










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 no.4 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i4.14
Genetic and phenotypic trends for female fertility traits derived from service records of South African Holstein cattle
R.D. KgariI, II, #; C.J.C. MullerII; K. DzamaII; M.L. MakgahlelaI, III
IAgricultural Research Council, Private Bag X2, Irene 0062, South Africa
IIDepartment of Animal Sciences, University of Stellenbosch, Stellenbosch 7600, South Africa
IIIUniversity of the Free State, Department of Animal, Wildlife and Grassland Sciences, PO Box 339 Bloemfontein 9301, South Africa
ABSTRACT
The objective of this study was to estimate breeding values (EBV) and genetic and phenotypic trends for female fertility traits derived from artificial insemination (AI) records. Data were collected from automated milk recording systems: 64464 records from 18 South African (SA) Holstein herds. Breeding values were estimated for heifer traits, i.e., age at first service (AFS), age at first calving (AFC), number of services per conception (SPCh); and for cow traits, i.e., calving to first service (CFS), number of days open (DO), number of services per conception (SPC), first service within 80 days post-partum (FS80d), and whether cows were confirmed pregnant within 100 days post-partum (PD100d) or within 200 days post-partum (PD200d) using THRGIBBSF90. Trends were estimated for the period 1984 to 2011. There was no clear trend on average breeding values for heifer traits. However, phenotypic trends for heifer traits showed a desirable downward trend, with a decrease of 0.14 and 0.13 months/year for AFS and AFC, respectively. Phenotypic trends for cow fertility traits appeared to increase at the rate 0.16 and 0.83 days/year for CFS and DO, respectively, with animals requiring 0.02 more services per year for SPC. The average EBV for binary traits FS80d, PD100d, and PD200d, showed no distinct trend. In general, there was no clear genetic and phenotypic trend for all traits defined. The trends that appeared to follow trait improvement could have been coincidental. Thus, efforts have to be made towards the improvement of heifer and cow fertility traits explored in this study.
Keywords: breeding values, genetic improvement, phenotypic means, THRGIBBSF90
Background
Until the 1990s, the main breeding objective of many dairy-producing countries was to increase yields of milk, protein, and fat (Miglior et al., 2005). This resulted in a deterioration of overall fitness, including health and fertility, because of the antagonistic association between fertility and milk yield (Van Arendonk et al., 1989; Bagnato & Oltenacu, 1994; De Jong, 1998; Kadermideen, 2004; Pryce et al., 2004). The decreasing reproductive performance in dairy cattle has a negative impact on the profitability of dairy herds (Britt,1985; Dijkhuizen et al., 1985), where high producing cows require more veterinary treatments and more AI services per conception, i.e., increasing input costs of production thereby extending the days open and the intercalving period. Owing to its economic importance, producers moved towards more balanced breeding programmes, which integrated genetic improvements in health, longevity, and fertility with production (Miglior et al., 2005).
In South Africa, genetic evaluations of fertility are based on age at first calving (AFC) and calving interval (CI) (Makgahlela et al., 2008). Although this is a good step towards the improvement of fertility, cows that do not have subsequent calving dates or are culled for not falling pregnant are excluded in genetic evaluations (Muller et al., 2018). This limits the potential use of CI as a selection criterion for fertility in the genetic evaluation of dairy cows, as information on the perceived least fertile group of cows is excluded; AFC is problematic as it is highly influenced by the breeder's decision to start breeding. Artificial insemination records provide information on additional reproductive performance that can be useful for inclusion in genetic evaluations. In any genetic improvement program, it is important to track the results and monitor the progress and effectiveness of a genetic selection program by evaluating the changes in genetic trends over time. A genetic trend is defined as a change in performance per unit of time due to a change in mean breeding value (Canaza-Cayo et al., 2016). A phenotypic trend is defined as a change in performance per unit time due to change in a phenotypic mean. The change indicates an improvement or deterioration in the desirable qualities of the animal, depending on the direction of the trend.
