










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.81 n.4 Pretoria Dec. 2010
ARTICLE ARTIKEL
Macroscopic description of the coronary arteries in Swiss albino mice (Mus musculus)
A YoldasI,*; E OzmenII; V OzdemirIII
IAdana Veterinary Control and Research Institute, 01122 Adana, Turkey
IIDepartment of Anatomy, Faculty of Veterinary Medicine, University of Mustafa Kemal, 31040 Hatay, Turkey
IIIDepartment of Anatomy, Faculty of Veterinary Medicine, University of Afyon Kocatepe, 03100 Afyon, Turkey
ABSTRACT
A total of 25 (13 male, 12 female) adult, healthy Swiss albino mice were used to investigate the origin, course and anastomoses of coronary arteries. Coloured latex was injected into the aortic arch to enable these arteries to be clearly discerned. A. coronaria sinista was larger than A. coronaria dextra. It was divided into a Ramus interventricularis paraconalis and a Ramus circumflexus sinister. However, in 2 specimens, the septal ramus, was observed to stem directly from the left coronary artery, and only 1 ventricular branch arose from the left circumflex. The collateral branches of the paraconal interventricular ramus had a larger diameter and more extensive distribution was observed in these specimens. The A. coronaria dextra was divided into a Ramus septalis and Ramus circumflexus dexter. The Ramus interventricularis subsinuosis was not detected in this study. The ventricular branches of the left coronary artery run intramyocardially whereas the branches of the right coronary artery course subendocardially.
Keywords: coronary artery, heart, mice, subgross anatomy.
INTRODUCTION
The course and distribution of the coronary arteries of laboratory animals1, 7, 9, 10, 25, 32, domestic mammals25, 34, and birds20 are well known. The coronary arteries of the dormouse29, ordinary mice17 and rodents that have anomalous hearts19 have also been investigated. However, in these studies the authors investigated abnormalites of the origin of the main coronary arteries and used different methods. In this study, we aimed to reveal the conformation and branching of the coronary arteries in Swiss albino mice subgrossly.
The mouse is a model that has been widely used to study several aspects of the cardiovascular system: embryology, physiology18, the molecular determinants of coronary arteries6, 33, the control of arteriosclerosis18, 31, and the mechanism of ischaemia-reperfusion23, 30. This opportunity prompted us to study the anatomy of the coronary arteries in the hope that our findings would be useful for clinical studies with similar anatomical settings.
MATERIALS AND METHODS
Twenty-five adult healthy Swiss albino mice (Mus musculus) (13 males, 12 females), 10-12 weeks old weighing 35-40 g, maintained at the animal house of the department of Adana Veterinary Control and Research Institute, Adana, were used for the present study. The colony was maintained under controlled conditions of temperature and light (Light: dark, 12 h: 12 h.). The animals were provided standard mice feed (procured from Tavas Ltd., Turkey) and water ad libitum.
They were first anaesthetised with combination a of 10 mg/kg xylazine (Rompun®; Bayer Turk Kimya San. Ltd. Sti. Istanbul) and 100 mg/kg ketamine HCl (Ketalar® Eczacbasi Istanbul) intraperitonaly. Heparin (Liquemine IV, Roche Mustahzarlari San. A.S.) was administered (450 U/20/g, IV) to prevent coagulation. Secondly, the animals were euthanased by means of an incision in the aorta ascendens, while they were in deep anesthesia. The vessels were washed with 0.9 % physiological saline, and red coloured latex was injected in to the aortic arch. After polymerisation at room temperature, the hearts were studied using a dissection microscope (Nikon SMZ-2T, Nikon Corp., Tokyo, Japan). The findings were recorded and photographed (Sony DSC F 717, Sony Corp., Tokyo, Japan).
All procedures were approved by the Adana Veterinary Control and Research Institute Experimentation Ethics Committee.
Anatomical nomenclature of the coronary arteries was in accordance with Nomina Anatomica Veterinaria (2005)22 and previous reports5, 14-16, 21.
RESULTS
The vascularisation of the heart was provided by the branches of the left and right coronary arteries and it was observed that the left coronary artery was dominant.
