










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 n.5 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i5.04
Replacement of soybean with canola improves short-term milk yield and nitrogen-use efficiency in high-producing, early-lactation Holstein cows
F. MohammadiI; M.S.S. FirouzabadiII, #; M. SavariI; M.A. KachoieIII, IV; A.R. Rayshan T. SivapriyaV; F. AbdollahzadehVII
IDepartment of Animal Sciences, College of Agriculture, Isfahan University of Technology, Isfahan 84156-83111, Iran
IIDepartment of Clinical Sciences, Faculty of Veterinary Medicine, Ardakan University, P.O. Box 184, Ardakan, Iran
IIIDepartment of Medicinal Plants, Shahrekord Branch, Islamic Azad University, Shahrekord, Iran
IVMedicinal Plants Processing Center, Shahrekord Branch, Islamic Azad University, Shahrekord, Iran
VScientific Affairs Department, University of Al- Qadisiyah, Iraq
VIDepartment of Food Technology, Hindustan institute of Technology and Science, Padur Chennai
VIIDepartment of Animal Science, Tabriz University, Tabriz, Iran
ABSTRACT
It has been demonstrated that canola meal (CM) is used more efficiently than soybean meal (SBM) by lactating dairy cows. This study was conducted to determine the effects of replacing SBM with CM on production performance, blood metabolites, and ruminal fermentation parameters of high-producing dairy cows in early lactation. Twelve lactating Holstein cows (4 primiparous and 8 multiparous; 36 ± 8 days in milk) were used in a 3 × 3 Latin square design with three treatments and four replicates (cows) over three 21-day periods. Cows were fed diets formulated with incremental amounts of CM as 0% (CM-0), 50% (CM-50), or 100% (CM-100) of SBM replaced by CM. Experimental diets were formulated to be isonitrogenous (16.8% crude protein) and isoenergetic (NEL = 1.65 Mcal/kg DM). Dry matter intake (23.2 kg/d), milk composition, energy-corrected milk production (34.5 kg/d), total VFA, and blood metabolites were not affected by treatment. However, cows fed CM diets produced an average 2.9 kg/d more milk than control cows (CM-0). Apparent N efficiency was greatest in cows fed CM-50. Compared with the control, crude protein digestibility and ruminal ammonia concentration were greater and lower for cows fed CM-50 or CM-100, respectively. CM can be a potential substitute for SBM in dairy cow diets as its partial or complete substitution of SMB improved the production performance and nitrogen-use efficiency.
Keywords: canola meal, dairy cow performance, early lactation, soybean meal
Introduction
Previous research suggests that the efficiency of dietary crude protein (CP) in dairy cows is higher than in any other ruminants (Savari et al., 2018). It has been also demonstrated that canola meal is used more efficiently than soybean meal by lactating dairy cows (Paula et al., 2020). In general, finding the most efficient way of converting plant protein into milk production is the key to improve farm profitability and decrease the environmental emissions of dairy farms. Soybean meal (SBM) is the most commonly-used protein supplement for dairy and beef cattle. However, the increased demand for SBM and its concomitant elevation in price in recent years have motivated animal nutritionists to find potential alternatives for SBM with minimum adverse effect on the production performance of dairy cows. Canola meal (CM) is one of the promising alternatives in this area (Mulrooney et al., 2009; Maxin et al., 2013; Toti et al., 2018). Currently, CM is widely used in North America as a protein supplement for lactating dairy cows (Paula et al., 2018). Three comprehensive reviews (Hill 1991; Huhtanen et al. 2011; Martineau et al., 2013) concluded that feeding CM might be as effective as feeding SBM to lactating dairy cows. Inclusion of CM in dairy rations has been reported to increase dry matter intake (DMI), milk yield, and milk protein when compared with not only SBM (Huhtanen et al., 2011), but also other protein sources (Martineau et al., 2013). The positive effects of CM on milk production have been attributed to the increased supply of His (Shingfield et al., 2003) and, more recently, to increased absorption of essential amino acids, especially Met and Lys (Martineau et al., 2014). Research suggests that lactating cows would be less responsive to rumen-protected Met when fed CM than SBM (Broderick et al., 2015). Furthermore, recent research has indicated that dietary CP content can be lowered substantially without an adverse effect on milk yields in lactating cows (Kalscheur et al., 2006). Mulrooney et al. (2009) reported that CM as the main protein source, or in combination with dried distillers grains with solubles, was successfully used in the diets of lactating dairy cows. The positive responses in milk and milk protein yields to CM feeding have been attributed to the amino acid profile in the bypass fraction of CM (Brito et al., 2007). Although rather poor in Lys content, CM is rich in essential amino acids (especially His and Met; NRC 2001), whereas Met is the first-limiting amino acid in SBM (Illg et al., 1987; Toti et al., 2018). CM contains more Met and Cys but low amounts of Lys; using a blend of protein supplements (CM and SBM) has therefore been claimed to improve both microbial protein synthesis and amino acid profile (Broderick et al., 2015).
