










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 n.3 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i3.11
Pre-treatments of Mirasolia diversifolia using Lactobacillus bulgaricus at different dosages and fermentation times: Phytic acid concentration, enzyme activity, and fermentation characteristics
R. PazlaI, #; E.M. PutriII; N. JamarunI; W. NegaraIII; F. A. KhanII; M. ZainI; A. AriefI; G. YantiI; A. AntoniusIII; T. P. PriyatnoIII; M. SurachmanIII; I. W. A. DarmawanIII; H. HerdisIII; L. MarlinaIV; S. AsmairicenIII; Y. MartaV
IDepartment of Animal Nutrition, Faculty of Animal Science, Andalas University, Jl. Unand Limau Manis Campus, Padang 25163, West Sumatra, Indonesia
IIPost-doctoral Researcher, Research Center for Animal Husbandry, Research Organization for Agriculture and Food, National Research and Innovation Agency (BRIN) Indonesia, Jl. Raya Jakarta-Bogor, Cibinong 16915, Indonesia
IIIResearch Center for Animal Husbandry, Research Organization for Agriculture and Food, National Research and Innovation Agency (BRIN), Jl. Raya Jakarta-Bogor Cibinong 16915, Indonesia
IVResearch Center for Agroindustry, Research Organization for Agriculture and Food, National Research and Innovation Agency (BRIN). Jl. Puspitek Tangerang Selatan 15314, Indonesia
VCenter for Breeding Superior Livestock and Forage for Animal Feed Padang Mengatas, Jl. Raya Payakumbuh-Lintau, KM.9 Pekan Sabtu, Payakumbuh 26201, Indonesia
ABSTRACT
The present study aimed to determine the optimal dosage and fermentation time of Mirasolia diversifolia (leaves and stems) using Lactobacillus bulgaricus bacteria by observing the reduction in phytic acid, the activity of enzymes, pH of fermentation, number of bacterial colonies, and tannin concentration. An experiment was conducted using a completely randomized design (CRD) and 2x3 factorial design with three replications. The treatment factors were Factor A (L. bulgaricus dosages): A1 = 2% (g/v), A2 = 3% (g/v); and Factor B (fermentation times): B1 = 1 d, B2 = 3 d, B3 = 5 d. The dosage of L. bulgaricus and fermentation time had an interaction on phytic acid and its degradation, enzyme activities, pH fermentation, and the number of bacterial colonies. Tannin concentration was not affected. Mirasolia diversifolia fermented with 3% of L. bulgaricus for 5 d resulted in the lowest phytic acid content. Future research requires evaluating in vitro and in vivo dietary formulations for cattle using M. diversifolia fermented with L. bulgaricus.
Keywords: dosage and fermentation time, enzyme activity, fermentation characteristics, Lactobacillus bulgaricus, Mirasolia diversifolia, phytic acid
Introduction
The utilization of Mirasolia diversifolia as forage on a small- or large-scale tropical or subtropical farm has been widely used (Odedire & Oloidi, 2014; Ribeiro et al., 2016; Garcia et al., 2017; Mauricio et al., 2017; Jamarun et al., 2020). Those previous studies showed that M. diversifolia could increase the production of cattle and dairy cattle effectively. The high content of phytic acid in M. diversifolia is able to chelate minerals and proteins that form a complex of insoluble compounds, causing a decrease in the availability of these minerals and proteins (Gupta et al., 2015). Fasuyi et al. (2010) reported that M. diversifolia contains phytic acid at 79.1 mg/100g. Phytic acid aids the storage of phosphorus in plants and feed ingredients (Denstadli et al., 2010) in the form of myo-inositol-hexakisphosphate (Silva et al., 2021). Phytic acid in M. diversifolia confers a bitter taste, so it has low palatability for ruminants. Mirasolia diversifolia contains tannins, saponins, oxalates, alkaloids, and flavonoids that can form complexes with nutrients that are not easily degraded and absorbed in the gastrointestinal of ruminants (Aye, 2016). These anti-nutritional substances require treatment to decrease their concentration in M. diversifolia. Fermentation technology involving microorganisms can increase nutritional value, preserve feed ingredients, and decrease anti-nutritional or toxic substances contained in feed ingredients (Jamarun et al., 2017; Pazla et al., 2021a).
The microbe that can be utilized for fermentation of M. diversifolia is Lactobacillus bulgaricus, a type of lactic acid bacteria that is capable of producing phytase enzymes (Myo-inositol-hexakisphosphate) to hydrolyse phytic acid into inorganic monophosphate so that the bound nutrients can be utilized (Balwani et al., 2017; Handa et al., 2020). In addition, Hashemi et al. (2018) explained that lactic acid bacterial strains have the ability to synthesize phytase. Phytase is able to hydrolyse phosphomonoester bonds of phytate into myo-inositol and phosphoric acid via penta- to monophosphate. This leads to the decrease in anti-nutritional factors (phytic acid) and higher mineral bioavailability from fermented products. De Pasquale et al. (2020) reported a decrease in phytic acid and phytase activity in Leguminosae fermented with Lactobacillus sp. There are various benefits to utilising microbes as enzyme producers, including inexpensive production costs, rapid growth rates, and simple control (Astuti et al., 2022). Besides that, tannin concentration and trypsin inhibitor activity also decrease after fermentation.
