










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 no.5 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i5.12
Genomic markers associated with antibody response to Newcastle disease virus of Sasso chickens raised in Ethiopia
M. GirmaI, II, #; K.M. MorrisIII; K. SuttonIII; W. EsatuII; B. SolomonII; T. Dessie A. PsifidiII; L. VerveldeIII; O. HanotteII, III, V; G. BanosIII, VI; D.P. MalatjiI
IDepartment of Agriculture and Animal Health, College of Agriculture and Environmental Sciences, University of South Africa, Florida 1710, South Africa
IICTLGH - LiveGene, International Livestock Research Institute (ILRI), P.O. Box 5689, Addis Ababa, Ethiopia
IIIThe Roslin Institute, University of Edinburgh, Easter Bush Campus, Midlothian, EH25 9RG, UK
IVThe Royal Veterinary College, Hawkshead Lane, Hatfield, Hertfordshire, AL9 7TA, UK
VCells, Organisms, and Molecular Genetics, School of Life Sciences, University of Nottingham, Nottingham, NG7 2RD, UK
VIScotland's Rural College (SRUC), Animal and Veterinary Sciences, Easter Bush, EH25 9RG, Midlothian, UK
ABSTRACT
Newcastle disease virus (NDV) is one of the highly contagious avian pathogens that threatens poultry producers in endemic zones due to its epidemic potential. Selection for antibody (Ab) response can effectively improve disease resistance in chickens. However, the molecular basis of the variation in Ab response to NDV is unclear. This study aimed to detect genomic markers and genes modulating Ab response to NDV in chickens reared under tropical, outdoor conditions. A genome-wide association study (GWAS) was conducted on Sasso T451A chickens that were naturally exposed to infectious diseases from 56 to 112 days of age to identify regions associated with Ab response to NDV. Phenotypic immune data from 935 chickens, monitored in two batches, and genotyping data of these chickens based low-pass sequencing (2,676,181 single nucleotide polymorphisms, SNPs) were used. BioMart data mining and variant effect predictor tools were used to annotate SNPs and candidate genes, respectively. A total of five SNPs (rs316795557 (FOXP2), chr 1; rs313761644 (CEP170B), chr 5; rs733628728, chr 13; and two unnamed SNPs, chr 30 and chr 33) were associated with the chicken antibody response to NDV at the suggestive significance level. These SNPs are located on chromosomes 1, 5, and 13 and are in genomic regions that contain several genes with roles in the regulation of the immune response. The results of this study pave the path for more investigation into the host immune response of chickens to NDV.
Keywords: antibody response, genome-wide association analysis, Newcastle disease, Sasso T451A, vaccine challenge
Introduction
The poultry industry is a very important factor in the world economy in terms of food safety and nutrition, and is the fastest-growing agricultural sub-sector, especially in developing countries. In low-income/developing countries, chickens play a vital role in daily life (Malatji et al., 2016). Poultry eggs and meat deliver high-quality sources of protein and micro- and macronutrients. Consumption of poultry meat and eggs and animal-source foods has increased rapidly over decades (FAO, 2020).
As in many developing countries, chickens are widely kept in Ethiopia (Hassen et al., 2006) where chicken meat contributes to approximately 5% of the total national meat production (EMDIDI, 2017). Relative to other livestock animals, cattle, sheep, and goats, chicken meat production remains relatively small. Poultry rearing provides a major income-generating activity for households in which women and children are the major beneficiaries, which is often not the case with larger livestock (Guèye, 2000). Chickens serve as a living savings bank and require very little investment, thereby offering rare opportunities for investment and security against risk. Chickens largely dominate flock composition and make up ~98% of the total poultry (chickens, ducks, and turkeys) population kept in Africa (Hassen et al., 2006).
Although the global production of chickens has experienced massive changes and growth over the past 50 years, the poultry industry is still threatened by various diseases (Jie & Liu, 2011), especially Newcastle disease virus (NDV). Poultry disease is believed to be the main constraint that frustrates investment in this scavenging and semi-scavenging system by rural households. Thus, the potential of free-range chicken production has not fully been exploited. If any success is to be achieved in the improvement of free-ranging chicken production in Africa, it will inevitably depend on the successful control of major poultry diseases, in general, and Newcastle disease (ND), in particular (Mekonnen et al., 2013).