The present study was aimed at verifying the hypothesis that a combination of CM and SBM can improve production performance, ruminal parameters, blood metabolites, and nutrient digestibility in dairy cows. Thus, the objective of this study was to evaluate the partial or complete replacement of SBM with CM in diets of lactation dairy cows on performance, ruminal parameters, blood metabolites, and nutrient digestibility.
Materials and Methods
The study was conducted at the facilities of a dairy farm (FKA, Animal Husbandry and Agriculture Co., Isfahan, Iran) in compliance with the regulations and guidelines recommended by the Animal Care and Use Committee of the Iranian Council of Animal Care (1995).
Twelve lactating Holstein cows (8 multiparous and 4 primiparous, 38 ± 6 DIM) were used in a trial to evaluate the effects of different proportions of CM and SBM used as protein supplements. Cows were randomly assigned to one of three diets in a 3 × 3 Latin square design with three treatments and four replicates (cows), where 0%, 50% and 100% of the dietary SBM was replaced with CM, resulting in diets with 0%, 8.2% and 16.4% CM (DM basis), respectively (Table 2). The diets were formulated based on NRC (2001) in a manner to be isonitrogenous and isoenergetic. The duration of each experimental period was 21 d, with 15 d devoted to dietary adaptation and 6 d to data collection. Cows were held in free stalls throughout the experiment, had free access to water, and were weighed at the beginning and at the end of each period. All the cows were in good health and allowed 2 h of outdoor exercise every morning over the entire experimental period with free access to feed. Cows were fed twice daily (08:00 and 16:00) in amounts that permitted about 10% refusal daily.
Samples of individual feeds and orts (~0.5 kg) were taken on a daily basis and stored at -20 °C. Weekly composited samples from feeds and orts were dried at 60 °C for 48 h to be used for adjusting the as-fed composition of the diets. Cows were milked three times daily (06:00, 14:00, and 22:00) and milk weights were recorded at each milking. Milk samples were collected on days 16 to 21 of each period and preserved with potassium dichromate for future analysis. On day 21 of each period, 3 h after the morning feeding, samples of the rumen fluid (about 3 mL) were collected from the ventral sac via rumenocentesis (Nordlund & Garrett, 1994). The ruminal pH was immediately determined using a portable digital pH meter (Schott Titrator Titroline pH meter) and the samples were immediately frozen at -18 °C. Blood samples were also collected on day 21 of each period (~3 h after the morning feeding) from the coccygeal vessels using an evacuated tube containing an anticoagulant (EDTA). The samples were then centrifuged at 3,000 × g for 15 min. Plasma samples were separated, stored in plastic tubes, and frozen at -20 °C until analysis. Faecal grab samples were collected directly from the rectum of each cow (three times a day; in 8-h intervals) on days 18 to 21 of each period for the determination of the apparent digestibility of nutrients. The faecal samples were stored at -20 °C awaiting analysis. Spot urine was also collected (100 mL per sampling) during the last 4 d of each period at 06:00 and 18:00. The aliquots were acidified with sulphuric acid (0.072 N) and kept at -20 °C awaiting analysis (Lee et al., 2012). The urine and faecal samples were pooled for each collection period.