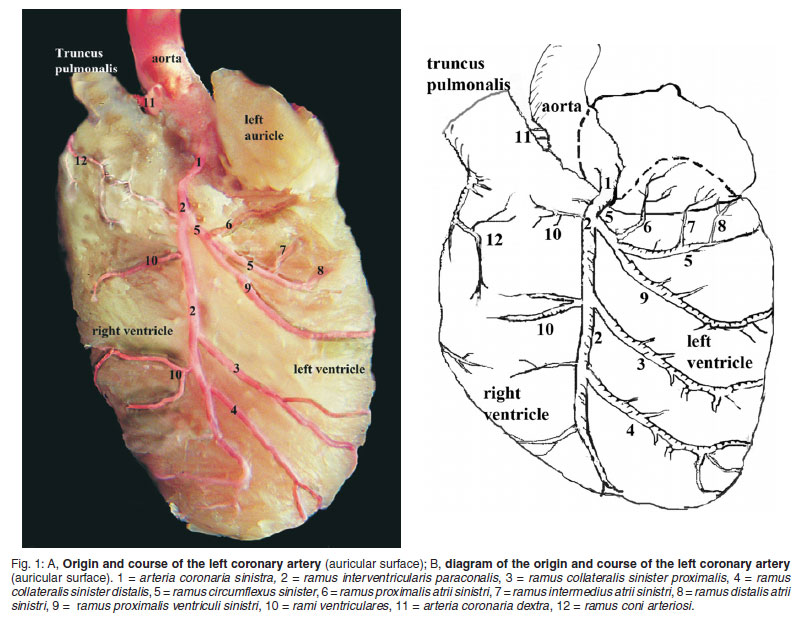
The left coronary artery arose from the sinus of the aorta, coursed left and distally between the pulmonary trunk and left auricle just after its origin and gave rise to 1-2 branches to the left atrium. Passing underneath the left auricle, the left coronary artery was observed to course in the sulcus interventricularis paraconalis, which was not evident (Figs 1A, B/1, 3A, B/1, 4A, B/3).
The ramus interventricularis paraconalis was the continuation of the left coronary artery which coursed in the paraconal interventricular groove. The terminal branches of this artery proceeded to the apex cordis and arrived at the atrial surface of the heart. Generally, the branches of this vessel course within the myocardium but in mice it was determined that this vessel ran subendocardially for the first 1/3 of its course (Fig. 1A, B/2, 3A, B/2).
This study indicated that the paraconal interventricular branch was the strongest artery to supply the heart.
The ramus interventricularis paraconalis gave off the branches of the so called ramus coni arteriosi, ramus collateralis sinister proximalis, ramus collateralis sinister distalis and rami septales.
The ramus coni arteriosi arose from the paraconal interventricular ramus and coursed subepicardially in the direction of the pulmonary trunk. This vessel was divided into terminal branches at the region of the conus arteriosus and in 10 specimens, anastomosed with same branches of the right coronary artery (Fig. 1A, B/12).
The ramus collateralis sinister proximalis originated from the paraconal interventricular ramus and after its origin it coursed on the wall of the left ventricle to arrive caudal to the border of the interventricular septum. In its course this vessel gave off branches which supply the wall of the left ventricle and caudal region of the interventricular septum. The terminal branches of this vessel were observed to reach the right ventricle where they anastomosed with the septal branches of the right coronary artery. This vessel and its branches were seen intramyocardially for their entire course (Fig. 1A, B/3).
The ramus collateralis sinister distalis stemmed from the distal half of the paraconal interventricular ramus. It ran parallel to the ramus collateralis sinister proximalis on the left ventricle and almost reached the apex of the heart. Along its course, the vessel gave off branches to supply the distal third of the left ventricle and caudal region of the interventricular septum. In addition, the terminal branches were seen to supply the apex of the heart (Fig. 1A, B/4).