The immune response associated with a specific disease may be a useful indicator of general disease resistance. The immune response and resistance to infectious diseases are difficult traits to measure and, thereby, to improve. Immunological characteristics such as antibody titres are heritable in poultry (Lamont et al., 2003), indicating the possibility of discovering loci or genes related to immune or disease-resistance traits. A common method to detect genetic markers (e.g., single nucleotide polymorphisms, SNPs) associated with phenotypic traits is the use of the DNA Beadchip technology, allowing simultaneous genotyping of thousands of genetic markers. This approach is commonly used for human disease and economically important animal trait research. In chickens, many genome-wide association studies (GWAS) have been conducted using the Illumina 60K SNP Beadchip or the 600K SNP Affymetrix array. Using this method, major loci associated with diseases (Raeesi et al., 2017), growth (Guo et al., 2017; Pertille et al., 2017), egg production (Wolc et al., 2012), carcass (Huang et al., 2018) and meat quantitative traits (Moreira et al., 2018), resistance to Marek's disease (Li et al., 2013), and immune responses to NDV (Luo et al., 2013) have been identified. Previous work has demonstrated the utility of this approach for the genomic study of health and immune traits in African chickens (Psifidi et al., 2016; Banos et al., 2020).
The aim of the present study was to identify candidate genes and genomic regions controlling antibody response to NDV of Sasso T451A chickens raised in Ethiopia.
Materials and Methods
All animal protocols were conducted as per the revised Animals (Scientific Procedures) Act 1986 procedures and approved by the Institutional Animal Care and Use Committee of the International Livestock Research Institute (Approval no: ILRI-IACUC2020-10).
A total of 1,022 day-old Sasso T451A dual-purpose chickens were acquired from EthioChicken in Ethiopia. The chickens were maintained at the International Livestock Research Institute (ILRI) poultry research facility in two batches [autumn (September to November) or winter (December to February)] and consisted of 507 (194 males and 313 females) and 515 (209 males, and 306 females) chickens, respectively. The chickens were maintained in a deep litter house during the brooder phase for 56 days from hatching. The chickens had intact beaks and received routine vaccinations. The chickens were immunized with the commercial NDV live vaccine of the Hichner B1/LaSota strain (National Veterinary Institute (NVI), Ethiopia), using the standard dose given in the instructions of the vaccine, by ocular eye drop at 7 and 21 days of age (NVI, 2016). In Ethiopia, Sasso chickens are distributed to farmers at days 56 (after they have been fully vaccinated). Wing tags facilitated the identification of individuals, which were sampled for genotyping and enzyme-linked immunosorbent assays (ELISAs) before being released into the paddock. The chickens were subsequently raised in outdoor, semi-scavenging conditions (fed mainly from scavenging with some supplementation of commercial feed) from 56 days of age until they reached an average market weight of approximately 1,500 g, when chickens were ~112 days old. Chickens were naturally exposed to infectious diseases agents while monitored outdoors.
Blood was retrieved from the wing vein at 56 and 112 days of age for 1022 and 795 chickens, respectively. The blood was allowed to coagulate overnight at room temperature before the removal of the serum. Serum samples were stored at -20 °C until use. Serum NDV-specific antibodies (Ab) were measured in serum collected at 56 and 112 days of age using the IDEXX NDV ELISA kit (IDEXX Laboratories, Westbrook, ME, USA, Cat#99-09263). All samples were tested at a 1:100 serum dilution in duplicate with a positive and a negative control being added to each plate.
Genomic DNA was extracted from blood preserved on QIAcard FTA Elute Micro cards (QIAGEN, Cat. No. WB120410) from all chickens before exposure to the natural environment and shipped to GeneSeek, Neogen Genomics (Lincoln, NE, United States). The DNA from a total of 963 chickens was genotyped using low-pass sequencing and imputed using the Gencove platform. GATK software was used for SNP calling with the default parameters and Indels were sequenced and/or imputed, but they were not considered in the study (Mckenna et al., 2010). Finally, 14,123,057 SNPs remained for further edits.
Before statistical analysis, evaluation of the data quality and pre-processing of the data was performed to avoid the introduction of bias into the analysis (Anderson et al., 2010). Raw SNP data were first subjected to quality control and loci with unknown chromosomal locations were removed. The SNP data quality control was carried out using PLINK v1.9 (Purcell et al., 2007), and samples with a high genotyping missing rate (>90%) were excluded. SNPs were removed if they failed to pass the following criteria: call rate >90%; minor allele frequency (MAF) >0.01 and P-value for Hardy-Weinberg equilibrium test >1×10-6. After a series of quality checks, 935 samples and 11,272,670 SNPs remained for the ensuing data analysis. Considering that clusters of SNPs in high linkage disequilibrium (LD) may bias the principal component analysis (PCA) results (Wang et al., 2009), all autosomal SNPs were pruned for LD using pair-wise genotype correction (r2 >0.8) in 100 SNP sliding widows with a step of 10 SNPs across the genome; 2,676,181 SNPs remained for analysis.
