










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkJournal of the South African Veterinary Association
On-line version ISSN 2224-9435
Print version ISSN 1019-9128
J. S. Afr. Vet. Assoc. vol.81 n.1 Pretoria Jan. 2010
ARTICLE ARTIKEL
The ovarian and uterine arteries in the chinchilla (Chinchilla lanigera)
A Çevik-DemirkanI,*; V ÖzdemirI; I DemirkanII
IDepartment of Anatomy, Faculty of Veterinary Medicine, Afyon Kocatepe University, 03200, Afyonkarahisar, Turkey
IIDepartment of Surgery, Faculty of Veterinary Medicine, Afyon Kocatepe University, 03200, Afyonkarahisar, Turkey
ABSTRACT
The purpose of this study was to describe arteries supplying the ovaries and uterus in the chinchilla. Five healthy adult female chinchillas were used. In order to reveal the arterial network by dissecting under a stereoscopic microscope, latex coloured with red ink was injected through the common carotid artery. The ovaries of the chinchilla are supplied by the arteriae ovaricae which formed end-to-end anastomoses with the cranial termination of the arteria uterina. Soon after leaving the aorta abdominalis, the arteriae ovaricae extended 2-3 mm caudolaterally, then released 1 branch and extended caudally and bifurcated into 2 further branches. One of these supplied branches to fat tissue. The other branch coursed caudally and anastomosed with the arteria circumflexa ilium profunda and dispersed into fat tissue. The arteria ovarica further subdivided into 2 rami ovaricae. The origins of the uterine arteries were exclusively from the left arteria iliaca externa. The arteria uterina gave a branch to the arteria umbilicalis and consecutive branches which supplied to the ureter, urinary bladder and cranial aspects of the vagina. It also gave rise to 2-3 branches to the cervix and further supplied 10-12 meandering branches to the uterine horns. The arteria uterina gave rise to many tortuous arteries to the uterus and provided 2 further branches to the ovary.
Keywords: anatomy, artery, chinchilla, ovaries, uterus.
INTRODUCTION
Chinchilla lanigera is a rodent species belonging to the family Chinchillidae10 and originates from the central Andes in Chile, Peru and Bolivia in South America15. They are kept for fur, as pets or laboratory animals20.
The female genital organs of mammals consist of ovaries, fallopian tubes, a uterus and a vagina. The ovaries lie on the dorsal wall of the abdominal cavity and caudal to the kidneys3,5,14. The ostium is the funnel-shaped opening of the fallopian tubes. Fimbriae tubariae extend as fingerlike projections from the edge of the ostium. The fallopian tubes are small, short and highly coiled, extending between ovaries and uterus. In rabbits the uterus is divided into 2 completely separate uterine horns that open caudally into the vagina through a single cervix that projects into the vagina14. Similarly, the uterus of the chinchilla is duplex and the uterine horns open by 2 lateral slit-like structures22.
Vascularisation of the uterus and ovaries in Lagomorpha, rodents and domesticated animals is described in detail in many textbooks3,5,13,14. The arteria ovarica runs along the dorsal abdominal wall into the mesovarium and the right artery crosses the vena cava caudalis ventrally. It gives rise to the ramus tubarius which lies in the mesosalpinx, and supplies the uterine tube by means of several meandering branches in mammals16. In many species, including pigs, hamsters, rats and rabbits, the ovarian and uterine arteries anastomose near the ovary to provide a dual or parallel supply13. The ovarian artery is generally assumed to supply the ovaries and also the uterus9,13. The arteria ovarica arises from the aorta abdominalis in mammals and rodents. The arteria uterina originates from the arteria umbilicalis and runs through the mesometrium in rabbits, pigs and ruminants3,16 whereas the uterine artery stems from the external iliac artery in horses16.
Our literature search on the arterial vascularisation of ovaries and uterus in the chinchilla was unrewarding. Thus, this paper describes the course and anastomosis of the arteria uterina and arteria uterina in the chinchilla.
