










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkJournal of the South African Veterinary Association
versión On-line ISSN 2224-9435
versión impresa ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.81 no.4 Pretoria dic. 2010
ARTICLE ARTIKEL
Tick communities at the expanding wildlife/cattle interface in the Eastern Cape Province, South Africa: implications for Corridor disease
E R Smith*; D M Parker
The Wildlife and Reserve Management Group, Department of Zoology and Entomology, Rhodes University, Grahamstown, 6140 South Africa
ABSTRACT
Corridor disease, transmitted by the brown ear tick (Rhipicephalus appendiculatus), is one of Africa's most pathogenic tick-borne diseases for cattle. With a focus on this species, we investigated the community parameters (richness, diversity and abundance) of ticks in the Eastern Cape, South Africa, and how this may be linked to the increasing wildlife/cattle interface in the region. There were significantly more ticks of a greater diversity and richness at sites positioned at the wildlife/cattle interface ('treatment sites') compared to sites where wildlife was absent (controls). Significantly, R. appendiculatus was only found at the treatment sites. Therefore, it is believed that the wildlife/cattle interface may be playing a crucial role in increasing the occurrence, abundance and distribution of R. appendiculatus in the Eastern Cape. The implications of a Corridor disease outbreak in the region are discussed.
Keywords: buffalo, disease-free, Theileria, ticks.
INTRODUCTION
Ticks and their associated diseases are a major constraint to livestock farming throughout the world but particularly in Africa where socioeconomic factors govern the extent to which livestock farmers can control ticks15, 17. Annually, in Africa, tick-borne diseases kill 1.1 million cattle resulting in economic losses of $160 million19. Most African wildlife have innate resistance to such diseases but may act as reservoirs for a number of tick-borne diseases such as Corridor disease12. The Cape buffalo (Syncerus caffer) is the natural host for the protozoan parasite Theileria parva, which causes Corridor disease and is transmitted to cattle via the vector Rhipicephalus appendiculatus (the brown ear tick)23. A change in land use from agricultural based farming to game farming has led to a growing cattle/wildlife interface in the Eastern Cape, South Africa21. Therefore it has become vital to establish the current status of the vector, R. appendiculatus, along these interfaces. The detection of its presence or extension of its previous range may represent a disease 'time-bomb' and could see the introduction of Corridor disease into a region where many rural livelihoods are based on livestock farming. In addition, the buffalo in the Eastern Cape have long been considered 'disease-free' on the basis of serological evidence and no clinical disease having been reported in the cattle associated with these buffalo24. Buffalo are classified as 'disease-free' when they test negative for foot-and-mouth disease, bovine brucellosis, bovine tuberculosis and Corridor disease26.
The brown ear tick is known to occur in the Eastern Cape but mainly in the areas closer to the coast10, 19 (Fig. 1). Currently, the farthest inland specimens are known from Bucklands Farm (33º06'S, 26º41'E)2. However, populations of R. appendiculatus are often patchy because they are limited by factors such as climate, vegetation and the presense of suitable hosts29. Adults show habitat preference for bush, scrub and long grass when vegetation coverage is adequate25, 29. Theileria parva is acquired by immature ticks during blood feeding and the parasite itself is only transmitted by the nymphal and adult tick stages.
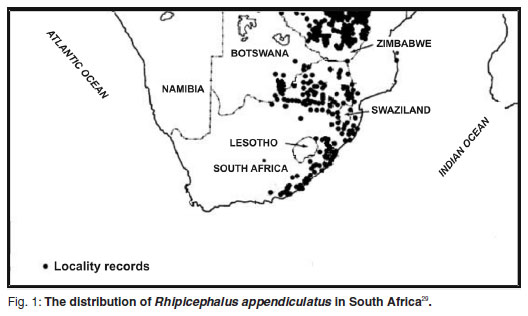
A species distribution and regional climate model has been used to assess the impact of climate change on the tick-borne disease theileriosis in sub-Saharan Africa19. This study concluded that with climate change the Northern and Eastern Cape Provinces of South Africa will rapidly increase in suitability for hosting R. appendiculatus and thus, the potential for outbreaks of Corridor disease19. Therefore, it is vital to assess the current distribution of R. appendiculatus in these areas.
