










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 n.5 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i5.15
Effects of rumen undegradable protein source on feed intake and milk yield of Holstein dairy cows
M. SavariI; M.S.S. FirouzabadiII, #; F. MohammadiI; R. AbigabaIII; A. R. RayshanIV; A. GharagozlooV; F. AbdollahzadehVI
IDepartment of Animal Science, College of Agriculture, Isfahan University of Technology, Isfahan 84156-83111, I. R. Iran
IIDepartment of Clinical Science, Faculty of Veterinary Medicine, Ardakan University, P.O. Box 184, Ardakan, lran
IIIDepartment of Biomolecular and Biolab Sciences, Collage of Veterinary Medicine, Animal Resources & Biosecurity, Makerere University, P.O. Box 7062, Kampala, Uganda
IVScientific Affairs Department, University of Al-Qadisiyah, Iraq
VDepartment of Animal Science, Shahrekord Branch, Islamic Azad University, Shahrekord, Iran
VIDepartment of Animal Science, College of Agriculture, University of Tabriz, Tabriz, Iran
ABSTRACT
This study was conducted to determine the effects of heat treated-soybean meal, fish meal, meat meal, and an equal blend of fish and meat meal as rumen-ungradable protein (RUP) sources on productive performance, nutrient digestibility, N-utilization efficiency, and blood metabolites in high-producing Holstein dairy cows. Twelve lactating Holstein cows (DIM = 89 ± 11; mean ± SD) were used in a replicated 4 × 4 Latin square design receiving four diets formulated with amounts (DM basis): (1) 3.1% heat treated-soybean meal (T-SBM), (2) 2.5% fish meal (FM), (3) 3.0% meat and bone meal (MM), and (4) an equal blend of 1.4% FM plus 1.4% MM (FMM), as dietary RUP sources. The crude protein concentration was kept constant across the treatments (16.7% of DM). The duration of each experimental period was 21 d with 15 d devoted to dietary adaptation and 6 d to data collection. Dry matter intake (24.2 kg/d), actual milk (41.8 kg/d), 4% fat-corrected milk yield (35.6 kg/d), milk composition (Fat = 3.03%, Protein = 2.90%), and blood metabolites were not affected by treatments. Apparent N efficiency (29.5%) was similar across the diets. Based on our results, T-SBM, FM, MM, and mixed of FM and MM had a similar effect on productivity and N-utilization efficiency in lactating dairy cows. Based on the income over feed cost (IOFC), under our experimental conditions, use of the meat meal as a source of dietary RUP can be recommended as a feeding strategy for feed to lactating cows receiving high-concentrate diets.
Key words: dairy cow, nitrogen efficiency, rumen-undegradable protein
Introduction
Protein can increase milk yield by providing more amino acids, increasing available energy, and altering efficiency of utilization of absorbed nutrients (Fadel et ai., 2017). The goal in protein nutrition should be optimizing N efficiency, which means minimizing total N intake, while simultaneously meeting the requirements for milk protein synthesis and, thereby, reducing feeding costs and N excretion into the environment (Savari et ai., 2018). Protein is one of the most important and expensive components of the diet and is considered as nutrient-limiting, especially in high-yielding cows (Binh Truong and Thanh Trung, 2023). Finding an efficient way of converting animal and plant proteins into milk production is the key to improve farm profitability and decrease the environmental emissions of dairy farms.
A part of the crude protein consumed by the animal is degraded in the rumen by the rumen microbes and can be incorporated into microbial protein entering the small intestine. This part of the protein that is synthesized by the microbes has the best amino acid profile to meet the needs of the animal (Schwab, 1994). Another portion of the crude protein consumed by the animal escapes ruminal fermentation and goes directly into the small intestine and is absorbed by the intestine as an amino acid (Jones et ai., 1988; Broderick, 2006). Ruminally-synthesized microbial CP (MCP), RUP, and, to a much lesser extent, endogenous CP, contribute to the passage of metabolizable protein (MP) into the small intestine (NRC, 2001).