Population structure and relatedness are major sources of confounding effects in genetic association studies (Astle & Balding, 2009). To assess the population structure, PCA was performed and implemented using PLINK v1.9. PCA was performed using an identity-by-state matrix based on SNP genotypes to estimate the genetic relatedness between individuals and investigate the presence of population stratification between batches, using the EIGENSOFT v6.1.4 software before the GWAS (Price et al., 2006).
The GWAS analyses were then performed separately for each immune trait (Ab response at age 56 and 112 days) for each batch using the following linear mixed model:

where y is the vector of trait values (Ab at age 56 and 112 days) for all individuals; W is a matrix of covariates (fixed effects that contains the batch, age, season, sex, growth traits, and the top three PCs); a is a vector of the corresponding coefficients including the intercept; x is a vector of genotypes of a marker; p is the effect size of the marker; u is a vector of random effects with a covariance structure as u ~ N(0, Gσ2 u), where G is the genetic relatedness matrix calculated from all SNP markers, and σ2 u is the polygenic additive variance; ε is a vector of residual errors with ε ~ N(0, Iσ2 ε), where I is the identity matrix and σ2 ε is the residual variance.
The significance threshold for the GWAS was adjusted for multiple testing using the Bonferroni correction method (Benjamini & Hochberg, 1995). We set the following two significance thresholds for our data: P <0.05/N for a 5% genome-wide significance and P <1/N for a suggestive significance level, where N is the number of SNPs remaining after pruning for LD. To visualize the result from the GWAS, Manhattan plots and quantile-quantile plots were created using the qqman package in R v4.1.0 for each trait. The genomic inflation factor (A) was calculated using the qqman package in R with the median option (Turner, 2015).
The Variable Effect Predictor (VEP) tool in the Ensembl database and the Galgal6 assembly were used to map all significant SNPs identified in the GWAS to the reference genome and to annotate them, respectively (http://www.ensembl.org/Tools/VEP). Additionally, using the Galgal6 assembly and the BioMart data mining tool, genes that were situated 100 kb upstream and downstream of the significant SNPs were annotated (http://www.ensembl.org/biomart/martview/). A list of all the genes that were close to the significantly associated SNPs for Ab response to NDV was then compiled.
Genetic parameters were estimated for Ab response to NDV using the model (Equation 1). Estimates of the obtained variance components were determined using the restricted maximum likelihood analysis option in GCTA v1.24.7. Bivariate analyses were also conducted with the same model to estimate immune phenotypic and genetic correlations among the studied traits. All the above analyses were performed separately for each age using the GCTA tool (Yang et al., 2011).
Results and Discussion
Genome-wide association study is a tool used for genetic analysis of traits of economical important in domestic animals. In this study, GWAS was employed to investigate the genetic variation in the immune response to NDV in Sasso T451A. Host immune reaction to viruses is a complex process (Fischer et al., 2013) and immune response to NDV can be observed as a quantitative trait under polygenic regulation (Luo et al., 2013; Saelao et al., 2019).
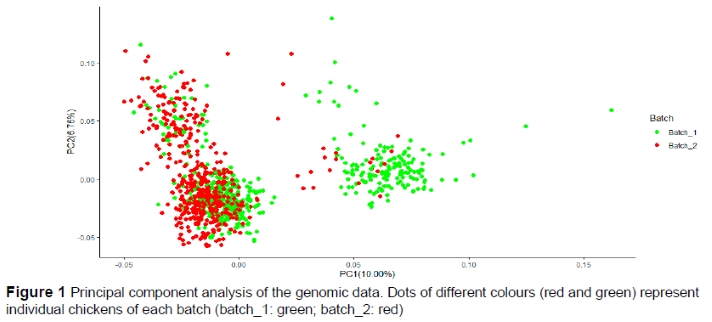
The PCA analysis did not reveal any clear genetic separation between the two batches (Figure 1). PCA results showed some structure in the genomic data that was not entirely attributed to batch. The first PCA component explained 10.0%, and the second principal component, 6.7%, of the total variance. Therefore, 2-3 principal components (PCs) were fitted in the model for statistical analysis in order to account for structure. GRM, that was fitted in the model, also accounted for structure.
The PCA results (Figure 1) support that the two batches had the same genetic origins. Furthermore, PCA results were compatible with a three-way crossing origin for the chicken of the two batches. Compared to two-way crossbred chickens, three-way crossbred chickens have lower mortality and better production traits, such as age at sexual maturity, annual egg production, feed intake, and feed conversion (Khawaja et al., 2013).