In animal improvement, it is important for breeders and farmers to assess genetic trends in order to monitor whether breeding objectives are achieved and are trending in the intended direction. Studies of genetic and phenotypic trends for fertility traits reported an increasing trend of 1.90, 1.25, and 1.34 days per year for calving intervals in the South African Holstein population (Makgahlela et al., 2008; Mostert et al., 2010; and Ramatsoma et al., 2014; respectively). These increasing trends in CI indicate a deterioration in post-partum fertility in the South African Holstein population.
The objective of the current study was to estimate phenotypic and genetic trends for fertility measures derived from AI service records in South African Holstein cattle.
Materials and Methods
Data consisted of artificial insemination records (n = 64 464) of heifers and cows, which were collected from 18 South African Holstein herds using on-farm automated milk recording system (DIMSSA). These data were used to derive the following AI service-based heifer and cow fertility traits: age at first service (AFS), age at first calving (AFC), number of services per conception for heifers (SPCh), calving to first service (CFS), number of days open (DO), number of services per conception for cows (SPC), first service within 80 days post-partum (FS80d), whether cows were confirmed pregnant within 100 days post-partum (PD100d), and whether the cows were confirmed pregnant within 200 days post-partum (PD200d).
Non-interval CFS80d, PD100, and PD200 traits were recorded as binary traits coded as 1 = no and 2 = yes. Data editing was carried out using the R-CRAN program (R Core Team, 2017). Two subsets were extracted from the edited dataset for heifer (1st parity only) and cow (2nd parity and above) traits. The datasets were edited to remove outliers and to meet an acceptable threshold for each trait. After editing, the final data available for statistical analysis consisted of 10 019 and 24 909 records for heifers and cows, respectively. The pedigree consisted of information on 18 592 animals born between 1981 and 2013. Pedigree data included individual animal birth dates, service and calving dates, lactation numbers of dams, and dam and sire identification numbers. These data were edited to remove animals with unknown birth dates, leaving 14 323 records for statistical analysis.
Descriptive statistics were computed and histograms were plotted using the Hist function in R (R Core Team, 2017). The lme4 package of Bates et al. (2015) implemented in R-CRAN was used to test non-genetic factors associated with heifer and cow fertility traits. The analysis of variance (ANOVA) function in R was used to test the most significant effects associated with the traits. The effects tested were HYS of birth or calving, age of insemination or calving, and the lactation number, which all affected fertility traits (P <0.05). The effects were included in the multivariate analysis to estimate heritabilities and EBV using THRGIBBS1F90 and POSTGIBBSF90 of BLUPF90 (Misztal et al., 2016). The following animal models were fitted for heifer (SPCh, AFS and AFC) and cow (SPC, CFS, DO, CFS80d, PD100d and PD200d) traits:
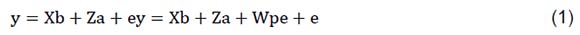
where: y is the vector of observations; b is the vector of fixed effects, which consists of herd-year-season of birth (for heifer traits), herd-year-season of calving (for cow traits), parity fitted only (for cow traits), age at insemination fitted only for (SPCh), and age at calving fitted (for cow traits) as a covariate; a was the vector of additive genetic effects; pe was the vector of random permanent environmental effects (fitted only for cow traits); e was the vector of residual effects; and X, Z, and W were the corresponding incidence matrices. It was assumed that the expectation E of the variables was:

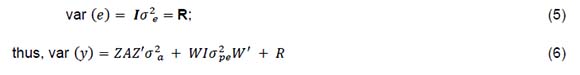
The genetic trends were then determined using the regression of EBV of the animals on the year of birth using the dplyr package and plotted using the ggplot2 package in R CRAN (Wickham, 2016; Wicklam et al., 2018). The phenotypic trends were estimated using average of phenotypic values over the birth years, and visualized in R using ggplot2 (Wicklam, 2016).