Dosage and time of fermentation are also factors that must be considered in the process of fermented feed. Inappropriate dosage and restricted fermentation affect limited opportunities for microorganisms to grow. Substrate components that can be broken down into cell mass will also be small. It is crucial to determine the proper dosage and fermentation time so that fermentative bacteria have a substantial opportunity to grow and reproduce. These abilities will affect the quality of feed ingredients physically, chemically, and biologically. Furthermore, this study is a subsequent study of Pazla et al. (2021b), who reported that the fifth day of M. diversifolia leaves fermented with L. bulgaricus produced the lowest level of phytic acid, but without considering the dosage of L. bulgaricus. The current study aimed to determine the optimal dosage and fermentation time of M. diversifolia (leaves and stems) by evaluating dosages and duration of fermentation using L. bulgaricus while observing the reduction in phytic acid content, activity of enzymes, pH value of fermentation, number of bacterial colonies, and tannin concentration.
Materials and Methods
The present study did not use any live animals because it was conducted on a laboratory scale, so ethical approval was not required. This study was conducted at the Ruminant Laboratory and Feed Industry Laboratory, Faculty of Animal Science, Andalas University from April to July, 2022. Lactobacillus bulgaricus culture in the form of agar slant was obtained from the Livestock Technology Laboratory, Faculty of Animal Husbandry, IPB University, Indonesia. Mirasolia diversifolia was harvested from Tanah Datar Regency, West Sumatera. The geographical position of harvested area was 0°17'39'' S and 100°19'51'' E at an elevation of 750-1000m above sea level. The average rainfall is 1750-2000mm/year. The experimental design used in this study was a completely randomized design (CRD) in a 2* 3 factorial pattern with three replications. The treatment factors were: Factor A (L. bulgaricus dosage): A1 = 2% (g/v), A2 = 3% (g/v). Factor B (fermentation time): B1 = 1 d, B2 = 3 d, B3 = 5 d.
Mirasolia diversifolia was dried at 60 oC for 24 h in a forced-air oven and milled using a 1-mm sieve. The nutrient composition of M. diversifolia was determined using proximate analysis (Table 1).The sample was weighed in plastic as 100 g for 18 experimental units, mixed with 160 ml of distilled water, and then homogenized. The mixture was sterilized in an autoclave at 120 oC for 30 min. Broodstock of L. bulgaricus was made by dissolving 5.22 g MRS broth with 100 ml distilled water. After homogenisation, the broodstock was sterilized in an autoclave at 121 oC for 15 min. The L. bulgaricus culture from MRS agar was inoculated in the broodstock. After 48 h, 2 ml, and 3 ml of L. bulgaricus were inoculated into experimental diets. All samples were fermented under anaerobic conditions at room temperature based on the treatment for one, three, and five days.
The phytic acid concentration was determined following the method of Davies & Nightingale (1975). One gram of sample was homogenised with 50 ml of 0.5 M HNO3 using a shaker for 2 h, then strained. The determination of phytic acid was done by analysing the filtrate. An amount of 0.5 ml of filtrate was mixed with 0.9 ml of 0.5 N HNO3 and 1 ml of FeCb (containing iron ions; 50 μg/ml), covered with aluminium foil, and then kept in boiling water for 20 min. After cooling, 5 ml of amyl alcohol and 1 ml of ammonium thiocyanate (10%) were added to each tube. Then tubes were centrifuged for 10 minutes at 12298xg and absorbance was measured at 465 nm using an ultraviolet (UV)-visible (Shimadzu) spectrophotometer.
An amount of 6.9 g of Na2HPO4 (A solvent) was dissolved in 250 ml of distilled water; 13.41 g of Na2HPO4 (B solvent) was dissolved in 250 ml of distilled water. Then 39 ml of A solvent was mixed with 61 ml of B solvent and 100 ml of distilled water. The determination of enzyme activity was performed by preparing the enzyme extraction using 2 g of sample dissolved in 20 ml of phosphate buffer (pH 6.5 and 0.2 M) in an Erlenmeyer flask, which was then tightly covered with a rubber lid. The mixtures were homogenized using a shaker at 37 oC for 30 min. Then the mixtures were filtered and stored in the refrigerator for enzyme activity analysis.
Phytase enzyme activity was determined using the method of Kim & Lei (2005). An amount of 0.15 ml of enzyme extract was mixed with 0.6 ml of 0.1 M Tris HCL buffer (pH 7) containing Ca-phytate and CaCl2. Then, 0.75 ml of TCA 5% was added to the mixture. The mixture was incubated at 37 oC for 30 min. Then, 1.5 ml of molybdate colour reagent was added and homogenized using a vortex. The absorbance of the characteristic colour was measured at 700 nm against a blank using an ultraviolet (UV)-visible Shimadzu spectrophotometer. The results of the spectrophotometer measurements were regressed on the KH2PO4 standard curve. Each unit obtained was defined as the amount of enzyme that could cause the transformation of 1 μηι of substrate at optimal temperature (37 oC).