Increasing milk production leads to an increased feed intake and animal protein requirements. Due to rumen restrictions in the production of microbial protein, the cow will need rumen undegradable protein sources to provide metabolizable protein. In high-producing cows, MCP supplies a lower portion of the protein requirement; therefore, substantial amounts of the dietary protein must escape ruminai degradation to meet the animal's protein requirements (Broderick, 2006b; Abdollahzadeh et ai., 2020). If the dietary protein is well balanced, the efficiency of dietary to milk protein conversion will be more than 30% but when the dietary CP is imbalanced, it will be decreased to 23% (Broderick, 2006; Schwab et ai., 2007).
Soybean meal is the most commonly used protein supplement for dairy and beef cattle and has a good amino acid profile; however, most of its protein content can be degraded by rumen microbes, leading to a surplus ammonia production in the rumen (Mielke & Schingoethe, 1981). The aim of SBM processing is to reduce ruminal degradability for high quality protein supplements with no negative effects on its intestinal digestibility. However, with the increased demand for treated-SBM and thus its elevated price in recent years, there have been motivation for animal nutritionists to find potential alternatives to SBM with minimum adverse effects on the productive performance of dairy cows. Animal protein sources are high in protein content and generally much more resistant to microbial degradation in the rumen than most of the commonly-fed plant protein sources. In reviewing studies on protein sources in animal nutrition, a significant advantage of FM and MM supplementation in ruminant diets was found, if the physiological status of the animal and the quality of the dietary ingredients were taken into account (Santos et ai., 1998; Hussein & Jordan, 1991). Caution is suggested in using animal protein feedstuffs since palatability, cost per unit of protein, consistency of product, and impact on animal performance are key factors to successful dairying. Using cheaper protein sources, such as meat meal, instead of expensive sources, such as treated-soybean meal and fish meal, can reduce feed costs (NRC, 2001). There is evidence suggesting that combinations of highly undegradable protein sources give better results when fed in conjunction with adequate degradable protein. A study was designed and conducted to determine the proper source of rumen undegradable protein according to the diets commonly used in feeding high-yielding cows, as re-evaluating the efficiency of RUP sources seems to be one way of improving the production status of dairy farms.
Material and Methods
This experiment was conducted at the Dairy Facilities of the Lavark Research and Teaching Farm (Isfahan University of Technology, Isfahan, Iran). Animals were cared for according to the guidelines of the Iranian Council of Animal Care (1995). This study was conducted using 12 high-producing Holstein cows (second parity; 89 ±11 DIM; milk yield = 48 ±3 kg/d) in a replicated 4 × 4 Latin square design with four treatments and three replicates (cows) during four periods of 21 d (16 d for dietary adaptation and 5 d for data collection). To minimize carryover effects, the animals within the squares were randomly assigned to one of four experimental dietary sequences. Cows were housed individually in box stalls (4 m × 4 m) located in a roofed area with open sides and had free access to water. Each box stall was equipped with a concrete feed bunk and automatic water trough. Clean wood shavings and sand were used for bedding and refreshed daily.
The cows were weighed at the beginning and end of each period (at 02:00). The feed was manually mixed and weighed, and diets were offered as a TMR twice daily (at 10:00 and 18:00) for ad iibitum intake allowing for 10% refusals with free access to water. In this study, treated soybean meal [RUP source that we used in our previous study (Savari et ai., 2018)], fish meal, meat meal, and an equal blend of fish and meat meal were used to determine the effects of RUP source on productive performance, nutrient digestibility, N-utilization efficiency, and blood metabolites in high-producing Holstein dairy cows. All diets were formulated using NRC (2001) for a multiparous, lactating dairy cow (600 kg BW) producing 48 kg/d of milk containing 3.0% true milk protein and 3.2% milk fat. Moreover, the diets were almost identical in composition except for the source of RUP.
