from the African continent, i.e. Lagos bat virus (LBV), Mokola virus (MOKV), Duvenhage virus (DUVV) and the more recently identified Shimoni bat virus (SHIBV)8,13. The primary reservoir for the rabies-related viruses (with the exception of MOKV) appears to be different bat species, from which spill-overs to various terrestrial species occur from time to time13. The reservoir species for MOKV is still unknown but this virus has been isolated from Crocidura spp.18 and a rodent17 and spill-over events in domestic (canine and feline)16 animals have also been reported. In South Africa, 602 animal rabies4 and 15 human rabies cases11 were laboratory confirmed during 2009. The majority of the animal cases were reported from 2 of the 9 provinces of South Africa, viz. KwaZulu-Natal (216 cases) and Mpumalanga (270 cases)4 (Fig. 1). No spill-over infections of rabies (mongoose/ canid variants) or of the rabies-related viruses were reported in 2009, although spill-over infections in dogs, cats, humans and mongooses have consistently been reported in South Africa and elsewhere in Africa in previous years9,10,16.

The fluorescent antibody test (FAT) is the gold standard for rabies diagnosis21,22 and yields the most reliable results when performed on fresh brain material. The test can also be modified for testing forma-lin-fixed material14,20. However, confirmatory and further characterisation tests such as virus isolation cannot be performed on formalin-fixed material due to inactivation of the virus and conventional reverse transcription polymerase chain reaction (RT-PCR) is problematic because viral RNA is known to degrade during formalin fixation1. Further characterisation of viral strains provides insights into the spread of the rabies virus variants and recognition of rabies-related virus cases in South Africa.
In August 2009 the owner of a 5½ year old German shepherd dog noticed abnormal behaviour in the animal. Apparently the dog had been confined to a property in the suburb of Linden, Johannesburg (Gauteng Province) for 4 years and had received rabies immunisation some 2-3 years prior to the observation. The owner did not recall any contact of this dog with wildlife or any other animals with the exception of a neighbour's pet rabbit. No other animal on the property or surrounding area presented with any unusual behaviour or characteristic rabies symptoms. The animal was euthanased by a local veterinarian and a Sellers stain revealed Negri bodies typical of rabies infection. Brain material in formalin was submitted to the Agricultural Research Council-Onderstepoort Veterinary Institute (ARC-OVI) for rabies diagnosis. Upon arrival at ARC-OVI the brain material was prepared for antigen detection by enzymatic digestion as described previously14. After digestion the sample was tested for the presence of Lyssavirus antigen with the FAT3 as described previously2,10, followed by examination under a fluorescent microscope (Axiovert 25, Zeiss). Brain tissues tested positive for Lyssavirus antigen with FAT (sample ID no. 864/09) (Onderstepoort Records 2009). In addition, total RNA was extracted by incubating brain material in 1 mℓ buffer (10 % SDS, 6 mg/m Proteinase K, 1 M Tris pH 7.5) at 37 ºC for 18 hours followed by RNA extraction using Trizol reagent (Invitrogen, USA) according to the manufacturer's instructions. Subsequently, RT-PCR and hemi-nested PCR (hnPCR) was performed according to previously published protocols2,10. However, RT-PCR and both hnRT-PCR assays were consistently negative despite repeated RNA extractions and increased amounts of RNA used for the cDNA synthesis steps.