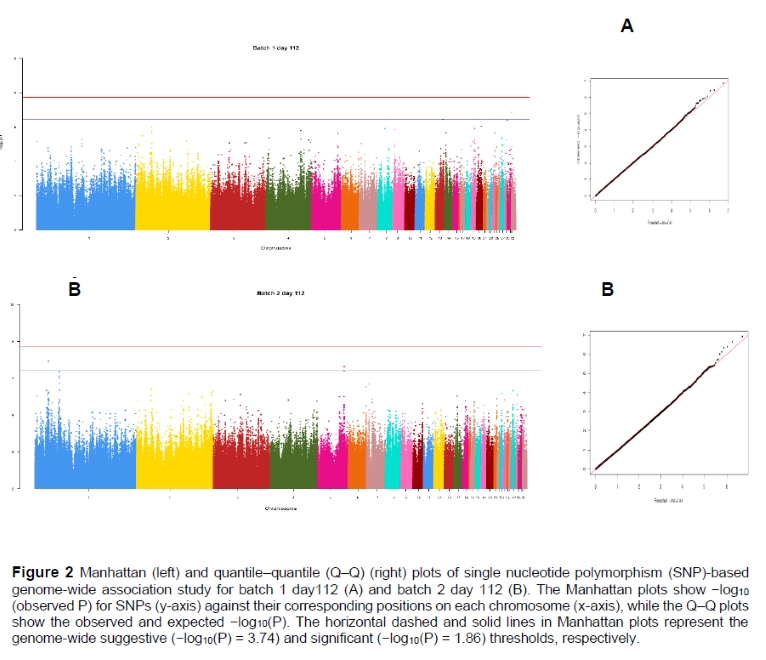
The univariate GWAS was performed to find genomic markers associated with the NDV antibody response of Sasso T451A at 56 and 112 days of age in each of the batches. GWAS results are presented as Manhattan and QQ plots (Figure 2). In the present study, genomic control inflation factors were very close to unity (λ = 1.02 and 1.00 for each analysis, respectively), which reflected low population stratification.
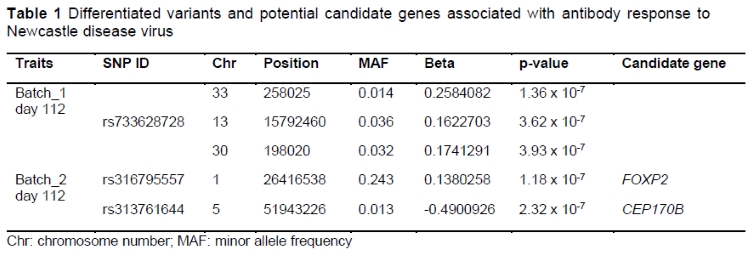
A total of five SNPs were identified at the suggestive level threshold for antibody response to NDV at day 112, of which two of them were novel SNPs. These SNPs were located in the position at 15792460 bp (rs733628728 chromosome 13, P = 3.62 x 10-7), 198020 bp (unnamed SNPs, chromosome 30, P = 3.93 x 107), and 258025 bp (unnamed SNPs, chromosome 33, P = 1.36 x 107) (batch one, day 112). For batch two at day 112 and two SNPs with the suggestive association, SNPs rs316795557 (P = 1.18 x 107) at position 26416538 bp on chromosome 1 and rs313761644 (P = 2.32 x 10-7) at position 51943226 bp, on chromosome 5, were identified. No significant P-values were obtained for the SNP effect of antibody response to NDV at day 56 for both batches (Table 1; Figure 2).
Previous studies have identified different genomic regions associated with antibody response to NDV (Yonash et al., 2001; Biscarini et al., 2010; Wang et al., 2015). According to Yonash et al. (2001), the quantitative trait loci QTLs for the Ab response to NDV in broiler chickens were found on chromosomes 2 and 18. Biscarini et al. (2010) found 13 QTLs located on chromosomes 3, 4, 5, 9, 13, 16, and 22, and Z, associated with the Ab response to NDV. Wang et al. (2015) reported six QTLs modulating Ab response to NDV on chromosomes 2, 4, and Z. Jacob et al. (2000) found a QTL on chromosome 16 associated with the major histocompatibility complex (MHC) of NDV (Zhou & Lamont, 2003; Zhang et al., 2015). Several reasons might explain these different results, including different doses of ND vaccine, time post-vaccination, the genotyping method used, choice of statistical models, and the genetic composition of the experimental populations (McElroy et al., 2006; Ioannidis et al., 2009; Saelao et al., 2019). Moreover, our study used the Bonferroni correction method for multiple testing and the significance thresholds used here might have been more stringent than the traditional cutoffs commonly used in studies aiming to identify candidate regions for functional associations between SNPs and immune phenotypes. Additionally, several studies (Yonash et al., 2001; Biscarini et al., 2010) have focused on broiler and layer populations, while in this study, we employed the Sasso T451A, a dual-purpose chicken.