Results and Discussion
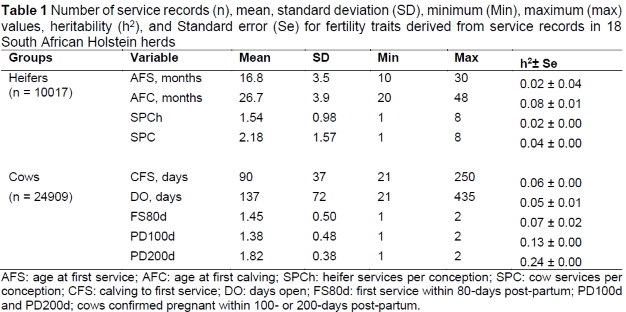
Descriptive statistics of the data, heritability estimates, and standard errors are presented in Table 1. The average SPCh in heifers was lower (1.54) than in cows (2.18), indicating that young females require fewer inseminations per conception than lactating cows. This is somewhat expected as reduced fertility in older animals could probably be due to lactation stress (Badinga et al., 1985); lactating cows are in a state of negative energy balance, which has subsequent effects on fertility, leading to poor conception rates (Wathes et al., 2007). On average, it took 90 days for the cows to be ready for first service after calving. High CFS intervals lead to greater DO, which averaged at 137 days in this study. This subsequently extended CI beyond the recommended 365 days, even though heat detection was good and high pregnancy rates followed.
Heritability estimates for all traits ranged from low (0.02 ± 0.00) to moderate (0.24 ± 0.00). Previously reported heritability estimates for AFS were 0.24 in Dutch Friesians (Jansen et al., 1987) and 0.12 in Canadian Holsteins (Raheja et al., 1989), whereas the estimate for AFC (0.08) was comparable to that of 0.09, obtained for the Kenyan Ayrshire (Amimo et al., 2006), although higher than 0.03 reported for Iranian Holstein cows using a single trait animal model (Eghbalsaied, 2006).
The SPC was defined for both heifers and cows, and the heritability estimate of SPC in heifers was slightly lower (0.02) than the corresponding cow trait (0.04). Differences between the estimates of heritability observed in this study and estimates from other studies could be due to the genetic variation among the populations, different statistical models used for analysis, varying reactions of the same breed to different environmental conditions, or differences in farm management strategies.
The heritability estimate for first service within 80 days post-partum was 0.07 and for the two binary traits (indicating whether cows were confirmed pregnant within 100 or 200 days post-partum) were 0.13 and 0.24, respectively. The low to moderate heritability of the traits indicates that heifer and cow fertility traits in this study are underlined by the genetic mechanisms, thereby warranting their genetic improvement through objective selection programmes.
A sound dairy breeding objective should include all traits that are economically relevant in dairy production. The Nordic countries (i.e., Denmark, Sweden, and Finland) were the first to include health and fertility traits into selection programmes in the mid 1990's. whereas other countries, such as New Zealand, followed in 1998, and the USA, in 2001 (Wesseldijk, 2004). The inclusion of fertility traits in South African selection programs was recommended in 2007 (Makgahlela et al., 2007). Genetic parameters for calving intervals were estimated and published in 2010 for use as an indicator trait for fertility in South African dairy breeds (Mostert et al., 2010). The key indicator traits for fertility in South African selection programs are calving interval (CI) and age at first calving (AFC); this may be one of the reasons for the observed results in this study, which showed little to no genetic improvement for the studied fertility traits.