The Bergmeyer and Grassl method (Bergmeyer et al., 1974) was used to determine protease enzyme activity. An amount of 2.5 ml of casein, Hammarsten Grade, was mixed and homogenized with 1.5 ml of phosphate buffer (pH 6.5 and 0.2 M) in a test tube and incubated with distilled water at 37 oC for 10 min. Then, 1 ml of enzyme extract was added and incubated at 50 oC for 10 min. A 2 ml aliquot was mixed with 5 ml NaOH 0.5 M and 0.5 ml of Folin reagent. Absorbance was measured at 660 nm using an ultraviolet (UV)-visible (Shimadzu) spectrophotometer. The results of the spectrophotometer measurement were regressed on the bovine serum albumin standard curve. Each unit obtained was defined as the amount of enzyme that could cause the transformation of 1 μπι of substrate at optimal temperature (37 oC).
Cellulase enzyme activity was determined following the method of Nelson (1944) by mixing 0.5 grams of carboxymethyl cellulase (CMC) with 10 ml of phosphate buffer (pH 6.5 and 0.2 M) in each tube. An amount of 0.5 ml of this solution and 0.5 ml of enzyme extract were homogenized and soaked in distilled water at 40 oC for 20 min. Then, 1 ml of Nelson AB was added and boiled for 20 min. After cooling, 1 ml of phosphomolybdate was added and homogenized, followed by the addition of 7 ml of distilled water to each tube. Absorbance was measured at 575 nm using an ultraviolet (UV)-visible (Shimadzu) spectrophotometer. The results of the spectrophotometer measurement were regressed on the glucose standard curve. Each unit obtained was defined as the amount of enzyme that could cause a transformation of 1 μm of substrate at optimal temperature (37 oC).
Ten grams of the sample was dissolved in 30 ml of distilled water and then homogenized with a shaker for 1 h. The pH of fermentation was measured with a pH meter (Eutech Instruments, pH 700 device). The number of bacterial colonies was determined using the plant count method. A 10-g sample was dissolved in 90 ml of distilled water (101). One millilitre of broodstock liquor (101) was transferred to each of 18 test tubes (102) and then homogenized, and the process was continued until 109 dilutions. Then 1 ml of solution from 18 test tubes was pipetted (109) and transferred into a petri dish. An amount of 10 ml of MRS Agar (De Man, Rogosa, and Sharpe) medium was added and homogenized. The dishes were incubated at 37 oC for 24 h. The number of bacterial colonies was then counted.
Tannin concentration was measured using a UV-visible (Shimadzu) spectrophotometer. The sample was mixed with 150 ml of distilled water and incubated in the water bath at 70 oC for 30 min. The cooled extract was filtered and used for the reaction. The extraction results were measured with an ultraviolet (UV)-visible (Shimadzu) spectrophotometer at a wavelength of 278.5 nm using pure tannin (RnD Center Inc.) as standard.
Data obtained from this study was analysed using analysis of variance (ANOVA) according to a completely randomized design (CRD) factorial pattern. Differences between treatments (P <0.05) were tested using Duncan Multiple Range Test (DMRT). SPSS software was used to analyse the data (IBM SPSS Statistics, USA; version 21.0).
Results and Discussion
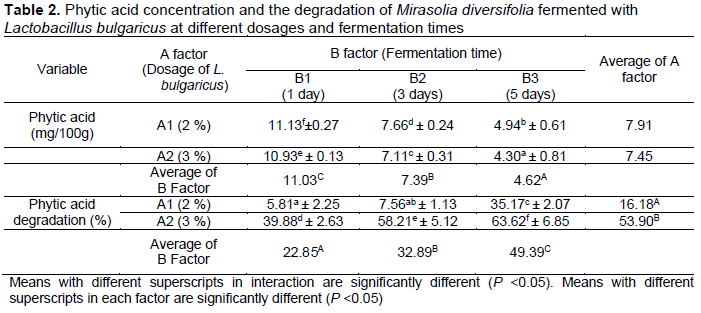
Statistical analysis showed that the inoculum dosage and fermentation time had an effect (P <0.05) on the phytic acid content and its degradation. From the interaction between inoculum dosage and fermentation time, the lowest phytic acid content was 4.30 mg/100g of M. diversifolia fermented with 3% L. bulgaricus for 5 days; the highest phytic acid degradation was 63.62%, observed in M. diversifolia fermented with 3% L. bulgaricus for 5 d (Table 2).
The present study showed a decrease in phytic acid concentration in M. diversifolia fermented with 2-3% of L. bulgaricus for 1-5 d. The decrease in phytic acid in M. diversifolia occurred due to the phytase enzyme produced by L. bulgaricus during the fermentation process. The phytase enzyme decreased linearly with the increase in fermentation time (Table 3). It was related to the decrease in the number of bacterial colonies. The decrease in bacterial colony number alters the production of the phytase enzyme. Phytase enzyme (myo-inositol-hexakisphosphate-3-phosphohydrolase) secreted by L. bulgaricus is an enzyme that is able to catalyse phytate (myo-inositol hexakisphosphate) into inorganic orthophosphate, releasing the phytate-phosphorus bond, and allowing phosphorus to be utilized by the ruminant (Selle & Ravindran, 2007; Pazla et al., 2021a). Phosphorus is associated with normal microbial rumen activity in degrading feed. Phosphorus is essential as material for protein synthesis in the body cell of the rumen microbes (Zain et al., 2010).
