










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkJournal of the South African Veterinary Association
versión On-line ISSN 2224-9435
versión impresa ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.81 no.3 Pretoria sep. 2010
ARTICLE ARTIKEL
Effect of heterologous seminal plasma and semen extenders on motility of frozen-thawed ram spermatozoa
G A MataveiaI; S J TerblancheII,*; J O NöthlingII; D GerberII
ISection of Reproduction, Department of Clinic, Veterinary Faculty, Eduardo Mondlane University, PO Box 257, Maputo, Mozambique
IISection of Reproduction, Department of Production Animal Studies, Faculty of Veterinary Science, University of Pretoria, Private Bag X04, Onderstepoort, 0110 South Africa
ABSTRACT
Ram seminal plasma increases the fertility of frozen-thawed ram spermatozoa deposited into the cervix. The aim of the current study was to compare the effect of ram seminal plasma to that of bull seminal plasma, dog prostatic fluid, protein-free TALP, TrilEq (Triladyl with 0.5 mℓ of Equex STM paste added to each 100 mℓ) and heat-treated skim milk on longevity and percentages of progressively motile and aberrantly motile frozen-thawed ram spermatozoa. Three ejaculates from each of 6 rams were extended in TrilEq, pooled and frozen in straws as a single batch per ram. One hundred and eight straws (3 straws from each ram for each fluid) were thawed in random order. Once thawed, a straw was emptied into a tube with 0.85 mℓ of the appropriate fluid at 37 ºC and kept at that temperature for 6 h. Motility was assessed at ×200 magnification immediately (time zero) and 2, 4 and 6 h after thawing. Progressive motility decreased from each time to the next (P < 0.05) and was 39.0 % (0 h), 26.0 % (2 h), 19.6 % (4 h) and 12.6 % (6 h); SEM 1.24, n = 108 for each group. Ram seminal plasma resulted in higher progressive motility than bull seminal plasma, lower than milk, and similar to the other fluids. Ram seminal plasma resulted in lower aberrant motility than protein-free TALP and similar aberrant motility to other fluids. The effect of ram seminal plasma and dog prostatic fluid was very similar. The effect of ram seminal plasma on the fertility of frozen-thawed ram spermatozoa deposited into the cervix is not due an exceptionally beneficial effect on the motility of spermatozoa.
Keywords: frozen-thawed semen, ram, seminal plasma sperm.
INTRODUCTION
Fresh ram semen may yield good fertility after cervical insemination, whereas frozen-thawed ram semen has to be deposited into the uterus for acceptable fertility. The need for uterine insemination restricts the use of frozen-thawed ram semen because there is no method that is easy, repeatable, non-invasive and that does not require expensive equipment to achieve intrauterine insemination. The addition of ram seminal plasma to frozen-thawed ram spermatozoa holds promise as it improves the fertility achieved with cervical insemination to acceptable levels11. Ram semen contains relatively little plasma, which makes it an impractical additive where large numbers of ewes need to be inseminated with frozen-thawed semen. There is thus a need to search for another fluid that would also yield acceptable fertility of frozen-thawed ram spermatozoa after cervical insemination. Like frozen- thawed ram spermatozoa, frozen-thawed dog spermatozoa also result in high fertility after uterine insemination but poor fertility after vaginal insemination1,7. The addition of dog prostatic fluid to frozen- thawed dog spermatozoa improves their fertility after vaginal insemination12,13. The ejaculate of the domestic dog contains a large amount of seminal plasma, largely derived from the prostate gland2,3. Adding bovine seminal plasma to frozen- thawed African buffalo (Syncerus caffer) spermatozoa improves their ability to fertilise bovine oocytes in vitro, confirming that heterologous seminal plasma can have an effect on the fertility of spermatozoa.6
The motility of ram spermatozoa is considered essential for their migration through the cervix and from the central region of the cervical canal towards the cervical mucosa9 where they are stored and released over time as required for fertilisation10. It is therefore possible that seminal plasma promotes the fertility of frozen-thawed ram spermatozoa deposited in the cervix by maintaining or promoting their motility. When searching for a fluid to replace ram seminal plasma as a post-thaw additive to frozen- thawed ram spermatozoa prior to cervical insemination, its effect on the motility of frozen-thawed ram sperm should be compared to that of ram seminal plasma.