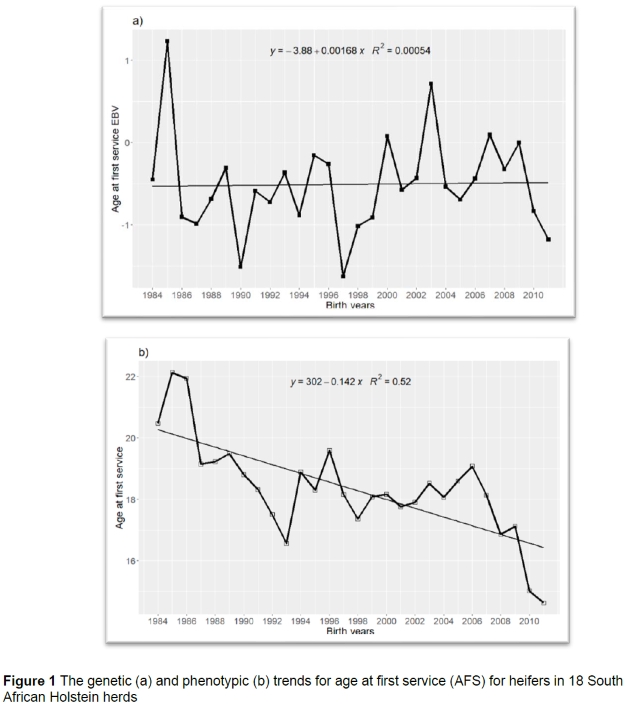
The average EBV for the studied traits generally showed no distinct trends, with the exception of cow linear traits, CFS and DO, which showed decreasing trends of small magnitudes. There were slight improvements in phenotypic means over the period 1984-2011. The genetic average in all the heifer fertility traits; AFS (0.002 ± 0.00), AFC (0.001 ± 0.00) and SPCh (-0.001 ± 0.00) was close to zero, indicating no particular trend. The example is illustrated in Figure 1(a) of a genetic trend of AFS. The mean EBV shows an increase from 1984 (-0.45) to 1985 (1.22), then decreases to -0.90 in 1986. In most years the mean EBV was negative with the lowest being (-1.51,-1.63, and -1.18) in 1990, 1997, and 2011, respectively. The average EBV fluctuated, indicating that there was no genetic improvement in the trait, AFS.
The phenotypic trend (Figure 1b) for AFS generally decreased over the years. There was a slight decrease of 1 month/year from 1984 to 1993, and an annual increase of 2 months/year from 1993 to 1994. The average decrease of (-0.14 months/year) was observed over the years for the phenotypic trend. The decrease was also observed in the phenotypic trend of AFC (-0.14 months/year). However, the phenotypic trend for SPCh was close to zero (0.003), indicating no particular trend. The phenotypic decrease in AFS and AFC was desirable; however, this could be due to improvement of the environmental conditions of the farm and a management decision to breed animals early. The decreasing trend might also represent earlier maturity from better calf rearing and good nutrition because feeding level in young animals affects the age at which they reach puberty (Shortle, 2014).
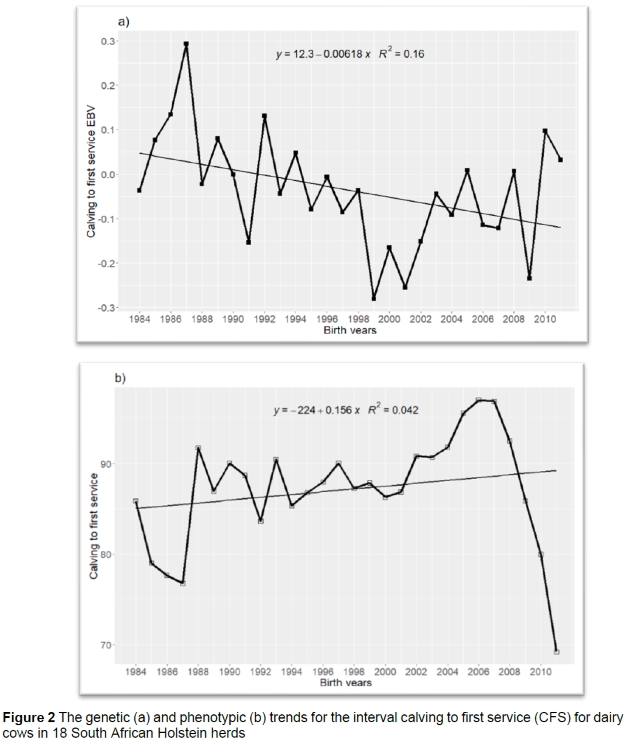
The genetic trends for cow linear traits showed a decrease of a small magnitude for CFS (-0.01 ± 0.00) and DO (-0.06 ± 0.00) days/year, while for SPC there was no particular trend as the slope of regression was zero (0.00). An example is illustrated in Figure 2(a) of a genetic trend in CFS whereby the average EBV of CFS tended to decrease from 1984 to 2011. The genetic trend of CFS is going in a favourable direction, although the decrease is very close to zero (-0.01) days/year. The current results are in agreement with report of Ghiasi et al. (2016), who reported a decrease of (-0.062) days/year for interval to first service. This genetic trend indicates that there is a genetic potential for reducing CFS following selection in the direction favouring the trait. In Nordic countries (Sweden, Finland, and Denmark), including traits such as CFS and SPC in the Total Merit Index (TMI) has resulted in improved fertility, which has saved input costs (Kargo et al., 2014).
