










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 n.3 Pretoria 2023
http://dx.doi.org/10.4314/sajas.v53i3.01
Estimating milk production and energy-use efficiency of pasture-grazed Holstein and Jersey cows using mathematical models
N.M. BanganiI, III, #; C.J.C. MullerI, II; K. DzamaI; C.W.C CruywagenI; F.V. Nherera-ChokudaIII; V.E. Imbayarwo-ChikosiIV
IDepartment of Animal Sciences, Stellenbosch University, Stellenbosch 7600
IIDirectorate Animal Sciences, Private Bag X1, Elsenburg 7607
IIIAnimal Production Institute, Agricultural Research Council, Irene
IVDepartment of Animal Science, University of Eswatini, Luyengo Campus
ABSTRACT
The efficiency of feed use for milk production is critical for sustainable and profitable pasture-based dairy systems. The aim of this study was to estimate milk production and energy-use efficiencies of pasture-grazed Holstein and Jersey cows. Lactation records of 122 Holstein and 99 Jersey cows varying from parities 1 to 6 that were managed under similar feeding and environmental conditions were collected from 2005 to 2014. Feed intake and nutrient requirements of the cows were calculated using the National Research Council and the Cornell Net Carbohydrate and Protein System equations. Holsteins had a higher milk yield/kg dry matter intake (1.36±0.01 vs. 1.27±0.01 kg), whereas Jerseys had higher efficiencies in milk fat (52.4±0.3 vs. 58.4±0.4 g), milk protein (42.7±0.3 vs. 45.1±0.3 g), and energy-corrected milk (1.30±0.01 vs. 1.36±0.01 kg) per kg dry matter intake. Jersey cows also had a higher dry matter intake/kg body weight (3.13±0.02 vs. 3.51±0.02%). During transition and early lactation stages, Holstein and Jersey cows were in negative energy balance for 102.4±2.3 vs. 74.2±2.3 days, with the lowest energy reserves (-53.9 MJ vs. -39.7 MJ) reached at 22.3±0.9 vs. 24.6±0.9 days post-calving, respectively. Compared to Holsteins, Jersey cows used proportionally less net energy intake to produce 100 g milk fat (13.7±0.10 vs. 12.5±0.10), 100 g milk protein (16.7±0.14 vs.16.2±0.15) and a 1-kg energy-corrected milk (5.52±0.04 vs. 5.35±0.04), making them a better breed for pasture-based dairy systems as they possess more production and feed-use efficiency traits, which are desirable in pasture-based production systems.
Keywords: energy balance, energy corrected milk, energy intake, feeding and environment
Introduction
Feed efficiency is a biological traits that is referred to as a trait of economic importance on a dairy farm. This is because, although the capacity to secrete milk is determined by the metabolic ability of the mammary tissues, maximum rates of milk synthesis are largely influenced by the continuous supply of nutrients, their digestion, and conversion efficiency for synthesis of the precursors for supply to the mammary tissue (Boyd & Kensinger, 1998; Cai et al., 2018). This therefore makes the efficiency with which feed is converted to milk a critical element for farm profitability and sustainability. A cow with a high feed-use efficiency presents with higher feed intake per unit liveweight, has lower maintenance energy requirements, partitions more metabolizable energy to milk than to body tissue, and loses less energy in faeces, urine, or methane for a given intake (Grainger & Goddard, 2004). The weight loss should, however, be on a short-term basis as long-term weight loss may predispose the cow to metabolic disorders and poor reproductive performance.
For milk synthesis, energy is the most essential nutrient, being the major determinant of milk volume as it is responsible for regulating osmotic pressure in the mammary system (Liu et al., 2013; Lin et al., 2016), is a precursor of milk fat (Gorewit, 1988; Rezaei et al., 2016), and is a supplier of energetic precursors for protein synthesis to manufacture milk protein (Mepham, 1982; Bionaz et al., 2012). Despite being essential, energy is often the most limiting nutrient (VandeHaar et al., 2016) as the recommended inclusion rate of non-fibre carbohydrates in lactating dairy cows' diet is 30-45% on a dry matter basis (Batajoo & Shaver, 1994; Afzalzadeh et al., 2010; Hall et al., 2010). The energy deficiency effects become more pronounced in grazing animals as the bulk of pasture grazed consists of cellulose. Approximately 20-70% of cellulose may not be digestible, resulting in only 10-35% of energy intake being captured as net energy (Varga et al., 1997). Consequently, there is less available energy for maintenance and production functions such as milk production, growth, and pregnancy. Moreover, the excess fibre in the diet of grazing animals results in longer physical fill of the rumen due to prolonged feed retention time and hence reduces intake to below the required levels. The efficiency in digesting fibre and partitioning the available net energy to maintenance and production functions is therefore of significance in cows on pasture.