MATERIALS AND METHODS
Five healthy, adult female, non-pregnant chinchillas (Chinchilla lanigera) obtained from the Center for Experimental Medicine, Research and Application, Afyon Kocatepe University, Turkey, were used in this study. The live body weight of chinchillas varied between 450 g and 500 g. The animals were euthanased by the methods described by Flecknell7. Regulations of the ethical committee of Afyon Kocatepe University were followed. Following euthanasia, 1 m of heparine sodium (Nevparin, Mustafa Nevzat, Istanbul, Turkey) was immediately injected via the jugular vein to prevent blood coagulation. Animals were bled by cutting the jugular vein. The vessels were flushed with warm isotonic saline solution until the tissues were blanched. Latex coloured with red ink was injected through the arteria carotis communis. The cadavers were fixed in 10% formaldehyde solution for 24 hours at room temperature. To observe the arteria ovarica and the arteria uterina, the broad ligament and the peritoneal tissue adjacent to the ventral midline of the abdominal cavity were dissected carefully under a stereo-microscope. The photographs were taken using a digital camera (Sony DSC F-717, Japan).
of heparine sodium (Nevparin, Mustafa Nevzat, Istanbul, Turkey) was immediately injected via the jugular vein to prevent blood coagulation. Animals were bled by cutting the jugular vein. The vessels were flushed with warm isotonic saline solution until the tissues were blanched. Latex coloured with red ink was injected through the arteria carotis communis. The cadavers were fixed in 10% formaldehyde solution for 24 hours at room temperature. To observe the arteria ovarica and the arteria uterina, the broad ligament and the peritoneal tissue adjacent to the ventral midline of the abdominal cavity were dissected carefully under a stereo-microscope. The photographs were taken using a digital camera (Sony DSC F-717, Japan).
The latest edition of the Nomina Anatomica Veterinaria was used for the terminology17. The figures in Popesko et al.19 were also used as a guide for the nomenclature.
RESULTS
The ovaries were exclusively supplied by the arteria ovarica. These arteries originated either from the aorta abdominalis (Figs 1a, 4a) at about the level of the origin of the arteria renalis or slightly caudal to it. (Figs 1b, 4b). The right arteria ovarica (Figs 1c, 4c) and the left arteria ovarica (Figs 1c', 2e, 4c') arose from the aorta 3 mm caudal to the arteria renalis.




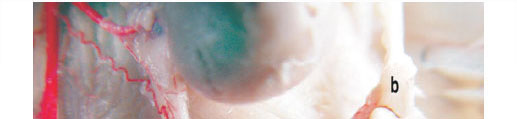




























Each of the arteria ovarica passed obliquely caudad across the ventral surface of the psoas muscle and the ureter. The ovarian rami formed meandering branches 2-3 mm from the aorta. Soon after leaving the aorta abdominalis, the arteria ovarica extended caudolaterally for a distance of approximately 2-3 mm, then produced 1 thin branch, which extended caudally and subdivided into 2 branches (Figs 1d, 4d). The course of these 2 branches was straight. One of these supplied rami to the fat tissue and the other branch ran caudally and anastomosed with the arteria circumflexa ilium profunda and dispersed into fat tissues in this region. The arteria ovarica gave off 2 rami tubarii (with an angle of 30º between them) (Figs 2a, 4e), and then continued as the ramus uterinus (the main trunk). One of the branches (cranial branch) of ramus tubarius extended to fimbriae tubariae (Figs 2b, 4h) and infundibulum of the fallopian tubes. The other branch (caudal branch) from the ramus tubarius extended to the hilus of the ovary (Figs 2c, 4i) and produced 2-3 branches while entering the hilus. Before entering the hilus of the ovary, the caudal branch anastomosed with 1 branch coming from the uterine artery. The ramus uterinus ran more caudally to form a major end-to-end anastomosis with the cranial termination of the arteria uterina (Figs 2d, 4f).