Heat-treated skim milk is an effective extender for fresh ram spermatozoa prior to cervical insemination4, resulting in high fertility, presumably due to its protein fraction, which may buffer against changes in pH5 and partially protect spermatozoa during cooling for storage15. Adding dog prostatic fluid to frozen- thawed dog spermatozoa not only improves their fertility after vaginal insemination12,13, but also their motility and longevity after thawing14. Adding protein-free TALP to frozen-thawed dog spermatozoa, however, maintains their post-thaw motility better than dog prostatic fluid14.
The aim of the current study is to compare the effects of bull seminal plasma, dog prostatic fluid, heat-treated skim milk and protein-free TALP on the motility of frozen-thawed ram spermatozoa to the effect of ram seminal plasma
MATERIALS AND METHODS
Experimental animals
The semen donors were 2 Dorper, 2 Merino and 2 Dohne Merino rams, aged 2-4 years, kept at Faculty of Veterinary Science of the University of Pretoria in open-roofed pens with cement floors and fed lucerne hay and commercial sheep pellets, with water ad libitum. Semen was collected by means of an artificial vagina, using a non-oestrous ewe tied in a neck clamp as a mount. The ejaculates were collected during May, which coincides with the optimum breeding season for sheep in South Africa.
Experimental design
One batch of semen from each of 2 rams from each of 3 breeds was frozen and 18 straws from each batch thawed and 3 straws individually allocated to each of 6 fluids that were added after thawing. In this way, the semen from 3 straws of each batch were separately diluted with the same type of fluid. The fluids were ram seminal plasma, bull seminal plasma, dog prostatic fluid, heat-treated skim milk, protein-free TALP and Triladyl with Equex STM paste (TrilEq). After thawing and dilution the semen was incubated at 37 ºC for 6 h and the motility evaluated zero (t0), 2 (t2), 4 (t4) and 6 (t6) hours after thawing. The response variables were percentage progressively motile spermatozoa, percentage aberrantly motile spermatozoa and percentage immotile spermatozoa.
Collection, preparation and storage of fluid
TrilEq. Triladyl was prepared by adding 1 volume of Triladyl concentrateTM (Minitüb, Tiefenbach, Germany), which contains glycerol, tris, citric acid, fructose, tylosin, gentamicin, lincomycin and spectinomycin in unknown concentrations as this is a trade secret, to 3 volumes of deionised water and 1 volume of egg yolk. Finally, to each 100 mℓ of Triladyl, 0.5 mℓ of Equex STM paste (Nova Chemical Sales, Scituate, MA, USA) was added to give 100.5 mℓ of TrilEq. TrilEq was used as freezing extender and as 1 of the fluids added to frozen-thawed semen. TrilEq was prepared in advance and stored at -70 ºC until shortly before use.
Seminal plasma from rams and bulls and prostatic fluid from dogs. An ejaculate was collected from each of the 6 rams, pooled and centrifuged at 3500 g for 30 minutes and the supernatant filtered through a 0.22 µm filter. This process was repeated and the supernatants pooled until a sufficient quantity was available to yield 20 aliquots of 0.85 mℓ of the sperm free, filtered seminal plasma that were transferred into Cryo-tubes (NUNC, Denmark) and stored at -70 ºC.
One ejaculate from each of 8 bulls (2 Holsteins, 2 Ayrshires, 2 Tulis and 2 Brahmans) belonging to the Taurus Artificial Insemination Centre was collected by means of an artificial vagina and treated in the same manner as described for rams to yield 20 aliquots of seminal plasma.
The post-sperm fraction of 1 ejaculate from each of 6 dogs was collected by means of digital massage and then processed in the same manner as described for seminal plasma of rams and bulls to yield 20 aliquots of prostatic fluid.
Heat-treated skim milk. Ultra high temperature (UHT) skim milk (Clover South Africa (Pty) Ltd) contained 4.9 g carbohydrates, 3.4 g protein, 0.4 g milk fat and 123 mg calcium per litre.
Protein-free TALP. The composition of Protein-free TALP used was: NaCL, 113.96 mmol/ℓ; KCl, 3.19 mmol/ℓ; NaHCO3, 24.88 mmol/ℓ; NaH2PO4 monohydrate, 0.40 mmol/ℓ; MgCl2 hexahydrate, 0.49 mmol/ℓ; CaCl2 dihydrate, 2.00 mmol/ℓ; HEPES, 10.07 mmol/ℓ; Phenol red, 0.01 mmol/ℓ; sodium pyruvate, 0.50 mmol/ℓ; sodium lactate, 15.78 mmol/ℓ; gentamycin, 0.025 g/ℓ 13,16.