While the present study used GWAS and SNPs, most earlier investigations used microsatellite markers. Furthermore, different types of Ab detection methods may be associated with different QTLs (Saelao et al., 2019). Earlier research concentrated on the primary Ab response to NDV, but here we examined the secondary Ab response to NDV. In the primary Ab response, the main class of Ab produced is immunoglobulin M (IgM), whereas, in the secondary Ab response, it is IgY (Sarker et al., 1999); thus, chicken's ranking on Ab response to the first immunization may differ from that to the second, resulting in the identification of separate QTLs (Biscarini et al., 2010). The current study revealed suggestive QTLs for Ab response to NDV at day 112 on chromosomes 13, 30, and 33 (batch one) and chromosomes 1 and 5 (batch two). However, apart from chromosomes 1, 5, and 13, which were reported earlier (Luo et al., 2013; Wang et al., 2015; Saelao et al., 2019), there were no other earlier studies that detected the presence of QTLs modulating NDV antibody response on chromosomes 30 and 33.
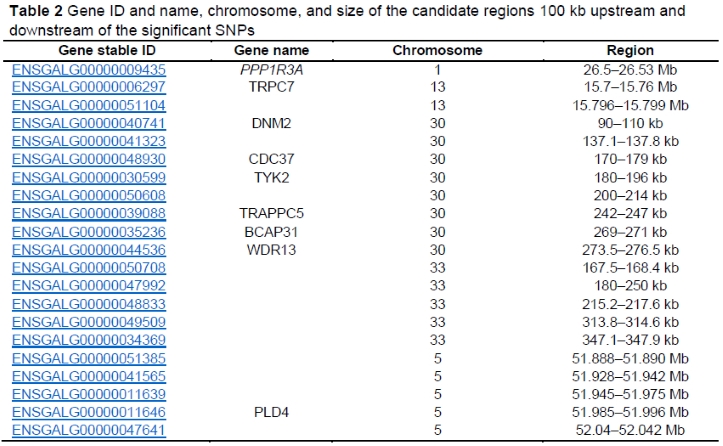
Candidate regions were defined as the genomic intervals 100 kb upstream and downstream of the significant SNPs detected in GWAS (Table 2) and annotated genes within those regions were identified. The list includes nine protein coding genes: protein phosphatase 1 regulatory subunit 3A (PPP1R3A), transient receptor potential cation channel subfamily C member 7 (TRPC7), dynamin-2-like (DNM2), cell division cycle 37 (CDC37), tyrosine kinase 2 (TYK2), trafficking protein particle complex 5 (TRAPPC5), B-cell receptor-associated protein 31 (BCAP31), WD repeat domain 13 (WDR13), and phospholipase D family member 4 (PLD4) (Table 2). The gene ontology annotation indicates that all these genes contribute to the regulation of cytokine activity, immune responses (innate and adaptive immunity), transcription, binding, transport, and development. Among these genes, six were found close to the significant SNPs (PPP1R3A, WDR13, FOXP2, BCAP31, TYK2, DNM2). These genes should be further interrogated to determine their relationship with Ab response to NDV in Sasso T451A populations in Ethiopia. Until now, none of these candidate genes for Ab response to NDV had been directly linked to the immunological response in chickens (Adhikari & Davie, 2018; Elbeltagy et al., 2019). Nevertheless, the functions of some of these genes have been explored in other animals (Zhou et al., 2014; Fleming et al., 2016; Zhuang et al., 2020).
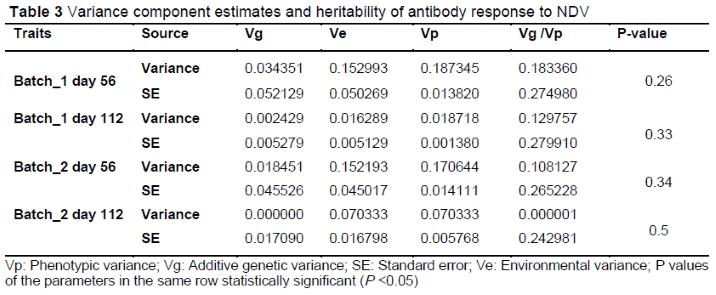
Heritability (h2) estimates were calculated as the ratio of the additive genetic variance (Vg) to the phenotypic variance (Vp = Vg + Ve) (Table 3). NDV Ab titres at Day 56 and Day 112 had estimated heritabilities of 0.09 ± 0.14 and 0.13 ± 0.15, respectively.