The overall aims of this study were: 1) to provide baseline data on the species diversity, abundance and richness of tick populations in areas at wildlife/cattle interfaces compared with areas without such an interface, and; 2) to determine the current status of the vector for Corridor disease, R. appendiculatus, and whether there is a relationship between its distribution and the increasing wildlife/cattle interface in the Eastern Cape.
MATERIALS AND METHODS
The study took place at Kwandwe Private Game Reserve (33º09'S, 26º37'E), Shamwari Private Game Reserve (33º28'S, 26º03'E) (referred to as the treatments). Both the treatment sites fall broadly into the Subtropical Thicket habitat9. Kwandwe Private Game Reserve consists of Bushclump Savanna and Medium Portulacaria Thicket20. Shamwari Private Game Reserve has a more mosaic vegetation distribution. In the reserve 44.2 % of the habitat is Subtropical Thicket and the remaining portions of the reserve are made up of Afromontane Forest, Subtropical Thicket, Bontveld, Bushclump Savanna, Grassy Fynbos, Karoo Scrub, Riverine Bush, Montane Grassland, Open Grassland, Primary Acacia Thicket, Secondary Acacia Thicket, cleared and cultivated land9, 20. Strowan Farm (33º18'S, 26º28'E) and Nanaga Farm (33º36'S, 25º55'E) were the control sites for this research. Nanaga farm consists of Kikuyu-dominated pastures (M. Mackenzie, landowner, pers. comm., 2009) while Strowan Farm consists of grazing pastures dominated by Themeda triandra and thicket clumps dominated by Rhus refracta (M. Palmer, landowner, pers. comm., 2009). The treatment locations were areas where cattle and buffalo were separated by a common fence. The controls consisted of 2 farms where cattle were surrounded by cattle pastures (no shared wildlife interface) and were at least 10 km away from any buffalo populations. In order to collect both adults and nymphs, adult tick collections were performed in February and March and nymphal collections in July and August.
In order to test the effects of the treatments (i.e. the wildlife/cattle interface), drags (n = 10 per treatment site) were performed in the cattle-dominated area (CB) and the buffalo-dominated area (BC) which were separated by a fence. For the controls, drags (n = 10 per control site) were performed in cattle pastures that were surrounded by other cattle pastures (CC).11
The ticks were collected using a tick drag apparatus consisting of 8 strips of flannel measuring 10 × 10 × 100 cm and weighted at the base with 25 g sinkers11. For each of the cattle-dominated (n = 2), buffalo-dominated (n = 2) and the control sites (n = 2), 10 drags were performed over 250 m with 50 m separating each transect11. Drags were performed in the morning (before 12:00) and in the afternoon (after 14:00). Wherever possible, adult ticks were also collected from buffalo and cattle. These ticks were removed and stored in 70 % ethanol and identified to species. Adult ticks were identified visually from field specimens. Larval and nymph stages were identified using scanning electron microscopy.
The tick community variables of species diversity and evenness were calculated using the program EstimateS® 8.0. Shannon's diversity index (H') was used as the descriptive statistic for diversity and Simpson's D index was used a measure of evenness. A non-parametric Kruskal-Wallis test was performed on these variables along with a multiple comparison of ranks post hoc test using Statistica® 8.0.
RESULTS
There was significantly higher species diversity at the treatment sites (BC, CB) than the control (CC) sites (Table 1, P < 0.001, H2, 60 = 45.4). Nevertheless, the overall species richness was similar across the treatment and control sites (Table 1, P > 0.05, H2, 60 = 17.7). Similarly, there was no significant difference between the treatment and control sites for evenness (Table 1, P > 0.05, H2, 60 = 3.6).
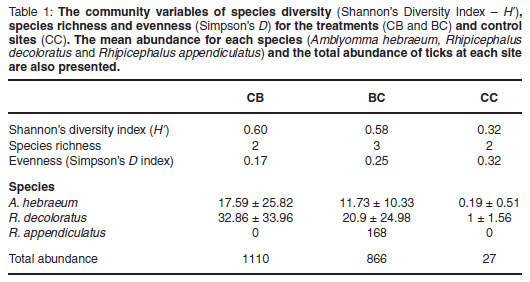
The total abundance data mirrored the species diversity data (Table 1). Total tick abundance was similar at the cattle (CB)and buffalo (BC)-dominated sites (1110 and 866, respectively, P > 0.05) but significantly lower at the control (CC) sites (Table 1, P < 0.001. H2, 60 = 32.9). The single greatest abundance of ticks was found at the cattle-dominated sites (1110 ticks) and the lowest abundance of ticks found at the control sites (27 ticks).