Freezing of semen
One ejaculate was collected from each ram on each of 3 consecutive days and discarded, subsequent to which each ram rested for 3 days before another 3 ejaculates were collected from each, with a 60-minute interval between ejaculates. These last 3 ejaculates from each ram were extended, cooled to 5 ºC, pooled and frozen as 1 batch per ram, as described below. Immediately after collection of an ejaculate destined for freezing, a small drop of it was suspended from a glass slide kept at 37 ºC and examined under a microscope at ×40 magnification to confirm that the semen had strong wave movement (at least 4 on a scale from 0 to 5). Each ejaculate with adequate wave movement was immediately extended with 2 volumes of TrilEq at 30 ºC in a 15-m tissue culture tube that was placed in a 50 mℓ water jacket at 25 ºC. This was placed in a beaker with 250 mℓ water at 15 ºC, which was transferred to a cool room at 5 ºC. The 2nd and 3rd ejaculates from the ram were processed in the same way as the 1st and 150 minutes after the 3rd was placed in the cool room they were pooled in a 50 mℓ tissue culture tube. The concentration of spermatozoa in the pool was determined by means of a haemacytometer and the pool further extended with TrilEq at 5 ºC to a final concentration of 200 million per mℓ . Two hours after the 3 ejaculates were pooled the suspension of spermatozoa was inverted repeatedly to ensure thorough mixing. Fifty 0.25 mℓ French straws, marked with the name of the ram, were filled and sealed with PVC powder. The straws were frozen by suspending them horizontally 5 mm apart and 4 cm above liquid nitrogen in stable nitrogen in a Styrofoam box for 20 minutes before plunging them into the liquid nitrogen. The straws were then stored in a liquid nitrogen container at -196 ºC.
Evaluation of sperm motility
In order to determine the sequence in which ejaculates and fluids had to be thawed, a random number was allocated to each ram-by-fluid-by-straw combination. The ram-by-fluid-by-straw combinations were then sorted in ascending order according to the random numbers, after which the straws were thawed, diluted, incubated and evaluated in the order of ascending random numbers.
One person thawed all straws and prepared all slides for the assessment of motility, using glassware kept at 37 ºC, and another person performed all the assessments. Ten minutes before a straw with semen was thawed, the fluid destined for it was thawed and warmed to 37 ºC. As soon as a straw with semen had thawed, its contents were transferred to a 4.5 mℓ centrifugation tube and diluted with 0.85 mℓ of the appropriate fluid. The person then placed a drop of the diluted semen onto the centre of a cover slip that was then inverted and placed on a glass slide labelled with the random number. This person then handed the slide over to the other who placed the slide on the warm stage (37 ºC) of a phase-contrast microscope at ×200 magnification in order to assess the motility. The 2nd person estimated the percentages of progressively motile, aberrantly motile and immotile spermatozoa in 10 neighbouring fields along the equator of the coverslip, starting at the edge and ending at the centre14. From the results obtained average values were calculated. The individual sperm motility is expressed in terms of percentages of progressively motile, aberrantly motile and immotile spermatozoa.
The motility of the semen was estimated within 5 minutes after dilution (t0), and again 2, 4 and 6 hours later (t2, t4, t6).
The straw with the next higher random number was thawed and diluted 10 minutes after the previous one, and so on until all straws were evaluated.
The tubes containing the extended semen were capped and placed in a water bath at 37 ºC until after the last evaluation at t6.
Statistical analysis
The statistical software package NCSS (www.ncss.com, Kaysville, UT USA) was used for all statistical analyses. Breed was initially included in the model, with ram nested in breed, but, because breed had no significant effect (P > 0.05) and breed was of no particular concern with respect to the aim of the study, it was removed prior to the final analysis.
A repeated measures ANOVA was used to test for differences among the postthaw motility with the rams as subjects and fluid and time as factors causing within-subjects variation. The interaction between fluid and time was included in the model, whereas the subject interactions were combined with error. Bonferroni's test was used to compare the mean of each fluid to that of ram seminal plasma, which served as control8. For all comparisons the level of significance was set to P < 0.05.
RESULTS
Fresh semen quality
Although there was a large variation in sperm concentration among rams, each ejaculate had good motility (mass motility score 5, percentage progressively motile spermatozoa exceeding 70 % and percentage aberrantly motile spermatozoa of 1 % or lower).