Results show that heritability estimates for NDV-specific Ab responses varied from 0.18 to 0.10 and from 0.000001 to 0.12 for day 56 and day 112, respectively. Overall, the heritability estimates were lower (0.000001-0.18) than previous reports on the same phenotypic trait (Lwelamira et al., 2009; Luo et al., 2013). However, many of these studies estimated heritability under indoor, standardized environmental conditions. Environmental factors can impact heritability, where genes involved in specific traits may respond differently according to environmental factors (Charmantier & Garant, 2005). To our knowledge, heritability estimates for the Ab response to NDV in Sasso T451A chicken are being reported for the first time in this work.
Conclusions
To our knowledge, this is the first study that has aimed to identify SNPs, genomic regions, and genes associated with Ab response to NDV in Sasso T451A chickens raised outdoor in semi-scavenging conditions in the Ethiopian highlands. Upon proper validation, our results may ultimately lead to new avenues for genetic improvement of Ab response to NDV. Understanding the genetic basis of NDV vaccine response will also assist in optimising vaccination protocols. The enhancement of genetic resistance to NDV will furthermore help in protecting the Ethiopian poultry industry and improve global food security. Moreover, the genes and variants revealed in this study demand more investigation to comprehend the underlying molecular pathways implicated in immune response in chickens.
Acknowledgments
The authors would like to acknowledge the following institutions and personnel for funding and facilitating the research. The International Livestock Research Institute (ILRI), Addis Ababa Poultry Research Facility members for their support in chicken monitoring, sample collection, and processing and the staff of the LiveGene program for their technical assistance, supported by the CGIAR Research Program on Livestock (CRP livestock project) sponsored by the CGIAR funding contributors to the Trust Fund (http://www.cgiar.org/about-us/our-funders/). The Bill and Melinda Gates Foundation and the United Kingdom Foreign Commonwealth and Development Office for financial support (Grant Agreement OPP1127286), under the auspices of the Centre for Tropical Livestock Genetics and Health (CTLGH), established jointly by the University of Edinburgh, Scotland's Rural College (SRUC), and ILRI. The findings and conclusions contained within are those of the authors and do not necessarily reflect positions or policies of the BMGF nor the UK Government.
Author contributions
MG carried out most of the experiments, data analysis, and manuscript draft; TD, AP, LV, OH, and GB conceived and designed the study; MG, KM, KS, WE, and BS carried out parts of the experiments; DM reviewed the manuscript; MG and OH designed a chicken facility for the natural challenge experiment. All authors read and approved the final manuscript.
Conflict of interest declaration
The authors declare that the research was conducted in the absence of financial or commercial relationships that could be considered a potential conflict of interest.
References
Adhikari, A. & Davie, J., 2018. JARID2 and the PRC2 complex regulate skeletal muscle differentiation through regulation of canonical Wnt signaling. Epigenetics Chromatin, 11(1), 1-20. [ Links ]
Anderson, C.A., Pettersson, F.H., Clarke, G.M., Cardon, L.R., Morris, A.P. & Zondervan, K.T., 2010. Data quality control in genetic case-control association studies. Nat. Protoc. 5(9), 1564-1573. [ Links ]
Astle, W. & Balding, D.J., 2009. Population structure and cryptic relatedness in genetic association studies. Stat. Sci. 24(4), 451-471. [ Links ]
Banos, G., Lindsay, V., Desta, T.T., Bettridge, J., Sanchez-Molano, E., Vallejo-Trujillo, A., Matika, O., Dessie, T., Wigley, P., Christley, R.M. & Kaiser, P., 2020. Integrating genetic and genomic analyses of combined health data across ecotypes to improve disease resistance in indigenous African chickens. Front. Genet. 11,543890. [ Links ]