In South Africa, milk prices are determined by milk processors, based mainly on specific amounts of fat and protein (Anonymous, 2017). The two components have a major effect on the quality and quantity of dairy products produced. Holsteins and Jerseys are the dominant dairy breeds in commercial herds; their differences in production and feed-use efficiency may have economic consequences and requires investigation. Several studies that compare the two breeds have been conducted (Mackle et al., 1996; Muller & Botha, 1998; Rastani et al., 2001; Thomson et al., 2001; Anderson et al., 2007; Aikman et al., 2008; Prendiville et al., 2009; Kristensen et al., 2015; Olijhoek et al., 2018). Breed variability in performance efficiencies between countries has been observed, indicating that research that compares the performance efficiencies of Holstein and Jersey cows from different countries should not be applied directly to another country due to differences in production systems, available feeds, and climatic conditions. Moreover, the duration of the conducted studies ranges from one season of the year to one lactation period or a year. A longitudinal study that will provide a repeated measure of milk production and feed-use efficiency on the same subjects at different lactation stages in different parities will provide information on the consistency and persistency of these variations in cows and therefore substantiate the available literature. The aim of this study was to estimate and compare energy use and milk production efficiencies of pasture-grazed Holstein and Jersey cows in a Mediterranean-type climatic region.
Materials and methods
Details of experimental animals, experimental area, diet, and management of experimental animals were presented in Bangani et al. (2022); only a summary will be provided in this paper. Lactation records of 122 Holstein and 99 Jersey cows that were managed under similar feeding and environmental conditions were used. The records were compiled as part of the National Milk Recording and Improvement Scheme under the Animal Production Institute of the Agricultural Research Council to estimate breeding values for sires, cows, and heifers for a genetic profile of individual herds. Collected records included cow birth date, calving date, lactation number, kg body weight (BW), kg milk yield (MY), % milk fat (MF), and % milk protein (MP). Milk was corrected for its fat and protein content to energy-corrected milk (ECM) using the equation of Tyrell & Reid (1965) (Table 1). The use of data was approved by the Information Officer for the Supply of Biological Specimens and other Data (Ref. No. 2015/001), Directorate: Animal Sciences, Western Cape Department of Agriculture, South Africa. Mathematical models from the National Research Council (NRC, 2001) and the Cornell Net Carbohydrate and Protein System (CNCPS) were used to predict feed intake, nutrient composition, and animal requirements from the secondary data (Table 1). Both the NRC and CNCPS models use equations from peer reviewed scientific articles (Fox et al., 2004; Tedeschi et al., 2014), making them suitable to use in this study.
The dry matter intake (DMI) for each cow was estimated using the NRC (2001) equation (Table 1). It was calculated as the sum of the concentrate mixture and pasture intake. The concentrate mixture was offered at 7 kg/day and contained 170 g/kg crude protein on an as-fed basis. Individual cow pasture intake could not be directly measured as cows from both breeds grazed as one herd. It was therefore calculated as the difference between the estimated DMI and concentrate offered. The CP content of the pasture averaged 184 g/kg dry matter. Although the NRC (2001) formula was developed for Holstein cows, it was also used to estimate the DMI of Jersey cows in this study. This is because the NRC (2001) formula uses predictor variables that influence feed intake which apply to both breeds, e.g., body weight, lactation stage, and milk production, which are corrected to account for the difference in milk yield and composition. The efficiency of DMI use was estimated as kg MY/kg DMI, g MF/kg DMI, g MP/kg DMI, kg ECM/kg DMI, and kg DMI/kg BW.
For estimating the energy content of feed, the net energy (NE) contents of ingredients were obtained from the feed formulation software package, Nutritional Dynamic System (NDS) Professional (NDS version 6.5, 2008 to 2018). The software package uses the CNCPS biological model as a formulation and evaluation platform (NDS Professional, version 6.5, 2008 to 2018). The ingredients and their NE values in megajoules (MJ)/kg were: wheaten bran (6.78), barley (8.37), maize (8.52), cottonseed oilcake meal (7.18), soybean oilcake meal (9.47), fishmeal (9.16), urea (0), molasses (7.30), wheat straw (1.54), limestone (0), and salt (0). The proportions of NE contributions from feed ingredients were summed up, making an average NE content of 7.81 MJ/kg for the concentrate. For the kikuyu over-sown with annual ryegrass, the metabolizable energy (ME) value used (9.43 MJ ME/kg DM) was obtained from Botha et al. (2008). Because approximately one-third of ME is lost as heat during the fermentation, digestion, and metabolism of nutrients (VandeHaar, 2011), pasture NE was calculated as two thirds of its ME, i.e., 6.28 MJ/kg. The net energy intake (NEI) was calculated as the sum of NE contributions from the concentrate mixture offered and estimated pasture intake.