Economic value is defined as the effect of a unit change in the phenotypic level of the trait in question, ceteris paribus, on profit. An unfavourable phenotypic trend was observed in all the linear cow traits, with average increases of 0.16 days/year for CFS, 0.83 days/year for DO, and 0.02 per year for SPC. This indicates that cows required more services to conceive and the length of days from calving to conception increased over the study period. This may be due to management delaying first service post-partum to overcome the detrimental effects of negative energy balance on fertility (Niozas et al., 2019). However, this strategy has a negative impact on the economics of dairy enterprises because DO is directly linked to CI due to the common physiological mechanisms controlling these traits (Ayalew et al., 2017). Banga et al. (2009) calculated the cost of one extra day in ICP as R5.75. The unfavourable phenotypic trend can be counter-acted through improved management strategies.
The genetic average in all the binary traits; FS80d (0.001 ± 0.00), PD100d (0.001 ± 0.00), and PD200d (0.001 ± 0.00) was close to zero, indicating a very slow increase. Figure 3a indicates the genetic trend for PD100d, whereby the rate of increase was very slow (0.0008). This shows that not much has been done to improve the traits through genetic selection. There was no distinct phenotypic trend for FS80d, indicating that there has not been much change in the number of cows returning to service within 80 days after calving. The phenotypic average of PD100d and PD200d showed a decrease of -0.006 and -0.003 days/year, respectively, indicating that more cows became pregnant later than 100 and 200 days postpartum (Figure 3b).
The studied traits showed no distinct genetic trends over the years as the EBV was fluctuating, whereas there were some observed phenotypic changes Hence, good nutrition and improved management strategies can cause a phenotypic improvement in fertility, while no impactful improvement is made genetically. The current results indicate that there is little to no genetic improvement in the studied Holstein population.
Banga & Rautenbach (1999) indicated that the primary focus of breeding programs in South Africa has been on increased milk production, which could be one of the reasons for the observed genetic trends in the current study. Several studies on the genetic relationships between fertility and production traits found that correlations between milk yield and fertility traits were antagonistic and statistically significant. An observation of 0.69 (Makgahlela et al., 2007), 0.59 (Toghiani, 2012), and 0.68 (Sahin et al., 2014) on the correlation between 305-d milk yield and calving interval could have negative long-term effects on the economics of dairy production as the economic importance of fertility has been demonstrated in previous studies (Visscher et al., 1994; Holmes et al., 2000; Banga et al., 2009; Toghiani, 2012). Selective animal breeding ensures a continuous improvement in farm animals, generation after generation. In the countries that include functional traits earlier in their breeding objectives, trends in functional traits have now improved in recent years (Berry et al., 2014). The use of genomic tools for selection of low heritable traits, such as fertility, in developed countries has resulted in rapid genetic gains in dairy cattle (Hutchison et al., 2014).
Conclusion
The average EBV for heifer traits in the studied population showed no distinct trend, whereas for cow traits there were some observed, favourable trends of small magnitude. Favourable phenotypic trends were observed for heifer fertility traits. This may not lead to an improvement in heifer fertility due to the overall undesirable genetic trends. The phenotypic improvement might have been from good management and environmental conditions, while the problem still lies genetically. Eighteen South African Holstein herds showed no genetic gains for fertility over the study period. Heritability estimates indicate that there is some genetic aspect of the fertility traits that could facilitate improvement through genetic selection, which is imperative for long-term benefits. The studied fertility traits could be used in addition to AFC and ICP to serve as basis for selection of reproduction in dairy cattle to minimize selection bias and improve fertility in the South African Holstein population. A balanced breeding program that includes all traits of economic importance could accelerate the improvement of fertility in South African dairy herds.