The amounts of feed offered and orts were measured daily for each cow during the last five consecutive days of each period, and DMI was recorded daily. Samples were collected for each TMR and of individual refusals during the 5-d sample collection period. Samples of other feed ingredients (pooled within a period) were also collected. All the samples were stored at -20 °C until analysis. Samples of feeds and orts were dried at 60 °C for 48 h to determine DM content. Dried samples were then ground in a Wiley mill and passed through a 1-mm screen (Arthur H. Thomas, Philadelphia, PA) before analysis to determine CP, ether extract, and ash, according to AOAC International (2002). Heat stable α-amylase (100 μL/sample) was used to determine NDF, and the ADF content was determined sequentially according to the method of Van Soest et al. (1991) using an Ankom 200 fibre analyser (Ankom Technology Corp., Macedon, NY). The RUP and RDP of feed ingredients were calculated according to the method of Sniffen et al. (1992) and Licitra et al. (1996). The NFC content (% of DM) of the diets was calculated (NRC, 2001) as:
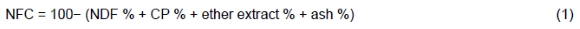
The intake of N (g/d) was calculated by multiplying DMI by the N content of the diet. Predicted urine N, faecal N excretion, and N efficiency were calculated (Kohn et al., 2002) using the following equations:
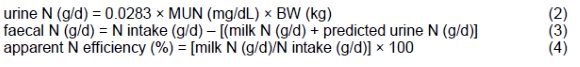
One faecal grab sample per cow was taken from the rectum daily at 4 h after the morning feeding during the last 5 d of each period and frozen at -20 °C until analysed. Frozen samples were thawed at room temperature, dried at 60 °C for 72 h, ground, and pooled (by cow within the period). Acid detergent insoluble ash was used as an internal marker to determine the apparent total-tract nutrient digestibility (Van Keulen & Young, 1977; Kargar et al., 2012).
Cows were milked three times daily at 01:00, 09:00, and 17:00. On days 16 to 21 of each period, milk yield was recorded and sampled at each milking event. Milk samples were analysed to determine fat, true protein, lactose, and SNF contents using the Milkoscan (134 BN, Foss Electric, Hillerod, Denmark; AOAC International, 2002). The MUN content was also determined using an enzymatic assay (Wilson et al., 1998). The yield of 4% FCM (kg/d) and ECM (kg/d) was calculated (NRC, 2001) using the following equations:
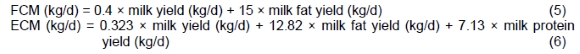
Feed efficiency was calculated by dividing the daily actual milk or 4% FCM production (kg/d) by DMI (kg/d). Income over feed cost (IOFC) was defined as the portion of income from milk sold that remained after paying for purchased and farm-raised feed used to produce the milk. Sorting index was calculated as described by Leonardi and Armentano (2003).
Blood samples were collected 4 h after the morning feeding from the coccygeal artery using evacuated tubes with anticoagulant (EDTA) on d 20 of each period and placed immediately on ice. Samples were centrifuged at 3,000 × g for 15 min to separate plasma samples, which were then stored at -20 °C until analysis (Savari et al., 2018). Concentrations of glucose, albumin, total protein, plasma urea N, cholesterol, and triglyceride were measured using commercial kits (Pars Azmoon Co., Tehran, Iran) according to the manufacturer's instructions. Globulin concentrations were calculated by deducting albumin from total protein.
On d 21 of each period, rumen contents were sampled via oesophageal tubing 4 h after the morning feeding. The pH of ruminal fluid was determined immediately before 10 mL of the sample was preserved with 2 mL of a 25% meta-phosphoric acid solution (Savari et al., 2018) and frozen at -20 °C until analyses for determination of NH3-N. Rumen fluid samples for NH3 analysis were thawed and centrifuged at 30,000 × g for 20 min at 4 °C; the supernatant was removed and analysed for NH3 using the colorimetric phenol-hypochlorite method (Broderick & Kang, 1980).
Data were analysed using the PROC mixed procedure in SAS (version 9.0; SAS Institute Inc., 2002) for a replicated 4 x 4 Latin square design. The following model was used:

where Yijki = each observation,
μ = overall mean,
Sl = fixed effect of square,
P(S)il = fixed effect of period within square,
A(S)ji= random effect of cow within square,
Tk = fixed effect of treatment,
and eijki = random error.