The energy requirements of cows, i.e., net energy for maintenance (NEm), energy for lactation (NEL), and energy for growth (NEg) were calculated using the equations from the CNCPS and NRC (2001) (Table 1). Energy balance (EB) was calculated as (NEI - (NEm + NEl + NEg)). Although the ME for pregnancy is part of EB, it was not included in the equation as the area of interest was body mobilisation of energy reserves, i.e., transition and early lactation stages. Cows whose estimated energy demands exceeded NEI were declared to be in a negative energy balance (NEB). To determine the number of days the cow was in NEB, the days in milk (DIM) in which the first positive EB was recorded was used to represent the duration of NEB. For NEB magnitude, the NEB nadir was defined as the lowest NEB point achieved by the cow. The number of days to reach NEB nadir was the DIM in which the lowest NEB value was recorded. The efficiency of NE use was calculated as NEI/100 g MF, NEI/100 g MP, NEI/kg ECM, and NEI/kg BW075.
Data were analysed using the repeated measures ANOVA in the PROC MIXED procedure of Statistical Analysis System (SAS) Enterprise Guide, version 7.1. A compound symmetry structure for the residuals was used as a covariance structure for repeated measures over time within cows. The equation that was used for statistical analysis was as follows:
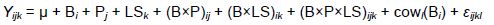
The response variables (Ya) were the milk yield and its components. The effect of the cow was fitted as a random effect and nested within the breed (Cow/(B,)). The fixed effects were breed (B,), parity (Pj), lactation stage (LSk), and their interactions. The between-breeds, between-parity, and between-lactation stage variations and their interactions were compared using the Bonferroni test and were declared different at P <0.05.
Results and discussion
Cows in parities 4 and beyond were grouped into one group of parity four and higher (parity 4+) for each breed (Bangani et al., 2022). Detailed results on milk, its components, and DMI are presented in Bangani et al. (2022). Below are the descriptive statistics for the data used (Table 2).
In all parities and lactation stages, the efficiency of converting DMI to MY was higher in Holstein than Jersey cows (Figure 1, Table 3). These results concur with what has been reported in previous studies, e.g., in primiparous cows in South Africa that were on TMR (Muller & Botha, 1998); and in two groups of cows in New Zealand that were either on ad libitum pasture, or restricted pasture plus a concentrate (Thomson et al., 2001). This indicates that although genetic improvement in MY traits for both Holstein and Jersey cows has been achieved over time, the difference in MY production efficiency between the two breeds remains unchanged. The kg MY/kg DMI for first and second lactation did not differ in Jersey cows (P >0.05); an increase was observed in the third lactation, with third and subsequent lactations being similar. In Holstein cows, MY/kg DMI increased from first to third lactation, with efficiency being similar in third and parity 4+ cows (Figure 1, Table 3).
In both breeds, however, a downward trend was observed in kg MY/kg DMI as lactation stages advanced (Figure 1), suggestive of homeorhetic regulation, i.e., nutrient partitioning in dairy cows to prioritise a physiological need, which Bauman & Currie (1980) define as the "orchestrated changes for the priorities of a physiological state". At the onset of lactation, the most critical role is the synthesis and secretion of high amount of milk; nutrient use is thus altered to prioritise this function (Bauman & Currie, 1980). This results in high milk production efficiency during transition and early lactation stages although it happens at the expense of body reserves. In mid- and late-lactation stages, nutrient partitioning shifts towards building body reserves and supporting pregnancy in preparation for the next calving, and therefore incurs a decrease in milk production efficiency. Strategic feeding of the cow to align with the lactation stage may be beneficial in improving her performance efficiency.