Motility after thawing and dilution
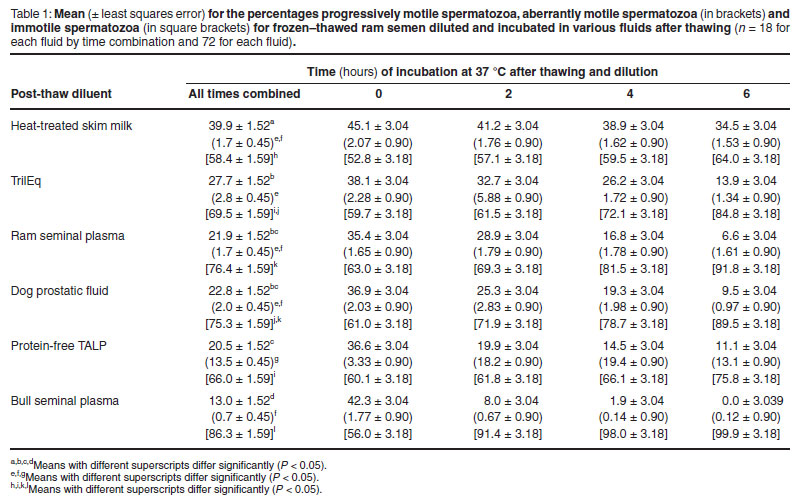
Ram, fluid and time significantly affected the percentage progressively motile spermatozoa, the percentage aberrantly motile spermatozoa and the percentage immotile spermatozoa (P < 0.001). Table 1 shows the mean percentages of progressively motile, aberrantly motile, and immotile spermatozoa for each fluid at different times of incubation. Ram seminal plasma resulted in a higher percentage progressively motile spermatozoa than bull seminal plasma and a lower percentage than heat-treated skim milk (P < 0.05), and a similar percentage to the other fluids (P > 0.05, Table 1). Ram seminal plasma resulted in a lower percentage aberrantly motile spermatozoa than protein-free TALP (P < 0.05), and a similar percentage to the other fluids. For all times combined, ram seminal plasma resulted in similar percentage immotile spermatozoa to dog prostatic fluid, which was lower than the percentage for bull seminal plasma, and higher than the percentages for other fluids (P < 0.05). Heat-treated skim milk resulted in a higher percentage progressively motile spermatozoa and a lower percentage immotile spermatozoa than all other fluids (P < 0.05).
The effect of ram seminal plasma and dog prostatic fluid on motility was strikingly similar at each time after the onset of incubation, as well as over all times combined (Table 1).
Fluid interacted with time with respect to percentage progressively motile spermatozoa, the percentage aberrantly motile spermatozoa and the percentage immotile spermatozoa (P < 0.001).
The interaction between fluid and time with respect to percentage progressively motile spermatozoa was mainly because the percentage progressively motile spermatozoa in spermatozoa samples diluted with bull seminal plasma declined more steeply during the 1st 2 hours of incubation than it did for spermatozoa diluted in other fluids. Fluid interacted with time with respect to the percentage aberrantly motile spermatozoa mainly because, for protein-free TALP, the percentage spermatozoa with aberrant motility increased by about 15 % during the 1st 2 hours of incubation and remained at a similar level during the following 4 hours, while no such rise occurred with the other fluids. The interaction between fluid and time with respect to percentage immotile spermatozoa was mainly because, for bull seminal plasma, the percentage immotile spermatozoa increased more steeply during the 1st 2 hours of incubation than for the other fluids.
DISCUSSION
Although ram seminal plasma improves the fertility of frozen-thawed ram spermatozoa deposited into the cervix11 and heat-treated skim milk does not, ram seminal plasma does not preserve the in vitro motility of such spermatozoa as well as heat-treated skim milk. This observation suggests that ram seminal plasma exerts its beneficial effect on the fertility of frozen-thawed ram spermatozoa deposited into the cervix via a mechanism other than exceptional preservation of progressive motility.
Assuming that ram seminal plasma preserves progressive motility of frozen- thawed spermatozoa that were deposited into the cervix sufficiently well to result in acceptable fertility, the results of the current study suggest that dog prostatic fluid and protein-free TALP, both of which preserved progressive motility as well as ram seminal plasma, should also adequately preserve their progressive motility. In the dog, however, vaginally inseminated frozen-thawed spermatozoa resulted in higher fertility after the addition of dog prostatic fluid than protein-free TALP13. The observation that seminal plasma of the domestic bull improves the ability of African buffalo spermatozoa to fertilise bovine oocytes in vitro6 suggests that it is reasonable to suspect that effects of seminal plasma on fertility need not be restricted to an autologous effect.