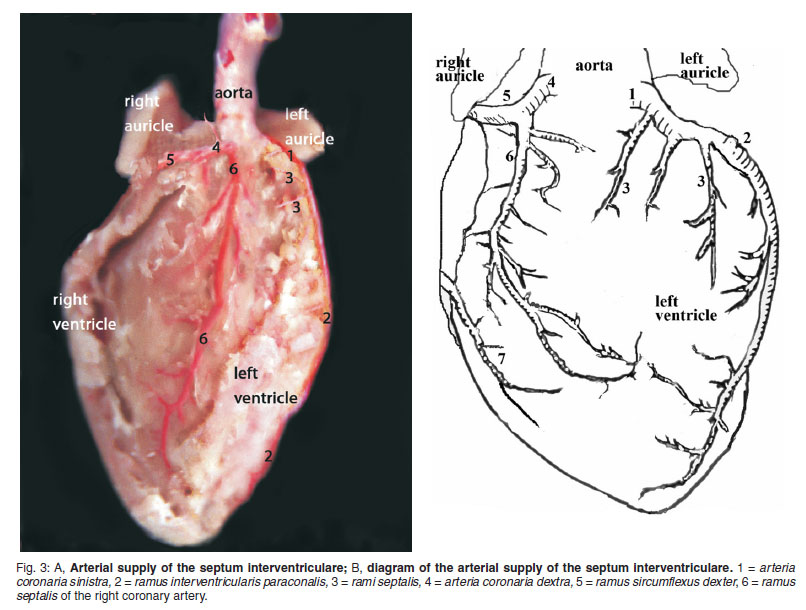
The rami septales were observed to arise from the medial margin throughout the paraconal interventricular ramus. These septal branches ran distally after their origin and their diameters were small. Terminal branches of these vessels were dispersed to the cranial region of the interventricular septum and mm. papillaris, supplying these areas (Fig. 3A, B/3).
The ramus circumflexus sinister arose from the left coronary artery at the level of the left auricle and coursed intramyocardially, below the left auricle, towards the margo ventricularis sinister (Fig. 1A, B/5).
The ramus proximalis atrii sinistri usually arose just after the origin of the left circumflex ramus generally (21 of 25), but in 4 mice it stemmed directly from the dorsal wall of the left coronary artery. After its origin, it coursed proximally and reached the free margin of the left auricle. In this area the vessel was observed to divide into terminal branches and in addition to these branches, it was determined that the ramus proximalis atrii sinistri gave off some branches to supply the medial surface of the right auricle and left ventricle (Fig. 1A, B/6).
The ramus intermedius atrii sinistri was seen to branch from the left circumflex ramus, run towards the left auricle and disperse to the medial and lateral surface of the left auricle. Along its course, on the wall of the left ventricle, a few branches started from the ramus intermedius atrii sinistri at the level of the left atrioventricular ostium to supply the left ventricle (Fig. 1A, B/7).
The ramus distalis atrii sinistri stemmed from the cranial edge of the left circumflex ramus on the left ventricle. This vessel and its terminal branches were dispersed to the medial region of the left atrium and auricle (Fig. 1A, B/8).
The ramus proximalis ventriculi sinistri was the strongest branch to arise from the left circumflex ramus and supply the left ventricle. After its origin, the vessel ran obliquely on the wall of the left ventricle and then coursed caudally towards the sulcus interventricularis subsinuosus. The terminal branches of this vessel were observed to supply the mm. papillares in the left ventricle (Fig. 1A, B/9).
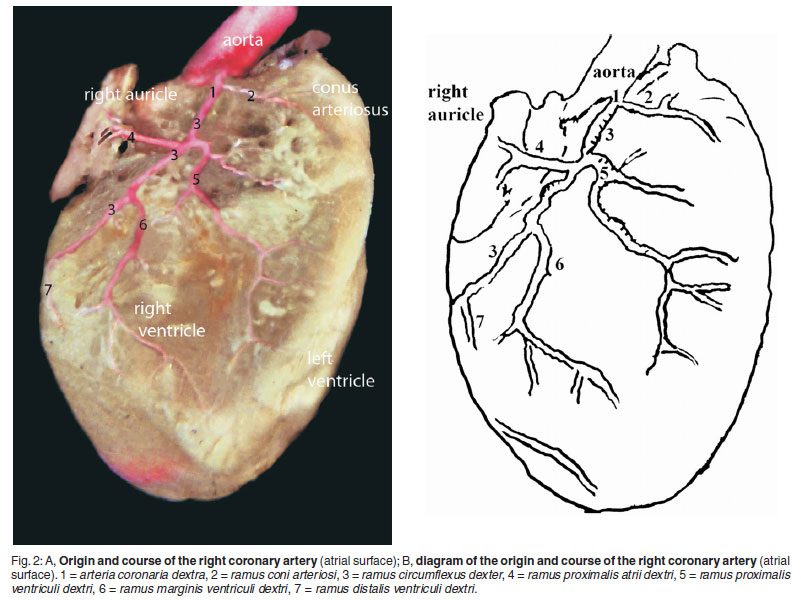
The arteria coronaria dextra arose from the aorta at the level of the sinus aorta, but in 2 cadavers it originated with the left coronary artery from a common root, which stemmed from the aorta. Just after its origin, the right coronary artery gave off the ramus septalis and then coursed caudally towards the right ventricle, between the pulmonary trunk and right auricle (Fig. 2A, B/1, 3A, B/4, 4A, B/1).