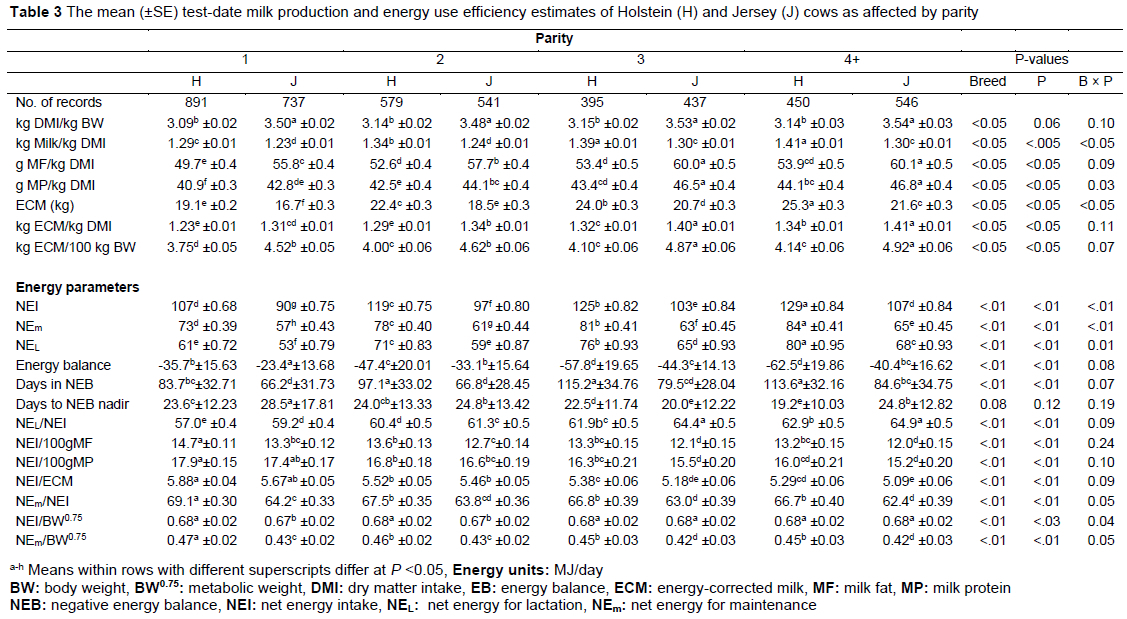
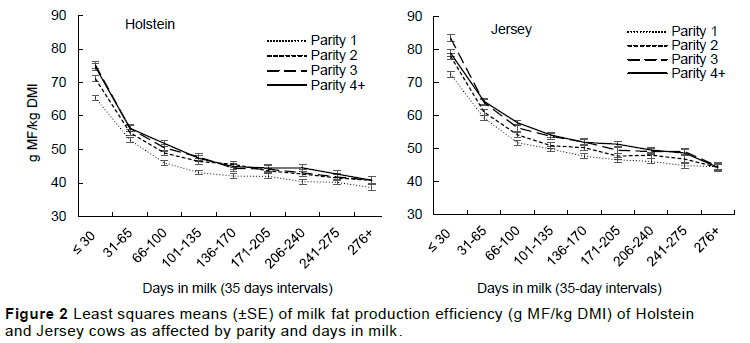
The efficiency of converting DMI to both MF and MP was higher in Jerseys compared to Holsteins in all production stages, i.e., parity and lactation stages (Table 3, Figures 2 and 3), with a mean test-day yield of 52.4±0.5 vs 58.4±0.4 g MF/kg DMI and 42.7±0.3 vs 45.1±0.3 g MP/kg DMI in Holsteins and Jerseys, respectively. Higher efficiency for MF/kg DMI in Jersey cows was reported by Mackle et al. (1996) on cows that grazed on pasture in New Zealand, and Thomson et al. (2001) in cows on either ad libitum pasture or restricted pasture, but these authors reported no breed effect in efficiency ratios for MP/kg DMI. Holstein MF/kg DMI was lower in the first parity compared to the subsequent parities, whose MF production efficiency did not differ. With Jerseys, MF/kg DMI increased gradually, reaching its peak in the third lactation, then levelled. In both breeds, MP/kg DMI increased up to third lactation; parities 3 and 4+ did not differ (Table 3). As lactation stages progressed, g MF/kg DMI and g MP/kg DMI decreased. This can be associated with the decreasing trend observed in MY with advancing lactation stages (Figure 2 and 3).