For chemical composition analysis, the weekly feed composites were ground through a 1-mm screen using a Wiley mill. The standard methods of the Association of Official Analytical Chemists (AOAC, 2002) were used for determination of DM (65 °C), ash (550 °C), crude protein (CP), and ether extract (EE). The concentration of rumen undegradable protein (RDP) was determined using the Cornell Net Carbohydrate and Protein System method (Licitra et al., 1996). Neutral detergent fibre (NDF; with inclusion of α-amylase and sodium sulphite) and acid detergent fibre (ADFom) contents were measured according to the procedure of Van Soest et al. (1991), using an Ankom2000 Fibre Analyzer (Ankom Technology Corporation, Macedon, NY, USA). Both NDF and ADF were ash-corrected. Apparent digestibility of nutrients was determined as the ratio of acid-insoluble ash method (Van Keulen & Young, 1977). Volatile fatty acids (VFA) in the rumen fluid were quantified by using a gas chromatograph (Chrompack, model CP-9002, Chrompack International BV, Middelburg, The Netherlands) using a 50-m (0.32 mm i.d.) fused-silica column (CP-Wax Chrompack Capillary Column; Varian Inc., Palo Alto, CA) and crotonic acid as the internal standard. Nitrogen was used as the carrier gas and oven initial and final temperatures were set at 55 °C and 195 °C, respectively. Detector and injector temperatures were set at 250 °C. Ammonia-nitrogen concentration in the rumen was determined using the colorimetric method (Broderick & Kang, 1980). Plasma levels of glucose, triglyceride, non-esterified fatty acids, beta-hydroxyl butyrate, total protein, albumin, globulin, and urea nitrogen were quantified using the commercial kits supplied by Pars Azmoon Company (Tehran, Iran). Milk samples were analysed for fat, true protein, lactose, solid non-fat, and total solids using Milkoscan (Foss Electric, Denmark). Milk urea-N concentration (MUN) was determined according to the Berthelot reaction (AOAC, 2002; ChemSpec 150 Analyzer, Bentley Instruments). Allantoin content in urine samples was determined according to the method of Young & Conway (1942). The concentration of creatinine in urine samples was quantified using a colorimetric method (Oser, 1965).
Data were analysed using PROC Mixed in SAS (SAS Institute 1999) for a replicated 3 × 3 Latin square design. The following model was used:
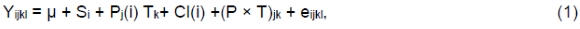
where, Yijkl is the dependent variable, μ is the overall mean, Si = fixed effect of square i, Pj(i) is the effect of period j, Tk is the fixed effect of treatment k, Cl is the random effect of cow k within square i, (P × T)jk is the interaction between period i and treatment j, and eijkl is the residual effect. Significance was declared at P <0.05 and trends at 0.05 <P <0.10.
Results and Discussion
The chemical composition of CM and SBM is presented in Table 1.
As expected, NDF and ADF contents of CM were greater than those of SBM, indicating that CM serves as a source of both CP and fibre; thus, the diets and fibre were balanced by increasing CM and decreasing barley grain and NFC, while crude protein was kept the same for all the treatments (equal to 16.8%; Table 2).
DMI tended to be greater (P = 0.10) in cows fed SMB diet than those fed CM (Table 3). The literature is inconsistent with regards to the substitution of SMB by CM; Maxin et al. (2013) studied the effect of substitution of SBM by CM in dairy rations and found no difference in DMI among treatments. However, Broderick et al. (2015) observed a positive effect in DMI only when CM was fed alone, but their combination had no effect on DMI, compared to SBM or CM diets. For unknown reasons, although we found about +1 kg difference in SBM diets compared to CM, BCS did not differ among treatments, which indicates that the cows fed CM diets produced more milk without using body deposits. Contrary to this observation, Robinson & Swanepoel (2018) found that BCS decreased linearly as the level of CM in the diet increased. The authors suggested that the maximum BCS gain occurred at a CM level of ~80g/kg DM in the diet. Contrary to the present study, Broderick et al. (2015) reported that feeding canola meal instead of soybean meal increased feed intake in lactating dairy cows.
Our findings are consistent with earlier observations (Martineau et al., 2013; Broderick et al., 2015) that reported greater milk yield when SBM and CM were compared for feeding lactating dairy cows. Huhtanen et al. (2011) suggested that greater milk production responses with CM inclusion could be due to the increased or more balanced amino acid supply, or both. Faverdin et al. (2003) suggested that the combination of SBM and CM provided a more palatable concentrate mixture than either SBM or CM alone. Martineau et al. (2014) reported that the positive effects of CM on milk production are attributed to increased His supply and, more recently, to increased absorption of essential amino acids (Shingfield et al., 2003). Contrary to the present study, Paula et al. (2018) reported that replacing SBM with CM did not affect milk yield in lactating dairy cows.