The ramus circumflexus dexter was the continuation of the right coronary artery and coursed onto the atrioventricular border. It terminated close to the sulcus interventricularis subsinuosus and gave off branches which anastomosed with terminal branches of the ramus proximalis ventriculi sinistri (Fig. 2A, B/3, 3A, B/5).
The ramus proximalis atrii dextri was seen to branch from the right coronary artery just opposite to the ramus proximalis ventriculi dexter. It was observed that this branch was the strongest vessel supplying the right auricle. After its origin, this branch reached the free edge of the right auricle and divided into 2 branches, of which the 1st dispersed on the free edge of the right auricle and the 2nd dispersed on the medial and lateral wall of the right auricle (Fig. 2A, B/4).
The ramus coni arteriosi started from the right coronary artery as 1 root in 22 mice. However, in 3 mice this branch sprang from the right coronary artery as 2 branches, close to one another. It coursed subendocardially towards the conus arteriosus and ended in this area after giving off terminal branches (Fig. 2A, B/2).
The ramus proximalis ventriculi dextri was the 1st branch of the right coronary artery to the right ventricle, which ran intramyocardially. While it coursed distally, it gave off 2-3 branches in the directions of the sulcus interventricularis paraconalis. After that, the ramus proximalis ventriculi dextri was observed to course towards the apex cordis and gave off terminal branches which ended at the distal region of the left ventricle (Fig.2A, B/5).
The ramus marginis ventriculi dextri arose the from the right circumflex ramus just after the point of origin of the ramus proximalis ventriculi dextri. It was the strongest branch that started from the right circumflex ramus to supply the right ventricle. After its origin, this vessel ran obliquely on the ventricular wall and divided into terminal branches which vascularised the apex cordis. Before giving off the terminal branches this vessel coursed intramyocardially, but the terminal branches ran subepicardially in the area of the apex. (Fig.2A, B/6).
The ramus distalis ventriculi dextri originated from the right circumflex artery after passing the margo ventricularis dexter. This thin branch was observed to course obliquely on the wall of the right ventricle. It was also recorded that this branch arose from the ramus marginis ventriculi dexter in 2 mice (Fig. 2.A, B/7).
The ramus septalis was seen to branch from close to the point of origin of the right coronary artery in 20 mice (Fig. 3A, B/6), but in 2 mice it arose from the left coronary artery and in 3 mice it originated directly from aorta (Fig. 4A, B/2). Just after its origin, the vessel was seen to enter the interventricular septum at the level of the right semilunar valve. While it coursed caudoventrally, the ramus septalis gave off a few branches. Some of these branches reached the mm. papillares and the others, which coursed caudally, ended at the wall of the left ventricle and had anastomoses with ventricular branches of the left coronary artery.
Originating from the aorta, the left and right coronary arteries ran subepicardialy before entering the myocardium. Primary and secondary branches of the left coronary artery ran intramyocardially on the wall of the left ventricle but the secondary branches of the right coronary artery coursed subendocardially. However, the atrial branches coursed subepicardially.
DISCUSSION
As reported by other authors3, 4, 7, 9, 21, 27, 28, it was observed in Swiss albino mice that the left and right coronary arteries originated from the sinus aorta and were distributed through the myocardium. However, it has been reported that the right coronary artery is normally absent in the chinchilla26.