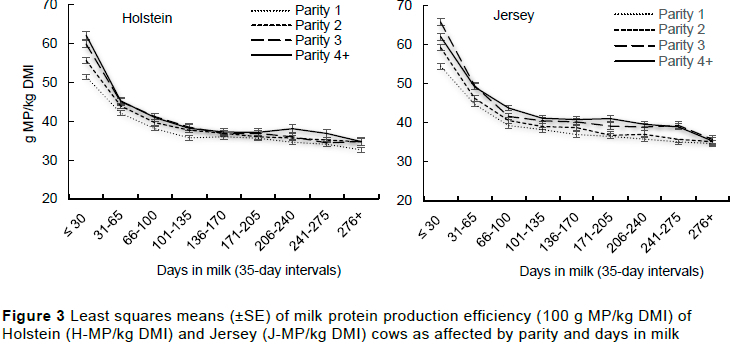
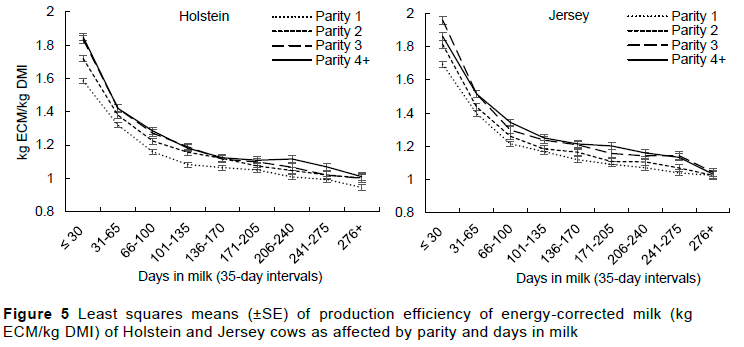
After correcting milk for its fat and protein content, the mean ECM was 22.7±5.9 kg/day in Holsteins and 19.4±4.8 kg/day in Jerseys. This indicates that Jersey cows produced on average 85.5% the ECM of Holsteins, a higher proportion compared to the average 74.1% MY reported by Bangani et al. (2022) in the same herd. This increase in proportion is attributable to the high solid component of the Jersey milk. The ECM increased with parity but decreased with lactation stage (Figure 4). Because of the increase in the amount of ECM produced, Jersey cows produced higher ECM/kg DMI and ECM/kg BW (Table 3) compared to Holsteins. The low DMI conversion efficiency to solid corrected milk in Holsteins compared to Jerseys was also reported by Mackle et al. (1996). Kristensen et al. (2015) also reported higher efficiency in Jerseys compared to Holsteins in cows that were in a total mixed or partial mixed ration for six months in Denmark. Olijhoek et al. (2018), however, reported no difference between the two breeds when they were allocated in two feeding levels, i.e., either high or low concentrate diets. Both ECM/kg DMI and ECM/kg BW increased with parity (Table 3) but decreased with lactation stage. This was expected as the milk production efficiency parameters (MY/kg DMI, g MF/kg DMI and g MP/kg DMI) all increased with parity and decreased with lactation stage.
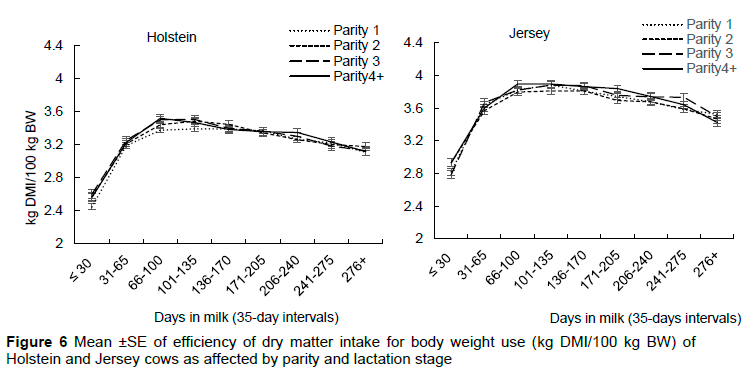
Holstein cows had a lower DMI/kg BW than Jersey cows (Table 3, Figure 6). In agreement, several authors reported lower DMI/kg BW in Holsteins compared to Jerseys that were on: TMR in South Africa (Muller & Botha, 1998); two feeding systems in New Zealand, i.e., either ad íib pasture or restricted pasture plus a concentrate (Thomson et al., 2001); TMR in the United States of America (Anderson et al., 2007); pasture in Ireland (Prendiville et al., 2009); or either a total mixed or partial mixed ration in Denmark (Kristensen et al., 2015). In contrast, these authors reported no difference in DMI/kg BW between Holstein and Jersey cows that were on: TMR in the United States of America (Rastani et al., 2001); TMR in the United Kingdom (Aikman et al., 2008); or TMR based on silage in the United States of America (Knowlton et al., 2010).
The high DMI/kg BW observed in Jersey cows in this study suggests that Jersey cows would be more suitable for pasture production systems in comparison to Holstein cows as energy in pasture is often limiting. It is suggestive of higher energy intake which may provide better energy reserves, thus preventing excessive lipolysis and the effects of negative energy balance, especially after calving. Moreover, cows that eat more often produce more as the excess food above maintenance is partitioned to production. Aikman et al. (2008) associated the higher DMI/kg BW in Jerseys with the higher passage rate of digesta in this breed compared to Holsteins. In agreement, Ingvartsen and Weisberg (1993), observed a 21% higher passage rate in Danish Jerseys compared to Holsteins. Retief (2000) and Bangani (2002) reported higher effective dry matter and neutral detergent fibre degradability in Jerseys compared to Holsteins at all fractional outflow rates, suggesting that with the higher DMI in this breed, extraction of nutrients from the digesta is also high. Parity had no effect on DMI/kg BW (Table 3, Figure 6), however, this parameter increased with lactation stage, reaching a peak in mid-lactation, thereby coinciding with the peak estimated DMI, and thereafter lowering in the late lactation stage (Figure 6). The lower DMI/kg BW in late lactation stage is attributable to a decrease in DMI with decreasing milk production, accompanied by an increase in BW as the cows regain their body condition and some may even be pregnant.