The greater N utilization in CM-50 might be related to the more efficient N metabolism with improved microbial protein synthesis or the greater supply of metabolizable protein (MP) from the rumen undegradable protein (RUP) fraction. Owing to the high degradability of CP and excellent amino acid profile of CM, Piepenbrink and Schingoethe (1998) suggested that RDP from CM favours microbial protein synthesis in the rumen.
The effects of treatment on MUN content are reported in Table 3. The MUN values decreased with CM usage in the ration (P <0.05). The lowest values assigned to the cows were fed CM-100, followed by those on CM-50; the highest was in cows fed CM-0. In agreement with this observation, Shingfield et al. (2003) found the lower urea-N concentrations in milk of cows fed a CM diet compared with those fed an SBM diet. In accordance with the results of the present study, Paula et al. (2020) reported that MUN concentration decreased when CM replaced SBM. Nousiainen et al. (2004) stated that dietary CP concentration was the most important nutritional factor influencing MUN. It seems that CM yields a better profile of amino acids than does SBM. Schwab et al. (2007) reported that Met and Lys are most often the first and second limiting amino acids for milk protein production, respectively. MP efficiency was observed to improve when Lys, Met, and other essential amino acids were supplied. Improved MP efficiency in the body reportedly led to reduced BUN and MUN levels in CM compared to the SBM diet (Wattiaux & Karg, 2004; Schwab et al., 2007; Toti et al., 2018).
The NRC (2001) model predicted an NEL-limited milk yield of 46.1 kg/d for all three experimental diets; the MP-limited milk yields of 38.4, 40.9, and 42.9 kg/d were also predicted by the same model for the diets containing 16.8% CP based on SBM and CM. Responses in milk yield indicated that NEL, MP, and amino acid supplies were greater than those predicted by NRC (2001) for all diets. Assuming that essential amino acids, such as Met and Lys, limited production, the predicted supply of digestible MP-RUP was lower than adequate for CM-0 and, due to the added CM, increased MP-RUP for the two lower diets (Table 2). In the present experiment, it appears that the diets containing CM provided the most favourable and optimal amino acid profiles in MP, which possibly improved the protein utilization efficiency (Schwab et al., 2003; Broderick et al., 2008).
Although the CP content of diets was equal in all of the treatments, the daily N intake varied and tended to decrease with decreasing DMI, whereas faecal N excretion did not change (Table 3). Faecal N primarily consists of indigestible microbial protein produced in the rumen, undigested feed proteins, endogenous proteins, sloughed cells from the gastrointestinal tract, as well as the microbial protein from hind gut fermentation (Davidson et al., 2003). In the current experiment, CP concentrations were found to be the same among the treatments. Since indigestible feed protein is a minor component of the total faecal N, differences among the treatments were not expected in light of the values of faecal N (Davidson et al., 2003). Predicted urinary N excretion did not differ among the groups. The decreased N efficiency with increasing CP degradability was expected. Broderick et al. (2015) reported that blending equal amounts of CP from both SBM and CM, when compared with that from SBM alone, decreased MUN and urinary N excretion at low CP concentrations. In support, Broderick et al. (2015) and Huhtanen et al. (2011) also suggested that that CP concentration and AA profile were the most important nutritional factors influencing MUN. The authors suggested that the improved quality of supplemental proteins was associated with the decreased MUN concentration.
Unlike N intake, milk N was not different among the treatments so that N utilization efficiency was higher in CM-50 than in CM-0. This may be attributed to the improved amino acid balance, especially Lys and Met and other essential amino acids in MP, which enhanced not only protein utilization, but also its apparent digestibility (Table 4) in the total tract (Schwab et al., 2007). Brito & Broderick (2007) did not find differences in N utilization efficiency between CM and SBM diets containing 16.6% CP.