The aorta abdominalis bifurcated into the left and right arteria iliaca communis (at the level of 5th and 6th lumbar vertebrae) (Figs 3a, 4p), each of which further subdivided into the external and internal iliac arteries. Arteria iliaca communis continued in a caudolateral direction on the dorsal aspect of the uterus. It bifurcated into the arteria iliaca externa and the arteria iliaca interna within 7 mm on the right and 9 mm on the left beyond the aorta (Figs 3b,c, 4q, 4/r). The arteria uterina originated from the external iliac arteries (Figs 3d, 4s). Initially, the arteria uterina gave off a branch to the arteria umbilicalis (Figs 3e, 4t) and supplied 1 branch to both the ureter (Fig2 3f, 4o), urinary bladder (Figs 3g, 4n) and cranial aspects of the vagina. The uterine artery passed medially toward the cervix where it gave rise to 2-3 branches to the cervix (Figs 3h, 4m). The arteria uterina then continued craniomedially on the lateral border of the uterus. Here, it supplied 10 to 12 meandering branches (each contained 3-4 arteries) (Figs 2h, 4g) to the caudal (n = 2 or 3 arteries) (Figs 3i, 4l) and cranial aspects of the uterine horns (n = 8 or 9 arteries) (Figs 2g, 3j, 4k). A bundle emerged from the posterior end of the ovary and continued to the anterolateral end of the uterus (ligamentum ovarii proprium) (Figs 2f, 4j). The arteria uterina gave origin to many tortuous arteries which supplied blood to the uterus. The arteria uterina gave off 2 further branches to the ovary. One of the branches extended to the mesovarium (subdivided into a thin ramus that merged with the ramus tubarius). The other branch extended to the isthmus tubae uterinae.
DISCUSSION
This study demonstrated that the ovarian and uterine arteries, throughout their course, were in close apposition, and sometimes showed various degrees of coiling in the chinchilla. This close anatomical relationship was described previously in other species, such as guinea-pigs9, rats13, rabbits11,18, mice6, primates such as Rhesus monkeys8 and, baboons21.
The arteria ovarica, in the chinchilla, originated from the aorta abdominalis about 2-3 mm from the caudal aspects of right and left arteriae renalis. However, in guinea pigs, the arteria ovarica originated either from the aorta at about the level of origin of the arteria renalis, direct branches of arteria renalis, or common trunks with arteria renalis and in some cases formed anastomoses with branches of the arteria renalis9. In rabbits, it arose from the aorta abdominalis, immediately behind the origin of the inferior mesenteric artery3, whereas it left the abdominal aorta slightly caudal to the arteria renalis in mice6, New Zealand rabbits11 and in rats2,23. Each of the ovarian arteries after emerging from the aorta abdominalis, divided into 2 branches, the caudal branches served the Fallopian tubes and uterine horns, and the cranial branch mainly supplied blood to the ovary in guinea pigs4,8,9, rats4,8, mice6 and in New Zealand rabbits11. Similar structures were observed in our study. The arteria ovarica produced 1 branch which extended caudally and subdivided into 2 rami. One of these supplied blood to fat tissue, the other branch went caudally and anastomosed with the arteria circumflexa ilium profunda and dispersed into fat tissue in this region. Later, the arteria ovarica subdivided into the ramus tubarius and the ramus uterinus. According to Hossain and O'Shea9, in the guinea pig the ramus tubarius produced a total of 5 or 6 small, tightly coiled rami before reaching the hilus of the ovary, 3 or 4 of which entered the hilus; in the chinchilla the arteria ovarica produced 2 rami tubariae, 1 of which reached to the hilus of the ovary and the other ran to the fimbriae tubariae and infundibulum of the Fallopian tubes.
In chinchillas, the arteria uterina originated exclusively from the external iliac artery, while it arose from the arteria vaginalis (arising from the arteria iliaca interna) in guinea pigs19 and coendou12, and was always the 1st branch of the arteria iliaca interna in rabbits1 and mice6; but in rats the arteria uterina originated from the umbilical artery23. The arteria uterina, in chinchillas, produced the arteria umbilicalis and sent branches to the ureter, urinary bladder and cranial vagina. After the uterine artery passed medially toward the cervix (where it gave rise to 2-3 branches to the cervix), it supplied 10-12 bundles of arteries to the uterus. A similar pattern was reported in other rodents6,9. In chinchillas, an anastomosis between the ateria uterina and the arteria ovarica was observed as previously reported in species of rodents, such as guinea pigs9, rats, hamsters2,4, mice6 and porcupines12.
In conclusion, the arterial vascularisation of the ovaries and the uterus was provided by the ovarian and uterine arteries in the chinchilla. Two branches, one arising from the the arteria ovarica and the other from the arteria uterina entered the hilus of the ovary. The arteria uterina originated from the arteria iliaca externa, and served as a source of blood supply to the ovaries through the anastomoses between the terminal ends of the uterine and ovarian arteries. It was hoped that the data generated here could be useful for those who are interested in surgery (especially in experimental studies), diseases, or clinical treatment of chinchillas.