In contrast to beavers7, the left coronary artery was stronger than right coronary artery in Swiss albino mice, as reported in dogs11,34, rabbits4, 8, 27, Syrian hamsters28, porcupines3 and dormice29.
As also reported in ruminants16, donkey25, rabbit5, 13, beaver7, porcupine3, cat and dog15, in the present study in 23 Swiss albino mice it was observed that the left coronary artery was divided into 2 branches, the ramus interventricularis paraconalis and the ramus circumflexus sinister. In addition to these, it gave off the ramus septalis in 2 animals. However, some authors stated that it was divided into a ramus marginis ventricularis sinistri, a ramus posterior ventriculi sinistri and a ramus septalis in New Zealand rabbits2 or into a ramus marginis ventricularis sinistri, a ramus posterior ventriculi sinistri and a ramus atrialis sinistri in Angora rabbits24 or into a ramus interventricularis paraconalis, a ramus marginis ventricularis sinistri and a ramus circumflexus sinister in rabbits8.
The atrial branches vascularising the left atrium have been documented to originate from the left posterior ventricular ramus in the New Zealand Rabbit2, or from the left coronary artery in the Angora rabbit4, 8 and dormouse29. However, this study recorded the origin of these branches to be the left circumflex ramus in Swiss albino mice, as reported in the chinchilla26, rabbit8, beaver7 and porcupine3.
It was observed that the paraconal interventricular ramus was stronger than the left circumflex ramus in the Swiss albino mice in this study, in contrast to reports about the dog11, Angora rabbit4 and donkey25.
Although some authours stated that the paraconal interventricular ramus did not reach the apex of the heart in ruminants16, donkey25, rabbit5, 13, New Zealand rabbits2, beaver7, cat and dog14, 15, it was observed in Swiss albino mice that this vessel reached the apex cordis and coursed through to the atrial surface of the heart.
Although it was reported that the left circumflex ramus gave off 3 branches, the ramus proximalis ventriculi sinistri, the ramus marginis ventriculi sinistri and the ramus distalis ventriculi sinistri, to supply the wall of the left ventricle in ruminants21, dog11 and Angora rabbit4, inthe present study the ramus marginis ventriculi sinistri and the ramus distalis ventriculi sinistri were not seen and this region was supplied by the ramus proximalis ventriculi sinistri in Swiss albino mice.
In this study it was observed that the left ramus coni arteriosi arose from the paraconal interventricular ramus as reported in rabbit8, 6 of 8 Angora rabbits4, New Zealand rabbit13 and mice17. However, it originated from the ramus marginis ventricularis sinistri in the Angora rabbit24 and New Zealand rabbit2 and from right coronary artery in the porcupine3.
Arterial supply of the septum interventriculare is supplied by the ramus septi interventricularis and rami septales originating from the ramus interventricularis paraconalis and by the rami septales branch from the ramus interventricularis subsinuosus in New Zealand rabbit13, beaver7, cat15, dog14, 15, sheep16, goat16, ox16, pig21, horse21, and donkey25. However, it is pointed out that, in the Swiss albino mice rami septales arose from the ramus interventricularis paraconalis or right coronary artery to supply the septum.
Although several authors stated that the ramus interventricularis subsinuosus originated from the ramus circumflexus sinister in ruminants16, 21 or from a. coronaria sinistra in the horse21, pig21 and donkey25, some authors pointed out that this vessel originated from the ramus circumflexus sinister or ramus circumflexus dexter, or from both as a bilateral coronary type in the cat and dog2, 15, 21 and that exceptionally it may not exist in the cat15, 21. In this study the ramus interventricularis subsinuosus did not exist in the Swiss albino mice, as reported in Angora rabbit4, in other rabbits2, 8, 13, 27 and in rats1.
As reported in the dog12, cardiac muscle bands were seen that crossed the coronary arteries obliquely, especially when they were running in the sulci, but when they leave the sulci, the arteries course parallel to muscle fibres of the myocardium.