Substitution of SBM by CM was observed to increase the apparent total tract digestibility of DM, CP, and OM. More recently, Robinson & Swanepoel (2018) found that apparent total tract digestibility of OM tended to increase linearly as the level of CM in the diet increased, whereas the CP digestibility remained unaffected. The increase in CP digestibility in the CM diet in the current study concurs with the findings of Paula et al. (2018), who reported greater mean apparent total-tract digestibility of DM, OM, and CP for the CM diet compared to the SMB diet. They found the increase in the amount of N truly digested in the rumen and an increase, therefore, in the RDP supply at the omasal canal in the CM diet compared to the SBM diet.
Ruminal fermentation patterns of dairy cows fed the different inclusions of CM are reported in Table 5. No difference was seen in ruminal pH, total VFA, propionate, butyrate, and acetate:propionate. However, cows fed CM-50 and CM-100 tended to have higher (P = 0.06) acetate concentrations than did cows fed CM-0. The replacement of CM for SBM led to a lower ammonia concentration in the rumen fluid, which is indicative of less ruminal degradation of CM protein.
Blood concentrations of glucose, triglyceride, p-hydroxybutyric acid, NEFA, BUN, total protein, and globulin remained largely unaffected by dietary treatment (Table 4). However, blood albumin and albumin: globulin were found to be greatest on the CM-100 diet. The higher plasma concentration of albumin in cows fed CM indicates that CM supplied more N to protein metabolism of lactating cows (Sanchez & Claypool, 1983).
Conclusion
The findings of the present study support the hypothesis that substituting SBM with CM in the diet had positive effects on milk production, nitrogen-use efficiency, and increased total tract digestibility of DM and CP in dairy cows. Canola meal as a total replacement for SBM or in combination with SBM, can be successfully used in dairy cow diets and improves the production performance and nitrogen-use efficiency of high-producing dairy cows.
Acknowledgments
We wish to thank J. Jalilnejad, General Manager of FKA Co., and his colleagues not only for supplying the animals required for the experiments but also for their financial support. The Head and staff members of the Research Station of the Nutrition Laboratory at the FKA Co. are also acknowledged. Our gratitude also goes to Dr. Ezzatollah Roustazadeh for editing the final version of this manuscript and to all others who helped us throughout this study.
Authors' Contributions
F.A and M.S (ORCID 0000-0002-1773-5287) participated in designing the study, laboratory analysis, and manuscript writing. MSSF was involved in drafting and revising the manuscript for intellectual content. All authors review and approved the manuscript before submitted for publication.
Conflicts of interest
The authors declare no conflicts of interest.
References
AOAC International, 2000. Official Methods of Analysis. 17th ed. AOAC International, Gaithersburg, MD. [ Links ]
Brito, A.F., Broderick, G.A., Colmenero, J.O. & Reynal, S.M., 2007. Effects of feeding formate-treated alfalfa silage or red clover silage on omasal nutrient flow and microbial protein synthesis in lactating dairy cows. J. Dairy Sci. 90, 1392-1404. [ Links ]
Brito AF, Broderick GA, 2007. Effects of different protein supplements on milk production and nutrient utilization in lactating dairy cows. J. Dairy Sci. 90,1816-1827. [ Links ]
Broderick, G.A., Faciola, A.P. & Armentano, L.E., 2015. Replacing dietary soybean meal with canola meal improves production and efficiency of lactating dairy cows. J. Dairy Sci. 98, 5672-5687. [ Links ]
Broderick, G.A. & Kang, J.H., 1980. Automated simultaneous determination of ammonia and total amino-acids in ruminal fluid and in vitro media. J. Dairy Sci. 63, 64-75. [ Links ]