REFERENCES
1. Binkert C A, Andrews R T, Pavcnik D, Uchida B, Loriaux M M, Rösch J 2002 Lack of suitability of the rabbit model for particulate uterine artery embolization. Journal of Vascular and Interventional Radiology 13:609-612 [ Links ]
2. Chiasson R B 1980 Laboratory anatomy of the white rat. Wm C Brown Publishers, Dubuque, Iowa [ Links ]
3. Craigie E H 1969 Bensley's practical anatomy of the rabbit (8th edn). Toronto Press, Canada. [ Links ]
4. Del Campo C H, Ginther O J 1972 Vascular anatomy of uterus and ovaries and unilateral luteolytic effect of the uterus: Guineapigs, rats, hamsters and rabbits. American Journal of Veterinary Research 33:2561-2578 [ Links ]
5. Dyce K M, Sack W O, Wensing C J G 1996 Textbook of veterinary anatomy (2nd edn). W B Saunders Company, Philadelphia [ Links ]
6. El-Akouri R R, Kurlberg G, Dindelegan G, Mölne J 2002 Heterotopic uterine transplantation by vascular anastomosis in the mouse. Journal of Endocrinology 174:157-166 [ Links ]
7. Flecknell P A 1980 Laboratory animal anaesthesia. Academic Press, London [ Links ]
8. Ginther O J, Dierschke D J, Walsh S W, Del Campo C H 1974 Anatomy of arteries and veins of uterus and ovaries in Rhesus monkeys. Biology of Reproduction 11:205-219 [ Links ]
9. Hossain M I, O'Shea D O 1983 The vascular anatomy of the ovary and the relative contribution of the ovarian and uterine arteries to the blood supply of the ovary in the guinea-pig. Journal of Anatomy 137:457-466 [ Links ]
10. Kuru M 1987 Omurgali Hayvanlar. Ataturk University Press, Erzurum, Turkey [ Links ]
11. Macchiarelli G 2000 The microvasculature of the ovary: a review by SEM of vascular corrosion cast. Journal of Reproduction and Development 46:207-225 [ Links ]
12. Machado G V, Gonçalves P R, Parizzi A, Miglino M A, Santos T C 2001 Blood supply of the coendou (Sphiggurus spp. Cuvier, 1825) (Rodentia: Mammalia) uterus and ovaries. Archives of Veterinary Science 6(1). Online at: http://ojs.c3sl.ufpr.br/ojs2/index.php/veterinary/issue/view/392 [ Links ]
13. Massa H M, Bruce N W 1997 Direction of blood flow and changes in resistance of major arteries supplying the ovary of the pregnant rat. Biology of Reproduction 57:43-48 [ Links ]
14. McLaughlin C A, Chiasson R B 1990 Laboratory anatomy of the rabbit. Briston, McGraw Hill, Toronto. [ Links ]
15. Meadow H 1969 Chinchilla. Tozer, Redwood City, CA [ Links ]
16. Nickel R, Schummer A, Seiferle E 1981 The circulatory system, the skin, and the cutaneous organs of the domestic mammals, Vol.3. Verlag Paul Parey, Berlin-Hamburg [ Links ]
17. Nomina Anatomica Veterinaria (5th edn) 2005 International Committee on Veterinary Gross Anatomical Nomenclature. World Association of Veterinary Anatomists, Hannover, Germany [ Links ]
18. Nottola S A, Macchiarelli G, Motta P M 1997 The angioarchitecture of estrous, pseudo-pregnant and pregnant rabbit ovary as seen by scanning electron microscopy of vascular corrosion cats. Cell and Tissue Research 288:353-363 [ Links ]
19. Popesko P, Rajtova V, Horak J 1992 A colour atlas of the anatomy of small laboratory animals, Vol.1, Rabbit and guinea pig. Wolfe Publishing, Bratislava [ Links ]
20. Poyraz Ö 2000 Laboratory animal science. Kardelen Offset, Ankara, Turkey [ Links ]
21. Shaikh A A, Shaikh S A, Celaya C L 1981 Utero-ovarian vein blood collection by the use of a laparoscope in the baboon. Biology of Reproduction 24:455-459 [ Links ]
22. Tibbitts F D, Hillemann H 1959 The development and histology of the chinchilla placenta. Journal of Morphology 105:317-365 [ Links ]
23. Walker W F, Homberger D G 1998 Anatomy and dissection of the rat. W H Freeman, New York [ Links ]
Received: February 2009.
Accepted: September 2009.
* Author for correspondence. E-mail: aysuncevik@yahoo.com