The individual abundance values of Amblyomma hebraeum, Rhipicephalus decoloratus followed a similar trend to the total tick abundance across the treatment and control sites (Table 1). However, and of particular pertinence in this study, R. appendiculatus was only recorded at the buffalo (BC)-dominated sites (Table 1).
The results show that tick species richness, diversity and abundance at the treatment sites were higher than the controls at the wildlife/cattle interface. This can be attributed to 2 major factors: the dipping of the cattle at the cattle-dominated treatment and the visible difference in grass length across the treatments.
DISCUSSION
The fact that the cattle were dipped at the control sites may well have affected the abundance and possibly the species richness and diversity of the ticks. Dipping may have reduced the abundance of ticks found at the control sites. However, evidence of the adaptive ability of ticks to evolve resistance seems to indicate that dipping may not be having an effect on reducing species richness14. It may, however, be affecting the species diversity as some species may evolve resistance more rapidly than others4, 6. The species that rapidly evolve resistance subsequently fill the niches abandoned by less rapidly evolving species and dominate the species diversity8, for example R. decoloratus and A. hebraeum, which were found at both the treatment and control sites. The filling of vacant niches (left by tick species that have not developed resistance) by resistant ticks indicates that it is likely that the overall species richness is not affected in the long term.
Moreover, it does appear that dipping of cattle is controlling the presence of R. appendiculatus. Rhipicephalus appendiculatus was only found in the buffalo-dominated areas of the treatment sites. Most wild animals in Africa have developed high levels of resistance to the parasites transmitted by ticks, therefore it is not common practice to dip wild animals unless they are being transported. In addition, the presence of R. appendiculatus on the buffalo may be a result of poor sanitation practices when transporting buffalo (e.g. not sanitising the transportation crates) into the Eastern Cape7 (C Bissett, Kwandwe Private Game Reserve, pers. comm., 2009).
It may be possible that the ticks in the buffalo-dominated area are more speciose because they are not under the selective influence of dipping. However, the obvious resistance of ticks to dipping seems to cast doubt on this as the only explanation. The patchy distribution of R. appendiculatus has been ascribed to climatic variables such as rainfall and vegetation18, 22. Thus, we contend that it is also the different vegetation structure between the treatment and control sites that is shaping the diversity, richness and abundance of ticks. An investigation into the control of ticks as disease vectors in wildlife in the Ngorongoro Crater, Tanzania, found that the suppression of fire in these areas for 27 years had resulted in the expansion of lush, tall grassland18. With this change in vegetation structure there was a shift in the dominant herbivores from wildebeest (Connochaetes taurinus), hartebeest (Alcelaphus buselaphus) and eland (Tragelaphus oryx) to buffalo. Buffalo prefer grazing taller, lower quality grasses as they supply cover and offer a predator avoidance strategy3.5. In addition, larger herbivores are often at a disadvantage in areas where grass height is low as small herbivores can more efficiently exploit this resource 2, 13. Significantly, buffalo provide excellent hosts for ticks and carry large parasite loads with burdens of up to 1000 R. appendiculatus having been found on a single animal12, 29.
The private game reserves used in our study do not undergo burning regimens. Thus it was visually observed that the grasslands consisted of long, poor quality grass resulting in dense congregations of buffalo in these areas, given their grazing preferences16. Vegetation consisting of long grass provides ideal environments for the completion of tick life cycles across species5. This may explain the increased diversity and richness of the ticks in the buffalo-dominated area. By contrast, ticks have a poor survival rate in short heavily grazed vegetation such as the kind found in the cattle-dominated pastures5. Short vegetation is susceptible to desiccation and damage from higher ultraviolet light intensities making it unfavourable for the development of ticks5. Furthermore, R. appendiculatus, in particular, implements a lifestyle strategy of moving up and down grass stems1. This is naturally restricted in short, over grazed pastures such as those found in the cattle-dominated treatments.