Anaemene & Fadupin (2022) also reported a decrease in phytic acid in pigeon pea fermentation due to the natural lactic acid activity. Natural lactic acid fermentation offers an optimal pH for the enzymatic degradation of phytates into organic phosphate and inositol. The optimum pH for L. bulgaricus to increase productivity is 5.5-6.2, in line with the studies of Rhee & Pack (1980) and Sneath et al. (1986). In contrast, Malaka & Laga (2005) stated that L. bulgaricus was still productive at pH 8.1. Phytic acid also decreases in rice bran fermented with L. bulgaricus, Streptococcus thermophilus, and L. acidophilus (Marsetyo et al., 2021). The phytic acid reduction may be due to the passive diffusion of water-soluble phytates (Bora, 2014). The phytic acid content in brown rice also decreased through the natural fermentation process by mixing brown rice and demineralized water (Liang et al., 2008).
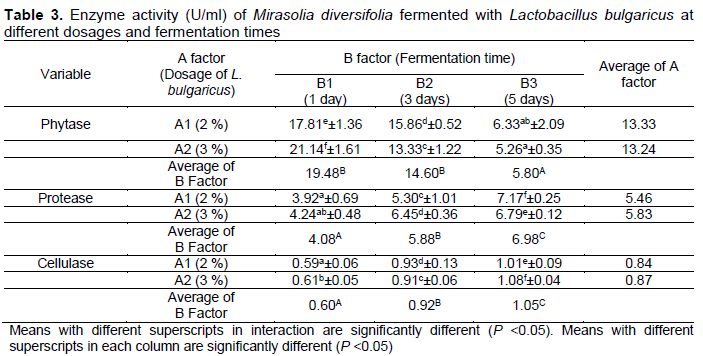
The statistical analysis showed that the inoculum dosage and fermentation time had an effect (P <0.05) on the enzyme activity (Table 3). From the interaction between the inoculum dosage and fermentation time, the highest phytase enzyme activity was 21.14 U/mL in M. diversifolia fermented with 3% L. bulgaricus for 1 d. The lowest activity of the phytase enzyme was 5.26 U/mL in M. diversifolia fermented with 3% inoculum for 5 d. The highest protease enzyme activity was observed in M. diversifolia fermented with 2% of L. bulgaricus for 3 d (7.17 U/ml). The lowest protease enzyme activity was observed in M. diversifolia fermented with 2% of L. bulgaricus for 1 d (3.92 U/ml). The highest cellulase enzyme activity was also observed in M. diversifolia fermented with 3% of L. bulgaricus for 5 d (1.08 U/ml); the lowest was observed in M. diversifolia fermented with 2% of L. bulgaricus for 1 d (0.59 U/ml).
Enzyme activity in fermentation is influenced by bacterial activity in producing enzymes. The substrates will be much more degraded by bacteria with a longer fermentation time. This concurs with Maftukhah & Abdullah (2018), who reported that enzyme activity in rice straw fermented with Aspergillus niger tended to increase at 4 d and decrease at 5-6 d of fermentation. Enzyme activity increases linearly with the increase of fermentation time and decreases when nutrients are depleted. In the present study, enzyme activity was optimal after 5 d of fermentation because nutrients were still available for microorganisms. In contrast, the phytase enzyme decreased with the increase in inoculum dosage and fermentation time. This indicated that more phytic acid had been degraded in 1-3 d of fermentation; the lowest phytic acid concentration was observed after 5 d of fermentation. In addition, the depression of the phytase enzyme was influenced by the number of bacterial colonies. A high number of bacterial colonies will produce more phytase enzyme, and conversely, a decrease in phytase enzyme would portend a depression in bacterial colony number (Thorsen et al., 2021).
In vitro protein digestibility increased by 92% and 47% in sorghum flours fermented with L. plantarum and natural fermentation, respectively (Pranoto et al., 2013). Protease enzyme produced by L. plantarum degrades the tannin-protein bond and complex proteins, thereby liberating more peptides and amino acids. Peptides and amino acids are materials needed by rumen microbes to synthesize the protein body (Zain et al., 2020; Putri et al., 2021).
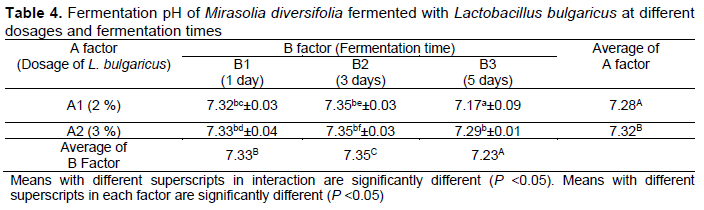
The range of pH fermentation in this study varied (Table 4). Statistical analysis showed that the inoculum dosage and fermentation time had an effect (P <0.05) on the pH value of fermentation. The interaction between inoculum dosage and fermentation time showed that the highest pH was 7.35 in M. diversifolia fermented with 2% and 3% L. bulgaricus for 3 d. The lowest pH value was 7.17 in M. diversifolia fermented with 2% of L. bulgaricus for 5 d.