Significance was declared at P <0.05 and trends at 0.05 <P <0.10.
Results and Discussion
Chemical composition of T-SBM, FM, MM, and FMM are presented in Table 1. CP content for FM was higher than that of the other sources, although diets were formulated to be both isonitrogenous and isoenergetic. The ingredient and chemical compositions of experimental diets are given in Table 2. All treatments had the same baseline with different RUP sources being used in the experimental diets. Crude protein was kept the same for all the treatments (equal to 16.7%). As anticipated, T-SBM heat processing extensively decreased the proportions of A, B1, and B2 fractions, but increased proportions of B3 and C fractions compared to SBM (Table 1). This resulted in an increase in RUP content of treated SB in comparison with SBM. Our previous experiment (Savari et al., 2018) recommended the use of a lower RUP ratio (35% of CP) and the use of ground corn to obtain a better production efficiency and reduce the dietary cost. Therefore, in this experiment, we used a 35% RUP level (RDP: RUP ratio of 65:35 based on CP) as well as ground corn in the diets.
Dry matter intake was not different across experimental diets (Table 3). In this study, feed intake (24.2 kg/d) was not affected by changes in the source of dietary RUP (P = 0.39). Experimental diets were found to have no effect on the production of actual or fat and energy corrected milk (Table 3). Composition and yields of milk fat and protein were not affected by the diets (Table 3). Although in this experiment, it was expected that milk fat percentage would be reduced substantially by fish meal, due to the use of the low levels of fish meal (2.5% DM) containing 10-15% fish oil, the negative effects on performance and milk composition were decreased. The percentage of milk fat was still numerically lower in fish meal treatments (2.98 vs. 3.04 %) than in other treatments.
Feed intake was not affected by changes in the source of dietary RUP. Our results are in line with an extensive literature review performed by Santos et al. (1998), who reported no significant differences in DMI when different high-RUP feeds were used in dairy rations. In different research, feeding RUP products vs. SBM resulted in varied DMI. Several studies have reported an increase in DMI for cows fed treated SBM products and a decrease in DMI for cows fed animal RUP products (Ipharraguerre & Clark, 2005). One study showed that replacing 2% soybean meal with fish meal in mid-lactation diets caused a slight increase in dry matter intake, but 4% fish meal replacement reduced the DMI (Schwab et al., 1992). In the current experiment, levels of less than 4% of the RUP source were used and the results of this study are consistent with the previous experiment. The lack of effect of the treatment on DMI was likely because of the no changes in CP, NEL, NDF among diets, or physiological status of animals (NRC, 2001).
Production of actual or fat- and energy-corrected milk were not affected by experimental diets. This agrees with the results of the study performed by Santos et al. (1998), who reported no marked differences in milk yield for cows fed different high-RUP feed (Santos et al., 1998). Conversely, Ipharraguerre and Clark (2005) suggested that substituting soybean meal with RUP sources reduced microbial protein production, with the exception of treated soybean sources that even increased microbial protein production compared to soybean meal. Similar milk yields across the treatments in the current study (Table 3) were likely because of the isoenergetic diets and same DMI across treatments.
In this study, composition and yield of milk fat and protein were not affected by the experimental diets. Palmquist et al. (2005) reported that fish oil, due to its high concentration of unsaturated fatty acids, can effectively reduce feed intake and milk fat. An increase in unsaturated fatty acids from fish meal will result in an increase in vaccenic acid, mainly related to incomplete ruminal biohydrogenation of linolenic acid. Inhibition of end-stage biohydrogenation of PUFAs by feeding fish oil may be related to the inhibition of the growth of group B bacteria or enzymes involved in vaccenic acid reduction (Bauman et al., 2000). An increase of trans C18:1 concentration due to the inhibition of the last stage of ruminal biohydrogenation has been reported to be the reason for reduced milk fat when feeding fish oil (Alizadeh et al., 2012).