The growing game farming industry in the Eastern Cape means that large numbers of charismatic megafauna such as buffalo are required in order to make game farms economically viable28.The 'disease-free' Addo stock has limited genetic heterogeneity7 and therefore, to ensure genetically sustainable herds, animals must be brought in from other areas of South Africa. Although buffalo populations at both treatments consisted of Addo-derived stock, these animals were sourced from other areas of South Africa. In these areas R. appendiculatus may be more abundant and therefore there is a high risk that they may have been inadvertently transported to the Eastern Cape where R. appendiculatus is not usually abundant14.
The wildlife/cattle interface appears to be having a significant effect on the species diversity, richness and abundance of ticks in the Eastern Cape. Critically, it seems to be increasing the presence and abundance R. appendiculatus. However, the range of R. appendiculatus has not extended dramatically inland, based on previous studies on its distribution29. This species is particularly susceptible to climatic variables such as temperature, rainfall and the resultant vegetation structure alteration. Therefore, it is likely that as climate change alters the environment of the Eastern Cape, R. appendiculatus will extend its distribution further inland. The impact of this is compounded by the effect of the wildlife/cattle interface, which is providing a micro-niche that allows R. appendiculatus to flourish. Owing to these factors it is vital to monitor the 'disease-free' status of buffalo in the Eastern Cape and the distribution of R. appendiculatus. An outbreak of Corridor disease would cripple the livestock industry and the livelihoods of the rural farming majority of the Eastern Cape.
ACKNOWLEDGEMENTS
This work was funded by Rhodes University. We also gratefully acknowledge the cooperation and assistance of Shamwari and Kwandwe Private Game Reserves, Malcolm MacKenzie and Mike Palmer. Charlene Bissett, Murray Stokoe, Jess Watermeyer, Kendall Crous, Tim Richardson and Ben Price are thanked for their assistance in the field. We are particularly grateful to Prof Ivan Horak for help with identification and sampling methods.
REFERENCES
1. Browning T O 1975 The aggregation of questing ticks, Rhipicephalus pulchellus, on grass stems, with observations on Rhipicephalus appendiculatus. Physiological Entomology 1: 107-114 [ Links ]
2. Cumming G S 1999 Theevolutionary ecology of African ticks. D. Phil. thesis, University of Oxford [ Links ]
3. De Graaff G, Schultz K C A, Van der Walt P T 1973 Notes on the rumen contents of Cape buffalo (Syncerus caffer) in the Addo Elephant National Park. Koedoe: 45-58 [ Links ]
4. Foil L D, Coleman P, Eisler M, Fragoso-Sanchez H, Garcia-Vazquez Z, Guerrero F D, Jonsson N N, Langstaff I G, Li A Y, Machila N, Miller R J, Morton J, Preutt J H, Torr S 2004 Factors that influence the prevalence of acaricide resistance in tick-borne diseases. Veterinary Parasitology 125: 163-181 [ Links ]
5. Fyumagwa R D, Runyoro V, Horak I G, Hoare R 2007 Ecology and control of ticks as disease vectors in wildlife of the Ngorongoro Crater, Tanzania. South African Journal of Wildlife Research 37: 79-90 [ Links ]
6. George J E, Pound J M, Davey R B 2004 Chemical control of ticks on cattle and the resistance of these parasites to acaricides. Parasitology 129: S353-S366 [ Links ]
7. Grobler J P, Van der Bank F H 1996 Genetic diversity and isolation in African buffalo (Syncerus caffer). Biochemical Systematics and Ecology 24: 757-761 [ Links ]
8. Herbold B, Moyle P B 1986 Introduced species and vacant niches. American Naturalist 128: 751-760 [ Links ]
9. Hoare D B, Mucina L, Rutherford M C, Vlok J H J, Euston-Brown D I W, Palmer A R, Powrie L W, Lechmere-Oertel R G, Procheº $ M, Dold A P, Ward R A 2006 Albany Thicket Biome. In Rutherford M C, Mucina M C (eds) The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute, Pretoria: 541-567 [ Links ]
10. Hoogstraal H 1956 African Ixodoidea. Research report NM 005050.29.07 for U.S. Naval Medical Research Unit, Cairo [ Links ]