The fermentation pH is associated with the growth of microorganisms. An acidic or alkaline pH can trigger microbial cell death. The high mortality rate of microorganisms will affect the speed of fermentation. The pH values obtained in this study were 7.17-7.35, and the increase in inoculum dosage and fermentation time depressed the pH value. The pH value still meets the requirements for the growth of L. bulgaricus bacteria. Mittal et al. (2011) reported that L. bulgaricus can grow at pHs of 5.5-7.5. Khota et al. (2016) confirmed that L. plantarum and L. casei had negative growth in a pH <3.5 in guinea grass and Napier grass silage. Enzymes produced by the bacteria can increase cell wall degradation and the availability of water-soluble carbohydrates for lactic acid bacteria, leading to a more rapid drop in pH.
The increase in inoculum dosage and fermentation time resulted in a reduced fermentation pH (Table 4). Generally, the fermentation process will decrease fermentation pH due to the action of lactic acid bacteria degrading organic matter into a simpler forms. The degradation of organic matter will produce valuable organic acids, such as formic, acetic, maleic, and succinic acid (Jeong & Lee, 2021). These organic acids will depress the pH of fermentation. This concurs with Puntillo et al. (2020), who observed that L. plantarum strains decreased in maize silage. The fermentation pH of hybrid maize harvested at 126 days of maturity and fermented with L. buchneri resulted in a decrease compared to the control silage (Pinto et al., 2020). Reale et al. (2007) also reported a decrease in fermentation pH in wheat, rye, and oats fermented with L. plantarum, L. amylovorus, and L. acidophilus. This is also related to the decrease in phytic acid content. The reduction in phytase enzyme activity will reduce phytate content resulting in a fall in pH during fermentation. In addition, the decrease of pH causes the decline in the number of bacterial colonies. The decrease in fermentation pH affected the decline in bacterial colony numbers with an increase in inoculum dosage and fermentation time (Table 4).
The number of bacterial colonies in this study differed among treatments (P <0.05; Table 5). With an interaction between inoculum dosage and fermentation time, the highest number of bacterial colonies was 184.67 CFU/mL in M. diversifolia fermented with 3% L. bulgaricus for 3 d. The lowest number of bacterial colonies was 56 CFU/mL in M. diversifolia fermented with 3% L. bulgaricus for 5 d.
The number of bacterial colonies decreased in line with the increase in both inoculum dosage and fermentation time. The lowest number of bacterial colonies was in M. diversifolia fermented with 2% L. bulgaricus for 5 days (Table 5). This decrease was due to low nutrient levels in M. diversifolia fermented for 5 d. Lactobacillus bulgaricus utilized all nutrients from days 1-4 such that the nutrients on the fifth day were available for degradation by the inoculum. The inoculum obtained optimal growth in 3 d. The number of bacterial colonies increased from days 1 -3 and then decreased after 5 d of fermentation (Table 5). This was also caused by the decrease in substrate after 5 d. Inadequate nutrient availability led to significant bacterial death after 5 d and supressed the bacterial population.
The number of bacterial colonies in this study was higher than the study of Pazla et al. (2021a) on M. diversifolia leaves fermented with L. bulgaricus, which was 8.5-31 CFU/mL. This difference was caused by the inoculum dosages and the fermentation substrate. Lactobacillus bulgaricus utilized the fermentable carbohydrate such as NFE (nitrogen free extract) that was available in fermented M. diversifolia. Pazla et al. (2023) reported that the fermentation of T. diversifolia would reduce the content of NFE because NFE is a fermentable energy source for microbial growth. Serial dilution and method or culture technique can also affect the results of the calculation of the number of bacterial colonies. Using serial dilution can reduce the density of bacterial colony growth in the sample.
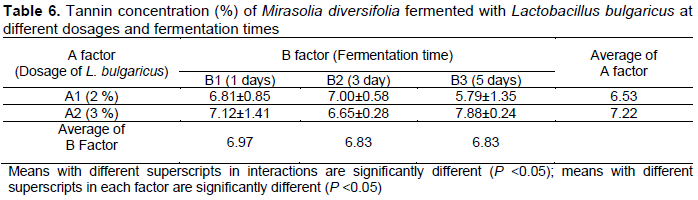
The present study showed that the inoculum dosage and fermentation time had no effect on the tannin concentration (P >0.05; Table 6). The dosage of L. bulgaricus and the duration of fermentation in the current study were not able to reduce tannin concentration in M. diversifolia. In contrast, Anaemene & Fadupin (2022) reported that fermentation reduced the tannin content of pigeon pea seeds by more than 80% and could be a result of the degradation of microbial enzymes released during fermentation.