Broderick, G.A., Stevenson, M.J., Patton, R.A., Lobos, N.E. & Colmenero, J.O., 2008. Effect of supplementing rumen-protected methionine on production and nitrogen excretion in lactating dairy cows. J. Dairy Sci. 91,1092-1102. [ Links ]
Christen, K.A., Schingoethe, D.J., Kalscheur, K.F., Hippen, A.R., Karges, K.K. & Gibson, M.L., 2010. Response of lactating dairy cows to high protein distillers grains or three other protein supplements. J. Dairy Sci. 93, 2095-2104. [ Links ]
Davidson, S., Hopkins, B.A., Diaz, D.E., Bolt, S.M., Brownie, C., Fellner, V. & Whitlow, L.W., 2003. Effects of amounts and degradability of dietary protein on lactation, nitrogen utilization, and excretion in early Holstein cows. J. Dairy Sci. 86, 1681-1689. [ Links ]
Edmondson, A.J., Lean, D., Weaver, I.D., Farver, T. & Webster, G., 1989. A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 72, 68-78. [ Links ]
Faverdin, P., M'hamed, D. & Vérité, R., 2003. Effects of metabolizable protein on intake and milk production of dairy cows independent of effects on ruminal digestion. Anim. Sci. 76, 137-146. [ Links ]
Hill, R., 1991. Rapeseed meal in the diets of ruminants. Nutrition Abstracts and Reviews. Series B, Livestock Feeds and Feeding (United Kingdom). [ Links ]
Huhtanen, P., Hetta, M. & Swensson, C., 2011. Evaluation of canola meal as a protein supplement for dairy cows: A review and a meta-analysis. Canadian J. Anim. Sci. 91, 529-543. [ Links ]
Illg, D.J., Sommerfeldt, J.L. & Schingoethe, D.J., 1987. Lactational and systemic responses to the supplementation of protected methionine in soybean meal diets. J. Dairy Sci. 70, 620-629. [ Links ]
Iranian Council of Animal Care, 1995. Guide to the care and use of experimental animals. Vol. 1. Isfahan Univ. Technol., Isfahan, Iran. [ Links ]
Kalscheur, K.F., Vi, R.B., Glenn, B.P. & Kohn, R.A., 2006. Milk production of dairy cows fed differing concentrations of rumen-degraded protein. J. Dairy Sci. 89(1), 249-259. [ Links ]
Kohn, R.A., Kalscheur, K.F., Russek-Cohen, E., 2002. Evaluation of models to estimate urinary nitrogen and expected milk urea nitrogen. J. Dairy Sci. 85, 227-233. [ Links ]
Lee, C., Hristov, A.N., Cassidy, T.W., Heyler, K.S., Lapierre, H., Varga, G.A., De Veth, M.J., Patton, R.A. & Parys, C., 2012. Rumen-protected lysine, methionine, and histidine increase milk protein yield in dairy cows fed a metabolizable protein-deficient diet. J. Dairy Sci. 95,6042-56. [ Links ]
Martineau, R., Ouellet, D.R. & Lapierre, H., 2013. Feeding canola meal to dairy cows: A meta-analysis on lactational responses. J. Dairy Sci. 96, 1701 -1714. [ Links ]
Martineau, R., Ouellet, D.R. & Lapierre, H., 2014. The effect of feeding canola meal on concentrations of plasma amino acids. J. Dairy Sci. 97, 1603-1610. [ Links ]
Mason, V.C., 1969. Some observations on the distributions and origin of in sheep feces. J. Agri. Sci. 73, 99-111. [ Links ]
Maxin, G., Ouellet, D.R. & Lapierre, H., 2013. Effect of substitution of soybean meal by canola meal or distillers grains in dairy rations on amino acid and glucose availability. J. Dairy Sci. 96,7806-7817. [ Links ]
Mulrooney, C.N., Schingoethe, D.J., Kalscheur, K.F. & Hippen, A.R., 2009. Canola meal replacing distillers grains with solubles for lactating dairy cows. J. Dairy Sci. 92, 5669-5676. [ Links ]
Nordlund, K.V. & Garrett, E.F., 1994. Rumenocentesis: A technique for the diagnosis of subacute rumen acidosis in dairy herds. Bovine Pract. 28, 109-112. [ Links ]
Nousiainen, J., Shingfield, K.J. & Huhtanen, P., 2004. Evaluation of milk urea nitrogen as a diagnostic of protein feeding. J. Dairy Sci. 87, 386-398. [ Links ]
NRC, 2001. Nutrient Requirements of Dairy Cattle. 7th rev. ed. National Academy of Sciences, Washington, D; 2001. [ Links ]
Oser, B.L., 1965. Hawk's Physiological Chemistry (14th ed). McGraw Hill, New York, pp. 1233-1238. [ Links ]