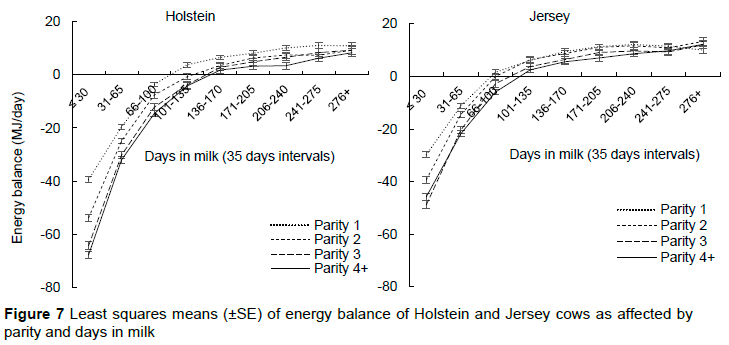
The overall mean estimated NEI was 120±0.7 vs. 99±0.7 MJ/day, whereas estimated net energy requirements for maintenance were 79±0.4 vs. 61±0.4 MJ/day; lactation, 72±0.7 vs. 61±0.7 MJ/day; and growth, 9.4±3.0 and 2.6±2.1 MJ/day in Holstein and Jersey cows, respectively. The estimated energy balance implied that cows were in a negative energy balance (NEB) state in transition and early lactation stages, achieving a positive energy balance in mid-lactation (Figure 7). This is in line with expectation as the cows are transitioning from a pregnant, non-lactating state to synthesising and producing large amounts of milk. The nutrient requirements of the cow rise rapidly for the initiation of milk synthesis after calving, followed by the high milk production, which peaks in early lactation while DMI is still lagging (Drackley et al., 2005).
To offset the energy deficiency resulting from producing large amounts of milk while DMI is low, dairy cows undergo increased levels of fat mobilisation, which involves lipolysis (Rodriguez et al., 2020).
During lipolysis, non-esterified fatty acids (NEFA) are released from adipose tissue into the bloodstream to be used as substrate for milk fat synthesis and as an energy source in tissues to compensate for the increased energy demands (Wathes et.al, 2007; Block, 2010; Rodriguez et al., 2020), resulting in tissue NEB.
The estimated NEB intensity was higher in Holstein cows, reaching nadir at -53.9±0.8 MJ compared to -39.7±0.8 MJ in Jersey cows (Figure 7). In agreement, Rastani et al. (2001) reported a tissue energy balance nadir of -6.19 Mcal/day (-26 MJ/day) in Jerseys and -12.9 Mcal/day (-54 MJ/day) in Holsteins. Friggens et al. (2007) also reported a less intense NEB in Jerseys compared to Holsteins that were fed either a normal or low-density energy TMR in Denmark, whereas Washburn et al. (2002), reported lower body condition scores (which can be seen as a proxy for NEB intensity) in Holsteins compared to Jerseys that were kept either on pasture or under intensive systems. Holsteins had a lower DMI/kg BW, which suggests a lower energy intake that may result in excessive depletion of body reserves to compensate for the insufficiency. The NEB intensity also increased with parity in both breeds. In agreement, Gallo et al. (1996) and Friggens et al. (2007) reported less marked depletion and faster recovery of body reserves in primiparous cows compared to multi-parous ones, whereas Lee & Kim (2006) observed an increase in loss (P <0.01) and delayed recovery of body condition with increase in parity (P <0.01) in cows that were on TMR. Macrae et al. (2019) and Walter et al. (2022) reported lower post-partum plasma NEFA concentrations in primiparous cows compared to multiparous ones. A higher plasma concentration of NEFA is indicative of excessive fat mobilisation (Tessari et al., 2020). Using plasma NEFA concentration as an index for lipid mobilisation may suggest that primiparous cows experience lower lipid mobilisation compared to multiparous ones, a possible reason for lower NEB intensity in primiparous cows.
The number of days it took to reach NEB nadir did not differ between breeds (P = 0.08); parity also had no effect (P = 0.12). The duration of NEB was, however, longer in Holsteins than Jerseys (Table 3, Figure 7). In agreement, Rastani et al. (2001) reported a tissue energy balance that occurred at week 1 of lactation and lasted for 7 weeks in Jerseys, whereas in Holsteins, it occurred at week 2 and endured for 11 weeks of lactation. In accord, Friggens et al. (2007) also reported a shorter NEB in Jerseys compared to Holsteins. The NEB duration increased with parity (P <0.01). With the higher NEB intensity observed in Holstein cows and multiparous cows, recovery was expected to be prolonged in these two groups, a possible reason for a longer NEB duration. Furthermore, the regenerative capacity of tissue in animals is known to decrease with age, hence the shorter NEB duration in primiparous cows compared to multi-parous ones.