11. Horak I G, Gallivan G J, Spickett A M, Potgieter A L F 2006 The effect of burning on the number of questing ticks collected by dragging. Onderstepoort Journal of Veterinary Research 73: 163-174 [ Links ]
12. Horak I G, Potgieter F T, Walker J B, De Vos V, Boomker J 1983 The ioxid tick burdens of various large ruminant species in South African nature reserves. Onderstepoort Journal of Veterinary Research 50: 221-228 [ Links ]
13. Illius A W, Gordon I J 1987 The allometry of food intake in grazing ruminants. Journal of Animal Ecology 56: 989-999 [ Links ]
14. Lessard P, L'Eplattenier R, Norval, RAI, Kundert K, Dolan T T, Croze H, Walker J B, Irwin A D, Perry B D 1990 Geographical information systems for studying the epidemiology of cattle diseases caused by Theileria parva. Veterinary Record 126: 255-262 [ Links ]
15. Mbati P A, Hlatshwayo M, Mtshali M S, Mogaswane K R, De Waal T D, Dipeolu O 2002 Ticks and tick-borne diseases of livestock belonging to resource-poor farmers in the eastern Free State of South Africa. Experimental & Applied Acarology 28: 217-224.16. [ Links ]
16. McNaughton S J, Georgiadis N J 1986 Ecology of African grazing and browsing mammals. Annual Review of Ecology and Systematics 17: 39-65 [ Links ]
17. Mugabi K N, Mugisha A A, Ocaido M 2009 Socio-economic factors influencing the use of acaricides on livestock: a case study of the pastoralist communities of Nakasongola District, Central Uganda. Tropical Animal Health Production. DOI 10.1007/s11250-0099396 [ Links ]
18. Okello-Onen J, Tukahirwa E M, Perry B D, Rowlands G J, Nagda S M, Musisi G, Bode E, Heinonen R, Mwayi W, Opuda-Asibo J 1999 Population dynamics of ticks on indigenous cattle in a pastoral dry to semi-arid rangeland zone of Uganda. Experimental & Applied Acarology 23: 79-88 [ Links ]
19. Olwoch J M, Reyers B, Englebrecht F A, Erasmus, ReyersBFN 2008 Climate change and the tick-borne disease Theileriosis (East Coast fever) in sub-Saharan Africa. Journal of Arid Environments 72: 108-120 [ Links ]
20. Parker D M 2004 The feeding biology and potential impact of introduced giraffe (Giraffa camelopardalis) in the Eastern Cape Province, South Africa. MSc thesis, Rhodes University [ Links ]
21. Parker D M, BernardRTF 2005 The diet and ecological role of giraffe (Giraffa camelopardalis) introduced to the Eastern Cape, South Africa. Journal of Zoological Society of London 267: 203-210 [ Links ]
22. Perry B D, Lessard P, Norval R A I, Kundert K, Kruska R 1990 Climate, vegetation and the distribution of Rhipicephalus appendiculatus in Africa. Parasitology Today 6: 100-104 [ Links ]
23. Perry B D, Young A S 1995 The past and future roles of epidemiology and economics in the control of tick-borne diseases of livestock in Africa: a case of theileriosis. Preventive Veterinary Medicine 25: 107-120 [ Links ]
24. Potgieter F T, Stoltsz W H, Blouin E F, Roos J A 1988 Corridor disease in South Africa: a review of the current status. Journal of the South African Veterinary Association 59: 155-160 [ Links ]
25. Rechav Y, Zeederberg M E 1986 Rhipicephalus appendiculatus. InSauerJ R, Hair JA (eds) Morphology, physiology, behavioural biology of ticks. John Wiley & Sons, London [ Links ]
26. Sibeko K P, Oosthuizen M C, Collins N E, Geysen D, Rambritch N E, Latif A A, Groeneveld H T, Potgieter F T, Coetzer J A W 2008 Development and evaluation of the real-time polymerase chain reaction test for the detection of Theileria parva infections in Cape buffalo (Syncerus caffer) and cattle. Veterinary Parasitology 155: 37-48 [ Links ]
27. Smallegange I M, BrunstingAMH 2002 Food supply and demand, a simulation model of the functional response of grazing ruminants. Ecological Modelling 149: 179-192 [ Links ]
28. Van der Merwe P, Saayman M 2003 Determining the economic value of game farm tourism. Koedoe 46: 103-112 [ Links ]
29. Walker J B, Keirans J E, Horak I G 2002 The genus Rhipicephalus (Acari: Ixodidae). A guide to the brown ticks of the world. Cambridge University Press, Cambridge [ Links ]
Received: May 2010.
Accepted: November 2010.
* Author for correspondence.E-mail: e.smithsa@gmail.com