Sheep and dogs share certain characteristics with respect to the utility of frozen- thawed spermatozoa in the 2 species: fertility resulting from deposition of frozen-thawed spermatozoa is good after deposition in the uterus, but poor after deposition at the external cervical opening, and fertility after deposition at the latter site is better if seminal plasma is added to the frozen-thawed spermatozoa prior to insemination.
Further research is required to compare the effects of ram seminal plasma and dog prostatic fluid on the fertility of frozen- thawed ram spermatozoa deposited into the cervix.
REFERENCES
1. Anderson K 1975 Insemination with frozen dog semen based on a new insemination technique. Zuchthygiene 10: 1-4 [ Links ]
2. EnglandGCW, Allen W E 1989 Seminal characteristics and fertility in dogs. Veterinary Record 125: 399 [ Links ]
3. EnglandGCW, Allen W E, Middleton D J 1990 An investigation into the origin of the first fraction of the canine ejaculate. Research in Veterinary Science 49: 66-70 [ Links ]
4. Evans G, Maxwell W M C 1987 Salmon's artificial insemination of sheep and goats. Butterworths, Sydney [ Links ]
5. Foote R H 1964 Influence of pH on survival and fertility of bull sperm. Journal of Dairy Science 47: 807-811 [ Links ]
6. Gerber D, De Haas K, Nöthling J O 2003 Effect of bovine seminal plasma on the ability of buffalo (Syncerus caffer) spermatozoa to fertilize bovine oocytes in vitro. IETS Meeting, Auckland, New Zealand, 11-15 January: Theriogenology 59: 392 [ Links ]
7. Linde-Forsberg C, Ström-Holst B, Govette G 1999 Comparison of fertility data from vaginal versus intrauterine insemination of frozen-thawed dog semen: a retrospective study. Theriogenology 52: 11-23 [ Links ]
8. Maroco J 2004 Análise Estatística com utilização do SPSS (2a Edição). Edições Sílabo LDA, Lisboa, Portugal [ Links ]
9. Mattner P E, BradenAWH 1969 Comparison of the distribution of motile and immotile spermatozoa in the ovine cervix. Australian Journal of Biological Science 22: 1069-1069 [ Links ]
10. Mattner P E 1966 Formation and retention of the spermatozoan reservoir in the cervix of the ruminant. Nature 212: 1479-1480 [ Links ]
11. Maxwell W M C, Evans, G, Mortimer, S T, Gillan L, Gellatly E S, McPhie C A 1999 Normal fertility in ewes after cervical insemination with frozen-thawed spermatozoa supplemented with seminal plasma. Reproduction Fertility & Development 11: 123-126 [ Links ]
12. Nöthling J O, Volkmann D H 1993 Effect of addition of autologous prostatic fluid on the fertility of frozen-thawed dog semen after intravaginal insemination. Journal of Reproduction and Fertility Supplement 47: 329-333 [ Links ]
13. Nöthling J O, Shuttleworth R, de Haas K, Thompson P N 2005 Homologous prostatic fluid added to frozen-thawed dog spermatozoa prior to intravaginal insemination of bitches resulted in better fertility than albumin-free TALP. Theriogenology 64: 975-991 [ Links ]
14. Nöthling J O, Gerber D, Colenbrander B, Dijkstra M, Bakker T, De Cramer K 2007 The effect of homologous prostatic fluid on motility and morphology of dog epididymal spermatozoa extended and frozen in Biladyl with Equex STM paste or Andromed. Theriogenology 67: 264-275 [ Links ]
15. O'Shea T, Wales R G 1966 Effect of casein lecithin, glycerol and storage at 5 ºC on diluted ram and bull semen. Australian Journal of Biological Science 19: 871-882 [ Links ]
16.YingY,ChowPH,CheungMPL,O W S 1999 Male accessory sex gland secretions affect oocyte Ca2+ oscillations during in vitro fertilization in golden hamsters. Molecular Human Reproduction 5: 527-533 [ Links ]
Received: November 2009.
Accepted: April 2010.
* Author for correspondence. E-mail: veepos@gmail.com


{kind=link}