The efficiency with which energy is used for lactation or milk production is a key driver of production efficiency (Xue et al., 2011) in dairy cows. Jersey cows allocated proportionally more of the estimated NEI to lactation (NEL/NEI) and lesser to maintenance (NEm/NEI) and liveweight NEm/BW0.75 compared to Holstein cows (Table 3). According to Grainger & Goddard (2004), an efficient cow partitions more metabolizable energy to milk and has lower maintenance energy requirements.
Attributable to the high solid component of milk produced by Jerseys, they also used proportionally less mean NEI to produce 100 g MF (13.7±0.10 vs. 12.5±0.10), 100 g MP (16.7±0.14 vs.16.2±0.15), and a kg ECM (5.52±0.04 vs. 5.35±0.04) compared to Holsteins (Table 3). In agreement, a higher efficiency of converting metabolizable energy intake to milk energy output was reported by Mackle et al. (1996), whereas Kristensen et al. (2015) reported a higher ECM/10 MJ of NEI in Jersey cows compared to Holstein cows. Using solid-corrected milk as a proxy for ECM, Blake et al. (1986), however, reported no difference in energy-use efficiency between the two breeds. In both breeds, the NEI used to produce 100 g MF, 100 g MP, and kg ECM also decreased with parity (Table 3), indicating higher efficiency with maturity. With lactation stages, the efficiency of energy use to produce 100g MF, 100 g MP, and kg ECM decreased by +50% from transition to late lactation stage, i.e., 10.0±0.17 to 16.4±0.12, 12.6±0.23 to 19.5±0.17, and 4.11 ±0.06 to 6.63 ±0.05 in Holsteins; and 9.3±0.17 to 15.0±0.12, 12.0±0.23 to 19.0±0.17, and 3.95 ±0.07 to 6.43±0.05 in Jerseys, respectively, indicative of shifting of nutrients from lactation towards building body reserves. Although the estimated NEI/kg BW0.75 was higher in primiparous and second lactation Holsteins than that of Jerseys (P <0.01), there was no breed effect in later parities (Table 3). Rastani et al. (2001) also reported no breed effect in NEI/kg BW075 (P = 0.89).
Conclusion
The difference in milk production efficiency between the two breeds was mainly as a result of the difference in milk yield and composition with Holstein cows showing a higher efficiency in milk yield while Jerseys had higher milk components per kg dry matter intake. The higher solid component of Jersey milk, which resulted in higher energy-corrected milk per kg of dry matter intake makes the Jersey cow a more production-efficient breed compared to Holstein cows. Jersey cows also possessed feed-use efficiency and energy balance traits that are desirable for a pasture-based system, e.g., higher dry matter intake per kg body weight and a shorter, less intense negative energy balance, making them a better breed for this production system. It was also evident in both breeds that production efficiency increased with parity and decreased with lactation stage, indicating the importance of a longer productive life and strategic feeding of the cow to align with the lactation stage for improved production efficiency.
Acknowledgement
The authors would like to acknowledge the Western Cape Department of Agriculture, Animal Production Division, Elsenburg for allowing them to use their data. This study was funded by the Western Cape Agricultural Research Trust (WCART) and the Agriculture Sector for Education and Training (AgriSETA).
Authors' Contributions
Drafting of paper: NMB; critical revision: CJCM, CWC, FN-C &VE I-C; final approval of version to be published: KD.
Conflict of Interest Declaration
The authors certify that they have no affiliations with any organization or entity with financial or non-financial interest in the subject matter or materials discussed in this manuscript.