Acknowledgements
The authors would like to acknowledge National Research Foundation, Dairy Genomics Project, and the Technology Innovation Agency as an implementing agency for the Department of Science and Innovation for financial support. DIMSSA for providing service records and HPC for providing a computing environment for data analysis are gratefully acknowledged.
Author's contributions
RDK assembled and analysed the data, designed, and wrote the initial draft of the manuscript; MLM drafted and reviewed the manuscript; CJCM and KD critically evaluated the manuscript.
Conflict of interests
Authors declare that they have no conflict of interests.
References
Amimo, J.O., Mosi, R.O., Wakhungu, J.W., Muasya, T.K., Inyangala, B.O., 2006. Phenotypic and genetic parameters of reproductive traits for Ayrshire cattle on large-scale farms in Kenya. Livest. Res. Rural Dev.18, 10. [ Links ]
Ayalew, W., Aliy, M. Negussie, E., 2017. Estimation of genetic parameters of the productive and reproductive traits in Ethiopian Holstein using multi-trait models. Anim Biosci. 30, 11, 1550-1556. doi.org/10.5713/ajas.17.0198 [ Links ]
Badinga, L., Collier, R.J., Thatcher, W.W., Wilcox, C.J., 1985. Effects of climatic and management factors on conception rate of dairy cattle in subtropical environment. J. Dairy Sci. 68(1), 78-85. doi.org/10.3168/jds.S0022-0302(85)80800-6. PMID: 3980812. [ Links ]
Bagnato, A., Oltenacu, P.A., 1994. Phenotypic evaluation of fertility traits and their association with milk production of Italian Friesian cattle. J. Dairy Sci. 77, 874-882. [ Links ]
Banga, C.B., Rautenbach, L., 1999. Genetic progress from 1980 to 1999. Annual Report National Dairy Cattle Performance Testing Scheme South Africa. ARC Animal Improvement Institute, 6-11. [ Links ]
Banga, C.B., Neser, F.W.C., van der Westhuizen, J., Garrick, D.J., 2009. Economic values for dairy production traits under different milk payment systems in South Africa. S. Afr. J. Anim. Sci. 39, 112-115. [ Links ]
Berry, D.P., Wall, E., Pryce, J.E., 2014. Genetics and genomics of reproductive performance in dairy and beef cattle. Animal. 8, 105-121. [ Links ]
Britt, J.H., 1985. Enhanced reproduction and its economic implications. J. Dairy Sci. 68(6), 1585-1592. [ Links ]
Canaza-Cayo, A. W., Cobuci, J. A., Lopes, P. S., Torres, R. A., Martins, M. F., Daltro, D. S., Silva, M. V. G. B., 2016. Genetic trend estimates for milk yield production and fertility traits of the Girolando cattle in Brazil. Livest. Sci. 190,113-122. https://doi.org/101016/j.livsci.2016.06.009. [ Links ]
De Jong, G., 1998. Index for daughters' fertility in the Netherlands. Interbull Bull. 18, 102-105. [ Links ]
Dijkhuizen, A.A., J. Stelwagen, Renkema, J.A., 1985. Economic aspects of reproductive failure in dairy cattle. Financial loss at farm level. Prev. Vet. Med. 3, 251-263. [ Links ]
Eghbalsaied, S., 2011. Estimation of genetic parameters for 13 female fertility indices in Holstein dairy cows. Trop. Anim. Health Prod. 43, 811-816. [ Links ]
Ghiasi, H., Honarvar, M., 2016. Genetic and phenotypic trends of fertility traits in Iranian Holstein cows. Iran. J. Appl. Anim. Sci. 6. [ Links ]
Hutchison, J. L., Cole, J. B., Bickhart, D. M., 2014. Short communication: Use of young bulls in the United States. J. Dairy Sci. 97, 3213-3220. doi: 10.3168/jds.2013-7525. [ Links ]
Jansen, J., van der Werf, J., de Boer, W. 1987. Genetic relationships between fertility traits for dairy cows in different parities. Livest. Sci. 17, 337-349. [ Links ]