Paula, E.M., Broderick, G.A., Danes, M.A., Lobos, N.E., Zanton, G.I. & Faciola, A.P., 2018. Effects of replacing soybean meal with canola meal or treated canola meal on ruminal digestion, omasal nutrient flow, and performance in lactating dairy cows. J. Dairy Sci. 101, 328-339. [ Links ]
Paula, EM., Broderick, GA. & Faciola, A. P., 2020. Effects of replacing soybean meal with canola meal for lactating dairy cows fed three different ratios of alfalfa to corn silage. J. Dairy Sci. 103(2), 1463-1471. [ Links ]
Piepenbrink, M.S. & Schingoethe, D.J., 1998. Ruminal degradation, amino acid composition, and estimated intestinal digestibilities of four protein supplements. J. Dairy Sci. 81, 454-461. [ Links ]
Robinson, P.H. & Swanepoel, N., 2018. Impacts of increasing levels of canola meal in diets of high producing Holstein cows on their productive performance. Anim. Feed Sci. Technol. 237, 175-185. [ Links ]
Sadri, H., Rahmani, H.R., Khorvash, M., Ghorbani, G.R. & Bruckmaier, R.M., 2009. Chromium supplementation and substitution of barley grain with corn: Effects on performance and lactation in periparturient dairy cows. J. Dairy Sci. 92, 5411-5418. [ Links ]
Sánchez, J.M. & Claypool, D.W., 1983. Canola meal as a protein supplement in dairy rations J. Dairy Sci. 66, 80-85. [ Links ]
SAS Institute, 1999. SAS User's Guide. Version 8 ed. SAS Institute Inc., Cary, NC. [ Links ]
Savari, M., Khorvash, M., Amanlou, H., Ghorbani, G. R., Ghasemi, E. & Mirzaei, M., 2018. Effects of rumen-degradable protein: Rumen-undegradable protein ratio and corn processing on production performance, nitrogen efficiency, and feeding behavior of Holstein dairy cows. J. Dairy Sci. 101(2), 1111-1122. [ Links ]
Schwab, C.G., Boucher, S.E. & Sloan, B.K., 2007. Metabolizable protein and amino acid nutrition of the cow: Where are we in 2007? Proc. Cornell Nutr. Conf. Feed Manufacturers. Pp. 121-138. [ Links ]
Schwab, C.G., Ordway, R.S. & Whitehouse, N.L., 2003. Amino acid balancing in the context of MP and RUP requirements. In: Proc. Four-State Applied Nutrition and Management Conference. LaCrosse, WI. Midwest Plan Service, Iowa State University, Ames, Pp. 25-34. [ Links ]
Shingfield, K.J., Vanhatalo, A. & Huhtanen, P., 2003. Comparison of heat-treated rapeseed expeller and solvent extracted soya-bean meal as protein supplements for dairy cows given grass silage-based diets. Anim. Sci. 77, 305-317. [ Links ]
Sjaunja, L.O., Baevre, L., Junkkarinen, L., Pedersen, J. & Setälä, J., 1990, A Nordic proposal for an energy corrected milk (ECM) formula. In: 27th Session of the International Commission for Breeding and Productivity of Milk Animals, Paris, France. Wageningen Academic Publishers, Wageningen, the Netherlands. Pp. 156-157. [ Links ]
Sniffen, C.J., O' Connor, J.D., Van Soest, P.J., Fox, D.G. & Russell, J.B., 1992. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 70, 3562-3577. [ Links ]
Van Keulen, J.Y. & Young, B.A., 1977. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci. 44, 282-287. [ Links ]
Van Soest, P.J., 1994. Nutritional Ecology of the Ruminant. 2nd. Cornell University Press, Ithaca, NY. [ Links ]
Van Soest, P.J., Robinson, J.B. & Lewis, B.A., 1991. Methods for dietary fibre, neutral detergent fibre, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Wattiaux, M.A. & Karg, K.L., 2004. Protein level for alfalfa and corn silage-based diets: I. Lactational response and milk urea nitrogen. J. Dairy Sci. 87, 3480-3491. [ Links ]
Young, E.G. & Conway, C.F., 1942. On the estimation of allantoin by the Rimini-Schryver reaction. J. Biol. Chem. 142, 839. [ Links ]
Submitted 18 February 2022
Accepted 20 October 2022
Published 23 October 2023
# Corresponding author: E-mail: m.s.safaee@ardakan.ac.ir


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}