In conclusion, this study revealed that the ramus septalis, a branch from the a. coronaria dextra, is the main vessel responsible for supplying blood to the interventricular septum and the septal branches of the paraconal interventricular ramus are also involved. Additionally, the ramus interventricularis subsinuosis is absent in Swiss albino mice. It is hoped that the results of the present study will encourage further research in this field.
REFERENCES
1. Ahmed S H, Rakhawy M T, Abdaila A, Assaad I 1978 The comparative anatomy of the blood supply of cardiac ventricles in the albino rat and guinea pig. Journal of Anatomy 26: 51-57 [ Links ]
2. Aksoy G, Karadag H 2002 An anatomic investigation on the heart and coronary arteries in the domestic cat and white New Zealand rabbits. Veteriner Bilimleri Dergisi 18: 33-40 [ Links ]
3. Atalar A, Yilmaz S, Ilkay E, Burma O 2003 Investigation of coronary arteries in the porcupine (Hystrix cristata) by latex injection and angiography. Annals of Anatomy 185: 373-376 [ Links ]
4. Bahar S, Ozdemir V, Eken F, Tipirdamaz S 2007 The distribution of the coronary arteries in the Angora rabbit. Anatomia Histologia Embryologia 36: 321-327 [ Links ]
5. Barone R, Pavaux C, Blin P C, Cuo P 1973 Atlas d'anatomie du lapin. Masson & Cie, Paris [ Links ]
6. Bellomo D, Headrick J P, Silins G U, Paterson C A, Thomas P S, Gartside M, Little M, Cummings M C, Hayward N K, Kay G F 2000 Mice lacking the vascular endothelial growth factor-b gene (vegfb) have smaller hearts, dysfunctional coronary vasculature, and impaired re-covery from cardiac ischemia. Circulation Research 86: 29-35 [ Links ]
7. Bisaillon A 1981 Gross anatomy of the cardiac blood vessels in the North American beaver (Castor canadensis). Anatomischer Anzeiger 150: 248-258 [ Links ]
8. Day S B, Johnson J A 1958 The distribution of the coronary arteries of the rabbit. Anatomical Record 132: 633-643 [ Links ]
9. Dowd D 1991 The coronary vessels in the heart of a marsupial (Trichosorus vulpecula). American Journal of Anatomy 140: 47-56 [ Links ]
10. Duran A C, Sans-Coma V, Arque J M, Cardo M, Fernandez B, Franco D 1992 Blood supply to the interventricular septum of the heart in rodents with intramyocardial coronary arteries. Acta Zoologica 73: 223-229 [ Links ]
11. Dursun N 1979 Kope in Kalp Arteria'lari Uzerinde Atomik Arastirmalar. Ankara Universitesi Veteriner Fakultesi Dergisi 26: 16-33. [ Links ]
12. Dursun N, Tipirdamaz S, Erden H, Celik I 1992 Evcil Memeli Hayvanlarda Kalp Kas Kopruleri Uzerinde Makroskopik ve Mikroskopik Arastirmalar. Selcuk Universitesi Veteriner Fakultesi Dergisi 8: 12-17. [ Links ]
13. Dursun N, Yildiz D, Kabak M, 1996 Yeni Zeland Tavsaninda (Oryctolagus cuniiiculus L.) Septum Intervenriiculare'nin Arteriel Vaskularizasyonu. Ankara Universitesi Veteriner Fakultesi Dergisi 43: 391-395 [ Links ]
14. Evans H E, Christensen G C 1979 Miller's anatomy of the dog (2nd edn). W B Saunders, Philadelphia [ Links ]
15. Habermehl K H 1994 Herz. In Anatomie von Hund und Katze. Blackwell Wissenschafts-Verlag Berlin: 251-255 [ Links ]
16. Hegazi A 1958 Die Bultgefass Beversorgung Des Herzens Von Rind, Schaf und Ziege. DVSc thesis, Justus Liebig Universitat, Gieben [ Links ]
17. Icardo J M, Colvee E, 2001 Origin and course of the coronary arteries in normal mice and in Iv/Iv Mice. Journal of Anatomy 19: 473-482 [ Links ]