A previous study also revealed that lactic acid bacterial fermentation could reduce the tannin content in Xuan Mugua, which is well known as a traditional Chinese medicine. Xuan Mugua consists of phenolics, tannins, triterpenes, glycosides, and organic acids; tannins and organic acids are particularly interesting. By incubation with lactic acid bacteria, the tannin concentration was reduced from 28% to 78% in Xuan Mugua (Shang et al., 2019). The present study reported that 2-3% of L. bulgarius and 1 -5 d fermentation was not effective in decreasing the tannin content of M. diversifolia. Factors that could affect the reduction in tannin concentration are the strain and dosage of inoculum, duration of fermentation, and the fermentable substrate.
Conclusion
The present study found that a dosage of 3% L. bulgaricus in M. diversifolia for 5 d of fermentation resulted in the lowest phytic acid content of 4.30 mg/100g with the highest degradation of 63.62%. Future research requires evaluating dietary formulations in vitro and in vivo for cattle using M. diversifolia fermented with L. bulgaricus.
Acknowledgments
The authors would like to thank the Research and Community Service Institute, Andalas University, which funded this research through an indexed publication research scheme with contract number: T/157/UN.16.17/PT.01.03/Pangan-RPT/2022. Thanks also to Laras Sukma Sucitra and the technicians of the Technology and Feed Industry Laboratory, Faculty of Animal Husbandry, Andalas University, and the technicians of the Livestock Technology Laboratory, Faculty of Animal Husbandry, IPB University.
Author contributions
RP, EMP, NJ, and MZ designed the concept, methodology, validation, investigation, writing - original draft, supervision, and project administration. WN, AA, TPP, MS, IWAD, HH, LM, SA, and YM designed methodology, formal analysis, investigation, resources, data curation, and writing-review. FAK, AA, and GY did the analysis, investigation, resources, data curation, and writing-original draft.
Conflict of Interests Declaration
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
Anaemene, D., & Fadupin, G., 2022. Anti-nutrient reduction and nutrient retention capacity of fermentation, germination, and combined germination-fermentation in legume processing. Appl. Food Res. 2(1), 2-6. DOI: 10.1016/j.afres.2022.100059 [ Links ]
Astuti, T., Akbar, S.A., Rofiq, M.N., Jamarun, N., Huda, N. & Fudholi, A., 2022. Activity of cellulase and ligninase enzymes in a local bioactivator from cattle and buffalo rumen contents. Biocatal. Agric. Biotechnol. 45(February), 2-7. DOI: 10.1016/j.bcab.2022.102497 [ Links ]
Aye, P.A., 2016. Comparative nutritive value of Moringa oleifera, Tithonia diversifolia, and Gmelina arborea leaf meals. Am. J. Food Nutr. 6(1), 23-32. DOI: 10.5251/ajfn.2016.6.1.23.32 [ Links ]
Balwani, I., Chakravarty, K., & Gaur, S., 2017. Role of phytase producing microorganisms towards agricultural sustainability. Biocatal. Agric. Biotechnol. 12(August), 23-29. DOI: 10.1016/j.bcab.2017.08.010 [ Links ]
Bergmeyer, H.U., Gawehn, K., & Grassl, M., 1974. Methods of Enzymatic Analysis. In: Bergmeyer, H.U., Ed., Verlag Chemie, Wienheim, Vol. 1, 481-482. [ Links ]
Bora, P., 2014. Anti-nutritional factors in foods and their effects. J. Acad. Industr. Res. 3(6), 285-290. [ Links ]
Davies, N.T. & Nightingale, R., 1975. The effects of phytate on intestinal absorption and secretion of zinc, and whole-body retention of Zn, copper, iron, and manganese in rats. Br. J. Nutr. 34(1965), 243-258. DOI: 10.1017/s0007114575000293 [ Links ]
De Pasquale, I., Pontonio, E., Gobbetti, M. & Rizzello, C.G., 2020. Nutritional and functional effects of the lactic acid bacteria fermentation on gelatinized legume flours. Int. J. Food Microbiol. 316, 1-37. DOI: 10.1016/j.ijfoodmicro.2019.108426 [ Links ]
Denstadli, V., Romarheim, A.T., S0rensen, M., Ahlstram & Skrede, A., 2010. Effects of dietary phytic acid on digestibility of main nutrients and mineral absorption in mink (Mustela vison). J. Anim. Feed Sci. 19(1), 104112. DOI: 10.22358/jafs/66274/2010 [ Links ]
Fasuyi, A., Dairo, F.A., & Ibitayo, F., 2010. Ensiling wild sunflower (Tithonia diversifolia) leaves with sugar cane molasses. Livest. Res. Rural Develop. 22(3). [ Links ]