Although in this experiment, it was expected that the milk fat percentage would be markedly reduced with fish meal, due to use of the low levels of fish meal (2.5% DM) containing 10-15% fish oil, the negative effects on performance and milk composition were decreased, but the percentage of milk fat was still numerically lower in fish meal treatments (2.98 vs. 3.04 %) than in other treatments. Limiting amino acids (methionine and lysine) play an important role in milk protein synthesis. Fish meal has a higher percentage of methionine and lysine than meat and soybean meal, and the increase in milk protein content due to fish meal is probably related to the improved balance of amino acids reached to the intestine (NRC, 2001). No differences were observed in milk fat, protein, and lactose yield and percentages among diets, which may be related to no substantial changes in ruminal fluid pH and fermentation characteristics (Table 5). Due to the lack of a statistically significant effect of experimental treatments on feed intake and milk yield in this experiment, feed efficiency was expected to be similar.
In this study, the mean digestibility of dry matter, organic matter, and protein of different diets was 67.4%, 68.2%, and 67.2%, respectively, and there was no substantial difference between the diets for the above criteria (Table 4). There was no substantial difference in mean apparent digestibility between the treatments (Table 3). In this study, cows consuming different treatments had the same nitrogen efficiency (P = 0.8).
In ruminants, there is a correlation between digestibility and DMI. Increasing digestibility up to 67% results in more feed intake (Van Soest, 1973). Our results showed that there was no substantial difference between the diets in the digestibility of dry matter, organic matter, and protein. Nutrient digestibility in different treatments could also not have had an effect on their DMI. As the milk production and feed intake were similar between the treatments, the feed efficiency was not affected by the treatments (Table 3).
Many studies have suggested that replacing fish meal with soybean meal, heated soybean meal, or corn gluten meal did not have a marked effect on nutrient digestibility (Kohn et ai., 2002; Santos et al., 1998). Replacing soybean meal with fish meal had no marked effect on the apparent digestibility of organic matter; McCarthy et ai. (1989) also reported that the type of protein source used in the diet (fish meal or soybean meal) had no marked effect on the apparent digestibility of organic matter in the rumen, post-rumen, and whole digestive tract (McCarthy et al., 1989). In the present study, there was no statistical difference between mean apparent total tract digestibility of protein between the experimental treatments.
According to Keery et ai. (1993), the use of different protein sources (fish meal, soybean meal or heated soybean meal) in the diet had no marked effect on digestibility of the protein in the digestive tract. They reported an average apparent digestibility of protein in the digestive tract of 67%, which is consistent with the current experiment. Klusmeyer et ai. (1991) also reported that using fish meal instead of soybean meal in the diet did not have a marked effect on the apparent digestibility of crude protein. They estimated the apparent total tract digestibility of crude protein in different diets to be 67-70%. Improving protein efficiency means less wastage of this valuable resource in the animal's body, which can be seen in the effects on protein excretion in the urine and faeces of animals, as well as in other excretions such as milk urea nitrogen. By wasting more and reducing protein efficiency in the body, most of the protein breaks down in the blood and converts to metabolites such as blood urea nitrogen, which is excreted through the body's excretory fluids, especially milk and urine (Schwab, 1994).
Due to similar feed intake, and thus equal nitrogen consumption, between experimental treatments receiving different sources of RUP; the same digestibility of nitrogen in the cows consuming these sources; and the production of identical milk in the cows, there was no statistical difference in the apparent nitrogen efficiency of the cows fed the experimental treatments (Table 4).
There was no significant difference in rumen pH between treatments (Table 5). In this study, there was no significant difference in ruminal ammonia nitrogen levels between the treatments with respect to the same dietary protein level and similar MUN concentration. No significant differences were observed among the treatments regarding the pH of the faeces and urine (Table 5). The RUP source did not affect feed sorting index (Table 5). Blood metabolites were not affected by the RUP source, which is in agreement with previous studies (Santos et al., 1998). According to McCarthy et al. (1989), substitution of RUP source for soybean meal had no significant effect on rumen pH. In this study, there was no significant difference in ruminal ammonia nitrogen levels between the treatments with respect to the same dietary protein level and similar MUN concentration.