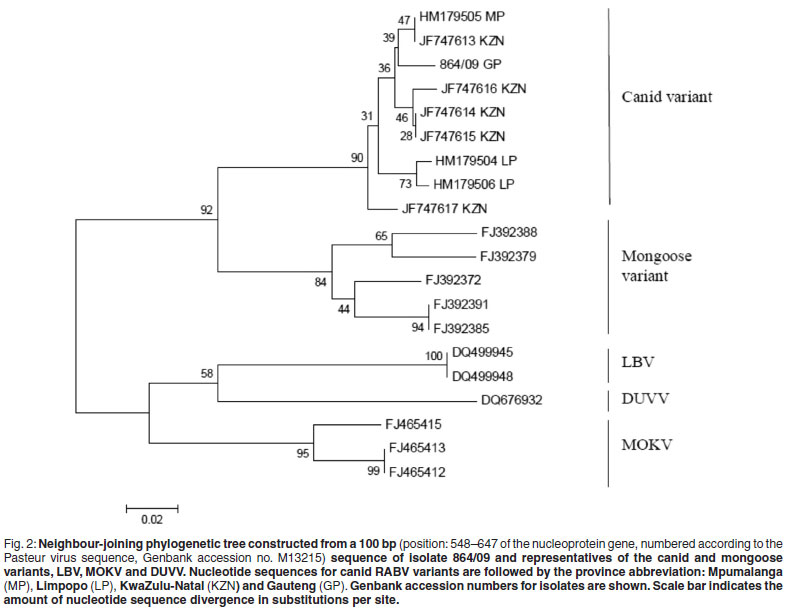
The immediate concern was with the aetiology of an unusual rabies case in the densely populated Gauteng Province of South Africa. Rabies in this province is relatively rare. Prior to 2005 there had been a long absence of cases and between 2005 and 2009 only sporadic cases were reported in dogs (7 cases), bovine (3 cases), mongooses (4 cases) and black-backed jackals (2 cases)4. All of these infections could be ascribed to classical RABV. Therefore, in addition to the usual tests, 1µl of extracted RNA was subjected to a quantitative real-time RT-PCR assay according to a previously described protocol2 using the LightCycler® RNA Amplification Kit HybProbe (Roche Diagnostics, Germany). Amplification was evident and 126 bp amplicons were purified and sequenced as described previously2,10. Sequences were trimmed using BioEdit Sequence Alignment Editor Version 7 and a consensus sequence constructed5. A multiple alignment of the consensus sequence of the specimen (reference no. 864/09) and representatives of other African lyssaviruses were constructed using the ClustalW subroutine of the BioEdit software5. This multiple alignment was used for the construction of a neighbour-joining (NJ) phylogenetic tree, using Molecular Evolutionary Genetics Analysis (MEGA) Version 3.1 software. Genetic distances were calculated using the Kimura 2-parameter method7. The reliability of the branching pattern was statistically evaluated by bootstrap analysis of 1000 replications.
Apart from the rarity of rabies in Gauteng Province to date, the apparent isolated nature of this case, the lack of possible sources for exposure to the dog and the vaccination history of the animal, raised concerns that the infection may have been due to a rabies-related virus. Phylogenetic analysis of a 100 bp sequence trimmed from the real-time RT-PCR amplicon sequence was performed with representative rabies and rabies-related Lyssavirus sequences available in our archive or in the public domain (Table 1). The resulting neighbour-joining (NJ) phylogenetic tree (Fig. 2) indicated that this isolate grouped with canine rabies virus variants from the Mpumalanga/ KwaZulu-Natal Provinces (97.9 % nucleotide identity and 97 % amino acid identity) indicating that the disease probably originated from one of these provinces. There is no efficient control of dog movement across provincial borders in South Africa or enforcement of vaccination of domestic animals when moved from a rabies-endemic area into areas where the disease has previously been under control. Rabies cases due to such movement have previously been reported12. In 2010, a rabies outbreak was identified in Gauteng province after the disease was laboratory confirmed in several domestic animals (with unknown exposure) as well as a human. Sequencing information of these cases also indicated the importation of the disease from the hyper-endemic KwaZulu-Natal Province15. In conclusion, the utility of a recently described real-time PCR protocol in the genetic typing of rabies virus RNA from a formalin fixed specimen was demonstrated where conventional typing methods were unsuccessful or not possible. This case is indicative of the continual spread of rabies into areas where the disease has been absent or under control for a number of years and highlights the importance of sustainable and continuous parenteral vaccinations.
ACKNOWLEDGEMENTS
This work was partially funded by the National Research Foundation (South Africa), Poliomyelitis Research Foundation, the National Health Laboratory Service Research Trust and the International Foundation for Science.