Garcia, R., Gutierrez, D., Chongo, B., & Gutiérre, O., 2017. Feeding of cattle, sheep, and goats with Tithonia diversifolia in Latin America and the Caribbean. Mulberry, moringa, and tithonia in animal feed, and other uses. March, 237-250. [ Links ]
Gupta, R.K., Gangoliya, S.S. & Singh, N.K., 2015. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J. Food Sci. Technol. 52(2), 676-684. DOI: 10.1007/s13197-013-0978-y [ Links ]
Handa, V., Sharma, D., Kaur, A. & Arya, S.K., 2020. Biotechnological applications of microbial phytase and phytic acid in food and feed industries. Biocatal. Agric. Biotechnol. 25, 1-13. DOI: 10.1016/j.bcab.2020.101600 [ Links ]
Hashemi, S.M.B., Gholamhosseinpour, A. & Khaneghah, M.A., 2018. Fermentation of acorn dough by Lactobacilli strains: Phytic acid degradation and antioxidant activity. LWT-Food Sci. Technol. 1 -31. DOI: 10.1016/j.lwt.2018.10.054 [ Links ]
Jamarun, N., Pazla, R., Zain, M. & Arief, A., 2020. Milk quality of Etawa crossbred dairy goats fed combination of fermented oil palm fronds, Tithonia (Tithonia diversifolia) and Elephant Grass (Pennisetum Purpureum). J. Phys.: Confer. Ser. 1469(1), 1-8. DOI: 10.1088/1742-6596/1469/1/012004 [ Links ]
Jamarun, N., Zain, M., Arief & Pazla, R., 2017. Effects of calcium, phosphorus, and manganese supplementation during oil palm frond fermentation by Phanerochaete chrysosporium on lactase activity and in vitro digestibility. Pak. J. Nutr. 16(3), 119-124. DOI: 10.3923/pjn.2017.119.124 [ Links ]
Jeong, S., & Lee, J., 2021. Effects of sugars and degradation products derived from lignocellulosic biomass on maleic acid production. Energies, 14, 1-11. DOI: 10.3390/ en1404091. [ Links ]
Khota, W., Pholsen, S., Higgs, D. & Cai, Y., 2016. Natural lactic acid bacteria population of tropical grasses and their fermentation factor analysis of silage prepared with cellulase and inoculant. J. Dairy Sci. 99(12), 97689781. DOI: 10.3168/jds.2016-11180 [ Links ]
Kim, T.W. & Lei, X.G., 2005. An improved method for a rapid determination of phytase activity in animal feed. J. Anim. Sci. 83(5), 1062-1067. DOI: 10.2527/2005.8351062x [ Links ]
Liang, J., Han, B.Z., Nout, M.J.R. & Hamer, R.J., 2008. Effects of soaking, germination and fermentation on phytic acid, total, and in vitro soluble zinc in brown rice. Food Chem. 110(4), 821-828. DOI: 10.1016/j.foodchem.2008.02.064 [ Links ]
Maftukhah, S. & Abdullah, A., 2018. Cellulase enzyme production from rice straw using solid state fermentation and fungus, Aspergillus Niger ITBCC L74. MATEC Web of Confer. 156, 1-7. DOI: 10.1051/matecconf/201815601010 [ Links ]
Malaka, R. & Laga, A., 2005. Isolasi dan Identifikasi Lactobacillus bulgaricus strain ropy dari yoghurt komersial. Sains & Teknol. 5(1), 50-58. [ Links ]
Marsetyo, Sulendre, I.W., Mustaring & Pamulu, M., 2021. The effect of fermentation without or with lactic acid bacteria and storage time on the phytic acid, in vitro dry matter digestibility, and nutrient contents of rice bran. J. Phys.: Confer. Ser. 1763(1), 1-5. DOI: 10.1088/1742-6596/1763/1/012029 [ Links ]
Mauricio, R., Calsavara, L.H., Ribeiro, R.S., Pereira, L.G., Freitas, D.S., Domingos, S., Barahona, R., Rivera, J., Chará, J. & Murgueitio, E., 2017. Feeding ruminants using Tithonia diversifolia as forage. J. Dairy Vet. Anim. Res. 5(4), 117-120. DOI: 10.15406/jdvar.2017.05.00146 [ Links ]
Mittal, A., Singh, G., Goyal, V., Yadav, A., Aneja, K.R., Gautam, S.K. & Aggarwal, N.K., 2011. Isolation and biochemical characterization of acido-thermophilic extracellular phytase producing bacterial strain for potential application in poultry feed. Jundishapur J. Microbiol. 4(4), 273-282. [ Links ]
Nelson, N., 1944. A photometric adaptation of the Somogyi Method for the determination of glucose. J. Biol. Chem. 153(2), 375-380. DOI: 10.1016/s0021-9258(18)71980-7 [ Links ]
Odedire, J. & Oloidi, F., 2014. Feeding wild sunflower (Tithonia Diversifolia Hemsl., A. Gray) to West African Dwarf goats as a dry season forage supplement. World J. Agric. Res. 2(6), 280-284. DOI: 10.12691/wjar-2-6-6 [ Links ]
Pazla, R, Jamarun, N., Zain, M., Yanti, G. & Chandra, R., 2021a. Quality evaluation of tithonia (Tithonia diversifolia) with fermentation using Lactobacillus plantarum and Aspergillus ficuum at different incubation times. Biodiversitas. 22(9), 3936-3942. [ Links ]