Cows typically opt for small particles of feed due to their higher nutritional value than longer feed; in all diets, cows sorted against 19-mm sieve residue (Table 5). Lactating cows can also choose mainly long particles of feed to meet their requirements for physically effective NDF when rumen pH is low due to low effective NDF intake. These differences are due to the size of the feed particles and possibly indicate the need of the animal for nutrients (Beauchemin & Yang, 2005).
In this experiment, treated soybean meal, fish meal, meat meal, and an equal mixture of fish meal and meat meal were used as sources of RUP. Due to the price difference of these protein sources, the prices of diets contained meat meal were lower than those of other diets. Because of the lower milk fat in cows fed fish meal, the price of milk in these cows was lower than in cows consuming other treatments. Income from milk sales for cows consuming meat meal tended to increase compared to other treatments. The results of this study showed that based on the amount of obtained profit and also considering the significant difference between the ratio of milk sales to ration cost (IOFC, P = 0.04, Table 5) between treatments and increasing this parameter in the treatments containing meat meal, the use of meat meal as a source of RUP for feed to high yield cows under our experimental conditions can be recommended.
Conclusions
In this study, the effect of dietary RUP source on the performance of high-yielding cows was investigated. Dry matter intake and milk production were not statistically different in cows fed different sources of RUP. Due to similar feed intakes and nitrogen consumption, as well as CP apparent digestibility among the different sources of RUP, apparent nitrogen efficiency was not affected by the treatments. Based on the income over feed cost (IOFC) analysis, use of meat meal as a source of dietary RUP can be suggested for high yielding dairy cows. Finally, the limitations in using meat meal as a ruminant feed should be noted.
Acknowledgements
The authors acknowledge the Iran National Science Foundation (no. 95825440, Tehran Province, Tehran, Iran) and Isfahan University of Technology (Isfahan, Iran) for their financial support of this study. They also express their appreciation to M. Mirzaei (University of Arak) and M. Bahrami-Yekdangi (Isfahan University of Technology, Isfahan, Iran) for their input on the research and laboratory analyses.
Authors' Contributions
MS (ORCID 0000-0002-1773-5287), FA, and MSSF participated in designing the study, laboratory analysis, and manuscript writing. MKH was involved in drafting and revising the manuscript for intellectual content. All authors review and approved the manuscript before submitted for publication.
Conflict of Interest Declaration
The authors declare that they have no conflict of interest.
References
Alizadeh, A., Alikhani, M., Ghorbani, G.R., Rahmani, H., Rashidi, L. & Loor, J., 2012. Effects of feeding roasted safflower seeds (variety IL-111) and fish oil on dry matter intake, performance, and milk fatty acid profiles in dairy cattle. J. Anim. Physiol. Anim. Nutr. 96, 466-473. [ Links ]
Alvarado, C.G., Anrique, R.G., Navarrete, S.Q., 2009. Effect of including extruded, rolled or ground corn in dairy cow diets based on direct cut grass silage. Chil. J. Agr. Res. 69, 356-365. [ Links ]
AOAC International, 2002. Official Methods of Analysis. 17th ed. AOAC International, Arlington, VA. [ Links ]
Abdollahzadeh, F., Ahmadi, F., Khani, M., Mirzaei, M., 2020. Poultry by-product meal as a replacement to xylose-treated soybean meal in diet of early- to mid-lactation Holstein cows. Trop Anim Health Prod. 53(1):38. [ Links ]