18. Isobe M, Suzuki J, Morishita R, Kaneda Y, Amano J 2000 Gene therapy for heart transplantation associated coronary arteriosclerosis. Annals of the New York Academy of Sciences 90: 77-83 [ Links ]
19. Jose M, Arque M D, Victoria C, Luis M, Sans-Coma R, Sans-Coma V 1986 Congenital anomalies of coronary arteries in rodents. American Journal of Cardiology 56: 498-499 [ Links ]
20. Linsday F E F, Smith H J 1965 Coronary arteries of Gallus domesticus. American Journal of Anatomy 116: 301-314 [ Links ]
21. Nickel R A, Shummer A, Seiferle E 1981 The anatomy of the domestic animals. Volume 3: The circulatory system. Verlag Paul Parey, Berlin-Hamburg [ Links ]
22. Nomina Anatomica Veterinaria, 2005 International Committee on Veterinary Gross Anatomical Nomenclature, World Association of Veterinary Anatomists, New York [ Links ]
23. Nossuli T O, Lakshminarayanan V, Baumgarten G, Taffet G E, Ballantyne C M, Michael L H, Entman M L 2000 A Chronic mouse model of myocardial ischemiareperfusion: essential in cytokin studies. American Journal of Physiology 278: 1049- 1055 [ Links ]
24. Nur I H, Duzler A 2004 Ankara Tavsaninda Kalp ve Koroner Arterler Uzerinde Makroanatomik bir Arastirma. Ulusal Veteriner Anatomi Kongresi Kusadasi, Turkiye, 16-19 Eylul 2004: 5 [ Links ]
25. Ozgel O, Haligur A, Dursun N, Karakurum E 2004 The macroanatomy of coronary arteries in donkeys (Equus asinus L.). Anatomia Histologia Embryologia 33: 278-283 [ Links ]
26. Ozdemir V, Cevik-Demirkan I, Turkmenoglu 2008 The right coronary artery is absent in the chinchilla (Chinchilla lanigera). Anatomia Histologia Embryologia 37: 114- 117 [ Links ]
27. Podesser B, Wollenek G, Seitelberger R, Siegel H, Wolner E, Firbas W, Tschabitscher M 1997 Epicardial branches of the coronary arteries and their distribution in the rabbit heart: The rabbit heart as a model of regional ischemia. Anatomical Record 247: 521-527 [ Links ]
28. Sans-Coma V, Arque J M, Duran A C, Cardo M, Fernandez B, Franco D1993 The coronary arteries of the Syrian hamster Mesocricetus auratus (Waterhouse 1839). Annals of Anatomy 17: 53-57 [ Links ]
29. Sans-Coma V C, Duran M, Cardo M, Arque J M 1995 The coronary arteries of the garden dormouse. Hystriw 6: 217-224 [ Links ]
30. Scherrer-Crosbie M, Steudel W, Ulrich R. Hunziker P R, Liel-Cohen N, Newell J, Zaroff J, Zapol W M, Picard M H 1999 Echocardiographic determination of risk area size in a murine model of myocardial ischemia. American Journal of Physiology 277: 986-992 [ Links ]
31. Suzuki J, Isobe M, Morishita R, Nishikawa T, Amano J, Kaneda Y 2000 Antisense Bcl-x oligonucleotide induces apoptosis and prevents arterial neointimal formation in murine cardiac allografts. Cardiovascular Research 45: 783-787 [ Links ]
32. Teofilovski P G, Kreclovi G 1998 Coronary artery distribution in Macaca fascicularis. Laboratory Animals 3: 200-205 [ Links ]
33. Tevosian S G, Deconinck A E, Tanaka M, Schinke M, Litovsky S H, Izumo S, Fujiwara Y, Orkin S H 2000 Fog-2, a cofactor for GATA transcription factors, is essential for heart morphogenesis and development of coronary vessels from epicardium. Cell 101: 729-739 [ Links ]
34. Tipirdamaz S, Dursun N, Yalcin H 1996 Kangal Kopeklerinde Kalbin Koroner Arterleri Uzerinde Makroanatomik Calismalar. Veteriner Bilimleri Dergisi 12: 115-120. [ Links ]
Received: November 2009.
Accepted: November 2010.
* Author for correspondence. E-mail: atillayoldas99@hotmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}