Benjamini, Y., & Hochberg, Y., 1995. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B (Methodological), 57(1), 289-300. [ Links ]
Biscarini, F., Bovenhuis, H.V., Van Arendonk, J.A.M., Parmentier, H.K., Jungerius, A.P. & Van Der Poel, J.J., 2010. Across-line SNP association study of innate and adaptive immune response in laying hens. Anim. Genet. 41(1), 26-38. [ Links ]
Charmantier, A. & Garant, D., 2005. Environmental quality and evolutionary potential: Lessons from wild populations. Proc. R. Soc. B: Biol. Sci. 272(1571), 1415-1425. [ Links ]
Elbeltagy, A.R., Bertolini, F., Fleming, D.S., Van Goor, A., Ashwell, C.M., Schmidt, C.J., Kugonza, D.R., Lamont, S.J., Rothschild, M.F., 2019. Natural selection footprints among African chicken breeds and village ecotypes. Front. Genet. 10, 376. [ Links ]
EMDDI, 2017. Feasibility study for the establishment of chicken meat production and processing industry, Bishoftu, Ethiopia. [ Links ]
FAO, 2020. Meat market review: Emerging trends and outlook, FAO-Food and Agriculture Organization of the United Nations: Rome, Italy. [ Links ]
Fischer, U., Koppang, E.O., Nakanishi, T., 2013. Teleost T and NK cell immunity. Fish Shellfish Immunol. 35(2), 197-206. [ Links ]
Fleming, D.S., Koltes, J.E., Markey, A.D., Schmidt, C.J., Ashwell, C.M., Rothschild, M.F., Persia, M.E., Reecy, J.M., Lamont, S.J., 2016. Genomic analysis of Ugandan and Rwandan chicken ecotypes using a 600k genotyping array. BMC Genom. 17(1), 1-16. [ Links ]
Guèye, E.F., 2000. The role of family poultry in poverty alleviation, food security, and the promotion of gender equality in rural Africa. Outlook on Agriculture, 29(2), 129-136. [ Links ]
Hassen, H., Neser, F.W.C., Dessie, T., De Kock, A., Van Marle-Koster, E., 2006. Studies on the growth performance of native chicken ecotypes and RIR chicken under improved management system in north-west Ethiopia. Dev. 18(6). [ Links ]
Ioannidis, J., Thomas, G., Daly, M.J., 2009. Validating, augmenting, and refining genome-wide association signals. Nat. Rev. Genet. 10(5), 318-329. [ Links ]
Jie, H. & Liu, Y.P., 2011. Breeding for disease resistance in poultry: Opportunities with challenges. Worlds Poult. Sci. J. 67(4), 687-696. [ Links ]
Khawaja, T., Khan, S.H., Mukhtar, N., Parveen, A., Fareed, G., 2013. Production performance, egg quality and biochemical parameters of three-way crossbred chickens with reciprocal F1 crossbred chickens in sub-tropical environment. Ital. J. Anim. Sci. 12(1), e21. [ Links ]
Lamont, S.J., Pinard-van der Laan, M.H., Cahaner, A., van der Poel, J.J., Parmentier, H.K., 2003. 22 Selection for Disease Resistance: Direct Selection on the Immune Response. In: Poultry Genetics, Breeding and Biotechnology (Ed. by W.M. Muir & S.E. Aggrey). CAB International, Wallingford, Oxon OX10 8DE, UK. pp. 399-414. [ Links ]
Li, D.F., Lian, L., Qu, L.J., Chen, Y.M., Liu, W.B., Chen, S.R., Zheng, J.X., Xu, G.Y., Yang, N., 2013. A genome-wide SNP scan reveals two loci associated with the chicken resistance to Marek's disease. Anim. Genet, 44(2), 217-222. [ Links ]
Luo, C., Qu, H., Ma, J., Wang, J., Li, C., Yang, C., Hu, X., Li, N., Shu, D., 2013. Genome-wide association study of antibody response to Newcastle disease virus in chicken. BMC Genet. 14(1), 1-9. [ Links ]
Lwelamira, J., Kifaro, G.C., Gwakisa, P.S., 2009. Genetic parameters for body weights, egg traits, and antibody response against Newcastle Disease Virus vaccine among two Tanzania chicken ecotypes. Trop. Anim. Health Prod. 41(1), 51-59. [ Links ]
Malatji, D.P., Tsotetsi, A.M., Muchadeyi, F.C., Van Marle-Koster, E., 2016. A description of village chicken production systems and prevalence of gastrointestinal parasites: Case studies in Limpopo and KwaZulu-Natal provinces of South Africa. Onderstepoort J. Vet. Res. 83(1), 1-8. [ Links ]
McElroy, J.P., Kim, J.J., Harry, D.E., Brown, S.R., Dekkers, J.C.M., Lamont, S.J., 2006. Identification of trait loci affecting white meat percentage and other growth and carcass traits in commercial broiler chickens. Poult. Sci. 85(4), 593-605. [ Links ]