Bauman, D., Baumgard, L., Corl, B., Griinari, D.J., 2000. Biosynthesis of conjugated linoleic acid in ruminants. J. Anim. Sci. 77, 1-15. [ Links ]
Beauchemin, K., Yang, W., 2005. Effects of physically effective fiber on intake, chewing activity, and ruminal acidosis for dairy cows fed diets based on corn silage. J. Dairy Sci. 88: 2117-2129. [ Links ]
Binh Truong, N., Thanh Trung, T., 2023. Effect of dietary crude protein levels on feed intake and nutrient digestibility of Wagyu crossbred cattle. Online J. Anim. Feed Res, 13(4): 295-301. [ Links ]
Broderick, G.A., 2006. Improving nitrogen utilization in the rumen of the lactating dairy cow. Florida Ruminant Nutr. Symp., February, 2006. [ Links ]
Broderick, G., Kang, J., 1980. Automated simultaneous determination of ammonia and total amino acids in ruminai fluid and in vitro media. J. Dairy Sci. 63: 64-75. [ Links ]
DePeters, E., Getachew, G., Fadel, J., Zinn, R., Taylor, S., Pareas, J., Hinders, R., Aseltine, M., 2003. In vitro gas production as a method to compare fermentation characteristics of steam-flaked corn. Anim. Feed Sci. Technol. 105: 109-122. [ Links ]
Fadel, M., EL-Garhi, M.S., Ghattas, T., Am, M., 2017. Effect of feeding different crude protein diets on reproductive performance of Holstein dairy cow. Assiut Vet. Med. J. 63: 128-135. [ Links ]
Fiems, L., Cottyn, B., Boucque, C.V., Vanacker, J., Buysse, F., 1990. Effect of grain processing on in sacco digestibility and degradability in the rumen. Archives of Anim. Nutr. 40: 713-721. [ Links ]
Huntington, G.B., 1997. Starch utilization by ruminants: From basics to the bunk. J. Anim. Sci. 75: 852-867. [ Links ]
Hussein, H.S., Jordan, R.M., 1991. Fish meal as a protein supplement in ruminant diets: A review. J. Anim. Sci. 69: 2147-56. [ Links ]
Ipharraguerre, I.R., Clark, J.H., 2005. Impacts of the source and amount of crude protein on the intestinal supply of nitrogen fractions and performance of dairy cows. J. Dairy Sci. 88: 22-37. [ Links ]
Jones, B., Mohamed, O., Prange, R., Satter, L., 1988. Degradation of methionine hydroxy analog in the rumen of lactating cows. J. Dairy Sci. 71: 525-529. [ Links ]
Kargar, S., Ghorbani, G.R., Alikhani, M., Khorvash, M., Rashidi, L., Schingoethe, D.J., 2012. Lactational performance and milk fatty acid profile of Holstein cows in response to dietary fat supplements and forage:concentrate ratio. Livest. Sci. 150: 274-283. [ Links ]
Keery, C., Amos, H., Froetschel, M., 1993. Effects of supplemental protein source on intraruminal fermentation, protein degradation, and amino acid absorption. J. Dairy Sci. 76: 514-524. [ Links ]
Klusmeyer, I., Lynch, G., Clark, J., Nelson, D., 1991. Effects of calcium salts of fatty acids and protein source on ruminal fermentation and nutrient flow to duodenum of cows. J. Dairy Sci. 74: 2206-2219. [ Links ]
Kohn, R., Kalscheur, K., Russek-Cohen, E., 2002. Evaluation of models to estimate urinary nitrogen and expected milk urea nitrogen. J. Dairy Sci. 85: 227-233. [ Links ]
Kononoff, P., Heinrichs, A., 2003. The effect of reducing alfalfa haylage particle size on cows in early lactation. J. Dairy Sci. 86: 1445-1457. [ Links ]
Leonardi, C., Armentano, L., 2003. Effect of quantity, quality, and length of alfalfa hay on selective consumption by dairy cows. J. Dairy Sci. 86: 557-564. [ Links ]
Licitra, G., Hernandez, T., Van Soest, P., 1996. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. Technol. 57: 347-358. [ Links ]
McCarthy, R., Klusmeyer, T., Vicini, J., Clark, J., Nelson, D., 1989. Effects of source of protein and carbohydrate on ruminal fermentation and passage of nutrients to the small intestine of lactating cows. J. Dairy Sci. 72: 2002-2016. [ Links ]