Pazla, R., Yanti, G., Jamarun, N., Arief, Elihasridas & Sucitra, L.S., 2021b. Degradation of phytic acid from tithonia (Tithonia diversifolia) leaves using Lactobacillus bulgaricus at different fermentation times. Biodiversitas. 22(11), 4794-4798. DOI: 10.13057/biodiv/d221111 [ Links ]
Pazla, R., Jamarun, N., Yanti, G. & Putri, E. M., 2023. In vitro evaluation of feed quality of fermented Tithonia diversifolia with Lactobacillus bulgaricus and Persea americana Miller leaves as forages for goats. Trop. Anim. Sci. J., 46(1), 43-54. DOI: 10.5398/tasj.2023.46.1.43 [ Links ]
Pinto, S., Warth, J.F.G., Novinski, C.O. & Schmidt, P., 2020. Effects of natamycin and Lactobacillus buchneri on the fermentative process and aerobic stability of maize silage. J. Anim. Feed Sci. 29(1), 82-89. https://doi.org/10.22358/JAFS/118179/2020 [ Links ]
Pranoto, Y., Anggrahini, S. & Efendi, Z. 2013. Effect of natural and Lactobacillus plantarum fermentation on in vitro protein and starch digestibilities of sorghum flour. Food Biosci. 2, 46-52. DOI: 10.1016/j.fbio.2013.04.001 [ Links ]
Puntillo, M., Gaggiotti, M., Oteiza, J.M., Binetti, A., Massera, A. & Vinderola, G., 2020. Potential of lactic acid bacteria isolated from different forages as silage inoculants for improving fermentation quality and aerobic stability. Front. Microbiol. 11(December), 1-17. DOI: 10.3389/fmicb.2020.586716 [ Links ]
Putri, E.M., Zain, M., Warly, L. & Hermon, H., 2021. Effects of rumen-degradable-to-undegradable protein ratio in ruminant diet on in vitro digestibility, rumen fermentation, and microbial protein synthesis. Vet. World. 14(3), 640-648. DOI: 10.14202/VETWORLD.2021.640-648 [ Links ]
Reale, A., Konietzny, U., Coppola, R., Sorrentino, E. & Greiner, R., 2007. The importance of lactic acid bacteria for phytate degradation during cereal dough fermentation. J. Agric. Food Chem. 55(8), 2993-2997. DOI: 10.1021/jf063507n [ Links ]
Rhee, S. K., & Pack, M. Y., 1980. Effect of environmental pH on fermentation balance of Lactobacillus bulgaricus. J. Bacteriol. 144(1), 217-221. DOI: 10.1128/jb.144.1.217-221.1980 [ Links ]
Ribeiro, R.S., Terry, S.A., Sacramento, J.P., Silveira, S.R.E., Bento, C.B.P., Da Silva, E.F., Mantovani, H.C., Da Gama, M.A.S., Pereira, L.G.R., Tomich, T.R., Maurício, R.M. & Chaves, A.V., 2016. Tithonia diversifolia as a supplementary feed for dairy cows. PLoS ONE. 11(12), 1-18. DOI: 10.1371/journal.pone.0165751 [ Links ]
Selle, P.H. & Ravindran, V., 2007. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 135(1-2), 1-41. DOI: 10.1016/j.anifeedsci.2006.06.010 [ Links ]
Silva, V.M., Putti, F.F., White, P.J. & Reis, A. R. dos. 2021. Phytic acid accumulation in plants: Biosynthesis, pathway regulation, and role in the human diet. Plant Physiol. Biochem. 164, 132-146. DOI: 10.1016/j.plaphy.2021.04.035 [ Links ]
Shang, Y.F., Cao, H., Ma, Y.L., Zhang, C., Ma, F., Wang, C.X., Ni, X.L., Lee, W.J. & Wei, Z.J., 2019. Effect of lactic acid bacteria fermentation on tannins removal in Xuan Mugua fruits. Food Chem. 274(August), 118-122. DOI: 10.1016/j.foodchem.2018.08.120 [ Links ]
Sneath, P.H.A., Mair, N.S., Sharpe, M.E. & Holt, J.G., 1986. Bergey's Manual of Systematic Bacteriology. 2nd ed. Williams and Wilkins, Baltimore. [ Links ]
Thorsen, M., Nielsen, L. A., Zhai, H. X., Zhang, Q., Wulf-Andersen, L. & Skov, L. K., 2021. Safety and efficacy profile of a phytase produced by fermentation and used as a feed additive. Heliyon 7(6), 1-12. DOI: 10.1016/j.heliyon.2021.e07237 [ Links ]
Zain, M., Jamarun, N. & Zulkarnaini, N., 2010. Effect of phosphorus and sulphur supplementation in growing beef cattle diet based on rice straw ammoniated. Asian J. Sci. Res. 3(3), 184-188. DOI: 10.3923/ajsr.2010.184.188 [ Links ]
Zain, M., Putri, E.M., Rusmana, W.S.N., Erpomen & Makmur, M., 2020. Effects of supplementing Gliricidia sepium on ration based on ammoniated rice straw in ruminant feed to decrease methane gas production and to improve nutrient digestibility (in vitro). Int. J. Adv. Sci. Eng. Inform. Technol. 10(2), 724-729. DOI: 10.18517/ijaseit.10.2.11242 [ Links ]
Submitted 12 January 2023
Accepted 2 April 2023
Published 23 July 2023
# Corresponding author: ronipazla@ansci.unand.ac.id


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}