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., Garimella, K., Altshuler, D., Gabriel, S., Daly, M., DePristo, M.A., 2010. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20(9), 1297-1303. [ Links ]
Mekonnen, G., Mahendra, P., Tadelle, D., Wondmeneh, E., Wondu, M., Simenew K., 2013. Sero-prevalence of Newcastle Disease in poultry under backyard system in villages of Bishoftu Town, Ethiopia. Res. J. Poul. Sci. 6(2), 38-41. [ Links ]
NVI, 2016. Product Catalogue, User manual for live Newcastle Disease vaccine. Issue no. 2. 23. [ Links ]
Price, A.L., Patterson, N.J., Plenge, R.M., Weinblatt, M.E., Shadick, N.A., Reich, D., 2006. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 38(8), 904-909. [ Links ]
Psifidi, A., Banos, G., Matika, O., Desta, T.T., Bettridge, J., Hume, D.A., Dessie, T., Christley, R., Wigley, P., Hanotte, O., Kaiser, P., 2016. Genome-wide association studies of immune, disease, and production traits in indigenous chicken ecotypes. Genet. Sel. Evol. 48(1), 1-16. [ Links ]
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M.A., Bender, D., Maller, J., Sklar, P., De Bakker, P.I., Daly, M.J., Sham, P.C., 2007. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 81(3), 559-575. [ Links ]
Rothschild, D., Weissbrod, O., Barkan, E., Kurilshikov, A., Korem, T., Zeevi, D., Costea, P.I., Godneva, A., Kalka, I.N., Bar, N., Shilo, S., 2018. Environment dominates over host genetics in shaping human gut microbiota. Nature, 555(7695), 210-215. [ Links ]
Saelao, P., Wang, Y., Chanthavixay, G., Gallardo, R.A., Wolc, A., Dekkers, J.C., Lamont, S.J., Kelly, T., Zhou, H., 2019. Genetics and genomic regions affecting response to Newcastle Disease virus infection under heat stress in layer chickens. Genes. 10(1), 61. [ Links ]
Sarker, N., Tsudzuki, M., Nishibori, M., Yamamoto, Y., 1999. Direct and correlated response to divergent selection for serum immunoglobulin M and G levels in chickens. Poul. Sci. 78(1), 1 -7. [ Links ]
Turner, 2018. qqman: An R package for visualizing GWAS results using Q-Q and Manhattan plots. J. Open Source Softw. 3(25), 731. [ Links ]
Wang, D., Sun, Y., Stang, P., Berlin, J.A., Wilcox, M.A., Li, Q., 2009. Comparison of methods for correcting population stratification in a genome-wide association study of rheumatoid arthritis: Principal component analysis versus multidimensional scaling. BMC Proc. 3(7), 1-6. [ Links ]
Wang, W., Zhang, T., Zhang, G., Wang, J., Han, K., Wang, Y., Zhang, Y., 2015. Genome-wide association study of antibody level response to NDV and IBV in Jinghai yellow chicken based on SLAF-seq technology. J. Appl. Genet. 56(3), 365-373. [ Links ]
Yang, J., Lee, S.H., Goddard, M.E., Visscher, P.M., 2011. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 88(1), 76-82. [ Links ]
Yonash, N., Cheng, H.H., Hillel, J., Heller, D.E., Cahaner, A., 2001. DNA microsatellites linked to quantitative trait loci affecting antibody response and survival rate in meat-type chickens. Poult. Sci. 80(1), 22-28. [ Links ]
Zhang, G.X., Fan, Q.C., Zhang, T., Wang, J.Y., Wang, W.H., Xue, Q., Wang, Y.J., 2015. Genome-wide association study of growth traits in the Jinghai Yellow chicken. Genet. Mol. Res, 14(4), 15331-15338. [ Links ]
Zhou, H. Lamont, S.J., 2003. Chicken MHC class I and II gene effects on antibody response kinetics in adult chickens. Immunogenetics. 55(3), 133-140. [ Links ]
Zhou, X. Stephens, M., 2014. Efficient multivariate linear mixed model algorithms for genome-wide association studies. Nat. Methods. 11(4), 407-409. [ Links ]
Zhou, X.H., Yang, C.Q., Zhang, C.L., Gao, Y., Yuan, H.B., Wang, C., 2014. RASSF5 inhibits growth and invasion and induces apoptosis in osteosarcoma cells through activation of MST1/LATS1 signaling. Oncol. Rep. 32(4), 1505-1512. [ Links ]
Zhuang, Z., Xu, L., Yang, J., Gao, H., Zhang, L., Gao, X., Li, J., Zhu, B., 2020. Weighted single-step genome-wide association study for growth traits in Chinese Simmental beef cattle. Genes. 11 (2), 189. [ Links ]
Submitted 21 June 2023
Accepted 29 August 2023
Published 28 October 2023
# Corresponding author: 10110771@mylife.unisa.ac.za; m.girma@cgiar.org


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}