REFERENCES
1. Bustin S A 2002 Quantification of mRNA using real-time reverse transcription PCR (RT-PCR): trends and problems. Journal of Molecular Endocrinology 29:23-30 [ Links ]
2. Coertse J, Weyer J, Nel L H, Markotter W 2010 Improved PCR methods for detection of African rabies and rabies-related viruses. Journal of Clinical Microbiology 48:3949-3955 [ Links ]
3. Dean D J, Abelseth M K, Atansiu P 1996 The fluorescent antibody test. In Meslin F X, Kaplan M M, Koprowski H (eds) Laboratory techniques in rabies (4th edn). World Health Organization, Geneva: 88-95 [ Links ]
4. Department of Agriculture, Forestry and Fisheries. Chief Directorate Food & Veterinary Services, South Africa. Animal diseases in the RSA. Online at: http://www.nda.agric.za/vetweb/# (accessed 5 May 2011) [ Links ]
5. Hall T A 1999 BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT Nucleic Acids Symposium Series 41:95-98 [ Links ]
6. Knobel D L, Cleveland S, Coleman P G, Fèvre E M, Meltzer M I, MirandaMEG, Shaw A, Zinsstag J, Meslin F X 2005 Re-evaluating the burden of rabies in Africa and Asia. Bulletin of the World Health Organization 83:360-368 [ Links ]
7. Kumar S, Tamura K, Nei M 2004 Mega3: integrated software for molecular evolutionary genetic analysis and sequence alignment. Brief Bioinformatics 5:150-163 [ Links ]
8. Kuzmin I V, Mayer A E, Niezgoda M, Markotter W, Agwanda B, Breiman R F, Rupprecht C E 2010 Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Research 149:197-210 [ Links ]
9. Markotter W 2007 Molecular epidemiology and pathogenesis of Lagos bat virus, a rabies-related virus specific to Africa Ph.D. dissertation, University of Pretoria [ Links ]
10. Markotter W, Kuzmin I V, Rupprecht C E, Randles J, Sabeta C T, Wandeler A I, Nel L H 2006 Isolation of Lagos bat virus from water mongoose. Emerging Infectious Diseases 12:1913-1918 [ Links ]
11. National Institute for Communicable Diseases, Special Pathogens Unit. Annual report 2009. National Health Laboratory Service, Johannesburg [ Links ]
12. National Institute for Communicable Diseases 2009 Communicable diseases communiqué No. 8, August. National Institute for Communicable Diseases of the National Health Laboratory Service, Johannesburg: 5-6 [ Links ]
13. Nel L H, Markotter W 2007 Lyssaviruses. Critical Reviews in Microbiology 33:301-324 [ Links ]
14. Reid F L, Hall N H, Smith J S, Baer G M 1983 Increased immunofluorescent staining of rabies-infected, formalin-fixed brain tissue after pepsin and trypsin digestion. Journal of Clinical Microbiology 18:968-971 [ Links ]
15. Sabeta C, Phahladira B, Mohale D, Miyen J, Weyer J, Blumberg L H, Leman P A, Shumba W, Paweska J T 2011 Outbreak of rabies in the Gauteng Province, South Africa, 2010. Proceedings of the 10th International Meeting of the Southern and Eastern Africa Rabies Group, Maputo, Mozambique, 25-28 January 2011 [ Links ]
16. Sabeta C T, Markotter W, Mohale D K, Shumba W, Wandeler A I, Nel L H 2007 Mokola virus in domestic mammals, South Africa. Emerging Infectious Diseases 13:1371-1373 [ Links ]
17. Saluzzo J F, Rollin P E, Dauguet C, Digoutte J P, Georges A J, Sureau P 1984 Premier isolment du virus Mokola a partir d'un rongeur (Lophuromys sikapusi). Annals of Virology 135:57-66 [ Links ]
18. Shope R E, Murphy F A, Harrison A K, Causey O R, Kemp G E, Simpson D I H, Moore D L 1970 Two African viruses serologically and morphologically related to rabies virus. Journal of Virology 6:690-692 [ Links ]
19. Von Teichman B F, Thomson G R, Meredith C D, Nel L H 1995 Molecular epidemiology of rabies in South Africa: evidence for two distinct virus groups. Journal of General Virology 76:73-82 [ Links ]
20. Warner C, Fekadu M, Whitfield S, Shaddock J 1999 Use of anti-glycoprotein monoclonal antibodies to characterize rabies in formalin-fixed tissues. Journal of Virological Methods 77:69-74 [ Links ]
21. WHO 2005 Expert consultation on rabies, technical report series 931. World Health Organization, Geneva: 13:22-23 [ Links ]
22. World Organisation for Animal Health (OIE) 2006 Manual of diagnostic tests and vaccines for terrestrial animals (5th edn). World Organisation for Animal Health, Paris, France. Online at: http://www.oie. int/eng/normes/mmanual/a_00044.htm (accessed 5 May 2011) [ Links ]
Received: June 2011
Accepted: September 2011
* Author for correspondence. E-mail: wanda.markotter@up.ac.za


{kind=link}
{kind=link}