Kadarmideen, H.N., 2004. Genetic correlations among body condition score, somatic cell score, milk production, fertility, and conformation traits in dairy cows. J. Anim. Sci. 79, 191 -201. [ Links ]
Makgahlela, M.L., Banga, C.B., Norris, D., Dzama, K., Ng'ambi, J.W., 2007. Genetic correlations between female fertility and production traits in South African Holstein cattle. S. Afr. J. Anim. Sci., 37(3), 180-188. [ Links ]
Makgahlela, M.L., Banga, C.B., Norris, D., Dzama, K., Ng'ambi, J.W., 2008. Genetic analysis of age at first calving and calving interval in South African Holstein cattle. Asian J. Anim. Vet. Adv. 3, 197-205. [ Links ]
Miglior, F., Muir, B.L., Van Doormaal, B.J., 2005. Selection indices in Holstein cattle of various countries. J. Dairy Sci. 88, 1255-1263. [ Links ]
Misztal, I., Tsuruta, S., Lourenco, D., Masuda, Y., Aguilar, I., Legarra, A., Vitezica, Z., 2016. Manual for blupf90 family of programs. University of Georgia. Retrieved from http://nce.ads.uga.edu/wiki/lib/exe/fetch.php?media=blupf90. [ Links ]
Mostert, B.E., van der Westhuizen, R.R., Theron, H.E., 2010. Calving interval genetic parameters and trends for dairy breeds in South Africa. S. Afr. J. Anim. Sci. 40, 156-162. [ Links ]
Niozas, G., Tsousis, G., Steinhöfel, I., Brozos, C., Römer, A., Wiedemann, S., Bollwein, H., Kaske, M., 2019. Extended lactation in high-yielding dairy cows. I. Effects on reproductive measurements. J. Dairy Sci. 102, 1, 799-810. [ Links ]
Pryce, J.E., Royal, M.D., Garnsworthy, P.C., Mao, I.L., 2004. Fertility in the high-producing dairy cow. Livest. Prod. Sci. 86, 125-135. [ Links ]
Ramatsoma, N.I., Banga, C.B., Mackneil, M.D., Maiwashe, A., 2014. Evaluation of genetic trends for traits of economic importance in South African Holstein cattle. S. Afr. J. Anim. Sci. 44(1), 85-89. [ Links ]
Raheja, K.L., Burnside, E.B., Schaeffer, L.R., 1989. Heifer fertility and its relationship with cow fertility and production traits in Holstein dairy cattle. J. Dairy Sci. 72, 2665-2669. doi.org/10.3168/jds.S0022-0302.89.79407-8. [ Links ]
R Core Team. 2017. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/ [ Links ]
Sahin, A., Ulutas, Z., Adkinson, A.Y., Adkinson, R.W., 2014. Genetic parameters of first 80 Salem and Hammoud lactation milk yield and fertility traits in Brown Swiss cattle. Ann. Anim. Sci. 14, 545. [ Links ]
Shortle, W., 2014. Fertility in dairy cattle: a review. www.sementaicscholar.org. [ Links ]
Toghiani, S., 2012. Genetic relationships between production traits and reproductive performance in Holstein dairy cows. Arch Tierz. 55(5), 458-468. [ Links ]
Van Arendonk, J. A. M., Hovenier, R., De Boer, W., 1989. Phenotypic and genetic association between fertility and production in dairy cows. Livest. Prod. Sci. 21, 1 -12. [ Links ]
Wathes, D.C., Fenwick, M., Cheng, Z., Bourne, N., Llewellyn, S., Morris, D.G., Kenny, D., Murphy, J., Fitzpatrick, R., 2007. Influence of negative energy balance on cyclicity and fertility in the high producing dairy cow. Theriogenology. 68, S232-S241. [ Links ]
Wickham, H., 2016. ggplot2: Elegant Graphics for Data Analysis. Springer-Verlag, New York. [ Links ]
Wickham, H., François, R., Henry, L., Müller, K., 2018. dplyr: A Grammar of Data Manipulation. R package version 0.7.6. https://CRAN.R-project.org/package=dplyr. [ Links ]
Submitted 4 October 2021
Accepted 25 January 2023
Published 26 September 2023
# Corresponding author: kgarir@arc.agric.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}