Mielke, C.D., Schingoethe, D.J., 1981. Heat-treated soybeans for lactating cows. J. Dairy Sci. 64: 1579-1586. [ Links ]
National Research Council (NRC), 2001. Nutrient Requirements of Dairy Cattle. 7th rev. ed. Natl. Acad. Sci., Washington, DC, USA. [ Links ]
Nikkhah, A., Alikhani, M., Amanlou, H., 2004. Effects of feeding ground or steam-flaked broom sorghum and ground barley on performance of dairy cows in mid-lactation. J. Dairy Sci. 87: 122-130. [ Links ]
Palmquist, D.L., Lock, A.L., Shingfield, K.J., Bauman, D.E., 2005. Biosynthesis of conjugated linoleic acid in ruminants and humans. Adv. Food Nutr. Res. 50: 179-217. [ Links ]
Rafiee-Yarandi, H., Ghorbani, G.R., Alikhani, M., Sadeghi-Sefidmazgi, A., Drackley, J., 2016. A comparison of the effect of soybeans roasted at different temperatures versus calcium salts of fatty acids on performance and milk fatty acid composition of mid-lactation Holstein cows. J. Dairy Sci. 99: 5422-5435. [ Links ]
Rajala-Schultz, P., Saville, W., 2003. Sources of variation in milk urea nitrogen in Ohio dairy herds. J. Dairy Sci. 86: 1653-1661. [ Links ]
Santos, F., Santos, J., Theurer, C., Huber, J., 1998. Effects of rumen-undegradable protein on dairy cow performance: A 12-year literature review. J. Dairy Sci. 81: 3182-3213. [ Links ]
Santos, J., Huber, J., Theurer, C., Nussio, L., Tarazon, M., Santos, F., 1999. Response of lactating dairy cows to steam-flaked sorghum, steam-flaked corn, or steam-rolled corn and protein sources of differing degradability. J. Dairy Sci. 82: 728-737. [ Links ]
Savari, M., Khorvash, M., Amanlou, H., Ghorbani, G.R., Ghasemi, E., Mirzaei, M., 2018. Effects of rumen-degradable protein: rumen-undegradable protein ratio and corn processing on production performance, nitrogen efficiency, and feeding behavior of Holstein dairy cows. J. Dairy Sci. 101: 1-12. [ Links ]
Schwab, C., 1994. Optimizing amino acid nutrition for optimum yields of milk and milk protein. Southwest Nutrition Management: 114-129. [ Links ]
Schwab, C., Bozak, C., Whitehouse, N., Mesbah, M., 1992. Amino acid limitation and flow to duodenum at four stages of lactation. 1. Sequence of lysine and methionine limitation. J. Dairy Sci. 75: 3486-3502. [ Links ]
Schwab, C., Boucher, S., Sloan, B., 2007. Metabolizable protein and amino acid nutrition of the cow: Where are we in 2007? Proceedings of the 68th annual Minnesota Nutrition Conference. pp 121-138. [ Links ]
Sniffen, C.J., O'Connor, J.D., Soest, P.J.V., Fox, F.D.G., Russell, J.B., 1992. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 70: 3562-3577. [ Links ]
Van Keulen, J. Young, B., 1977. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci. 44: 282-287. [ Links ]
Van Soest, P., 1973. Collaborative study of acid-detergent fiber and lignin. J. Assoc. Office. Anal. Chem. 23: 455-464. [ Links ]
Van Soest, P.J., Robertson, J.B., Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74: 3583-3597. [ Links ]
Wilson, R., Overton, T., Clark, J., 1998. Effects of Yucca shidigera extract and soluble protein on performance of cows and concentrations of urea nitrogen in plasma and milk. J. Dairy Sci. 81: 1022-1027. [ Links ]
Yu, P., Huber, J., Santos, F., Simas, J., Theurer, C., 1998. Effects of ground, steam-flaked, and steam-rolled corn grains on performance of lactating cows. J. Dairy Sci. 81: 777-783. [ Links ]
Zhong, R., Li, J., Gao, Y., Tan, Z., Ren, G., 2008. Effects of substitution of different levels of steam-flaked corn for finely ground corn on lactation and digestion in early lactation dairy cows. J. Dairy Sci. 91: 3931 -3937. [ Links ]
Submitted 18 February 2022
Accepted 20 October 2022
Published 17 November 2023
# Corresponding author: Email: m.s.safaee@ardakan.ac.ir


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}