References
Afzalzadeh, A., Rafiee, H., Khadem, A.A. & Asadi, A., 2010. Effects of ratios of non-fibre carbohydrates to rumen degradable protein in diets of Holstein cows: 1. Feed intake, digestibility, and milk production. S. Afr. J. Anim. Sci. 40 (3), 204-212. doi: 10.4314/sajas.v40i3.5. [ Links ]
Aikman, P. C., Reynolds, C. K., & Beever, D. E., 2008. diet digestibility, rate of passage, and eating and rumination behaviour of Jersey and Holstein cows. J. Dairy Sci. 91, 1103-1114. doi.org/10.3168/jds.2007-0724. [ Links ]
Anderson, T., Shaver, R., Bosma, P., & De Boer, V., 2007. Case Study: Performance of lactating Jersey and Jersey-Holstein crossbred versus Holstein cows in a Wisconsin confinement dairy herd. Prof. Anim. Sci. 23, 541-545. doi.org/10.1532/s1080-7446(15)31017-2. [ Links ]
Anonymous, 2017. Genetic improvement of dairy herds. Chapter 37 In: The Dairy Farming Handbook. Compiled by: Muller, C. J. C., Directorate: Animal Sciences, Research and Technology Development Services (RTDS), Western Cape Department of Agriculture. Elsenburg. pp 186-190. [ Links ]
Bangani, N.M., 2002. Oat silage in milk production systems in the Western Cape. MSc Thesis, Stellenbosch University. [ Links ]
Bangani, N.M., Muller, C.J.C., Dzama, K., Cruywagen, C.W.C. & Imbayarwo-Chikosi, V.E, 2022. Effects of production stage and fertility traits on milk production of pasture-grazed Holstein and Jersey cows in a Mediterranean-type climate region. S. Afr. J. Anim. Sci. 52, 552 - 563. doi.org/10.4314/sajas.v52i4.15. [ Links ]
Batajoo, K. K., & Shaver, R. D., 1994. Impact of non-fiber carbohydrate on intake, digestion, and milk production by dairy cows. J. Dairy Sci. 77, 1580-1588. doi.org/10.3168/jds.s0022-0302(94)77100-9. [ Links ]
Bauman, D. E., & Bruce Currie, W., 1980. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 63, 1514-1529. doi.org/10.3168/jds.s0022-0302(80)83111-0. [ Links ]
Bionaz, M., Hurley, W., & Loor, J., 2012. Milk protein synthesis in the lactating mammary gland: Insights from transcriptomics analyses. Chapter 11 in Milk Protein. Edited by Walter Hurley. Open access peer-reviewed edited volume, Intechopen Publishers. [ Links ]
Blake, R. W., & Custodio, A. A., 1984. Feed efficiency: A composite trait of dairy cattle. J. Dairy Sci.67, 2075-2083. doi.org/10.3168/jds.s0022-0302(84)81548-9. [ Links ]
Blake, R. W., Custodio, A. A., & Howard, W. H., 1986. Comparative feed efficiency of Holstein and Jersey cows. J. Dairy Sci.69, 1302-1308. doi.org/10.3168/jds.s0022-0302(86)80536-7. [ Links ]
Block, E., 2010. Transition cow research - what makes sense today? Proceedings of the High Plains Dairy Conference, Amarillo, Texas. pp 75-98. [ Links ]
Botha, P. R., Meeske, R. & Snyman, H. A., 2008. Kikuyu over-sown with ryegrass and clover: Dry matter production, botanical composition and nutritional value. Afr. J. Range Forage Sci., 25:3, 93-101, DOI: 10.2989/AJRF.2008.25.3.1.598. [ Links ]
Boyd, R.D. & Kensinger, R.S, 1998. Metabolic precursors for milk synthesis. In: The Lactating Sow. By: Verstegen, M.W.A. & Moughan, P.J. Editors Verstegen, M.W.A. Moughan, P.J. & Schrama J.W. Wageningen Academic Publishers. pp 69-93. [ Links ]
Cai, J., Wang, D. & Liu, J., 2018. Regulation of fluid flow through the mammary gland of dairy cows and its effect on milk production: A systematic review. J Sci Food Agric, 98: 1261-1270. doi 10.1002/jsfa.8605. [ Links ]
Cornell Net Carbohydrate and Protein System (version 6.1). Cornell University, Ithaca, NY. [ Links ]
Drackley, J. K., Dann, H. M., Douglas, G. N., Janovick Guretzky, N. A., Litherland, N. B., Underwood, J. P., & Loor, J. J., 2005. Physiological and pathological adaptations in dairy cows that may increase susceptibility to periparturient diseases and disorders. Ital. J. Anim. Sci. 4, 323-344. doi.org/10.4081/ijas.2005.323. [ Links ]
Fox, D. G., Tedeschi, L. O., Tylutki, T. P., Russell, J. B., Van Amburgh, M. E., Chase, L. E., Pell, A. N., & Overton, T. R., 2004. The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion. Anim. Feed Sci. Technol. 112, 29-78. doi.org/10.1016/j.anifeedsci.2003.10.006. [ Links ]
Friggens, N. C., Berg, P., Theilgaard, P., Korsgaard, I. R., Ingvartsen, K. L., L0vendahl, P., & Jensen, J., 2007. Breed and parity effects on energy balance profiles through lactation: Evidence of genetically driven body energy change. J. Dairy Sci.90, 5291-5305. doi.org/10.3168/jds.2007-0173. [ Links ]
Gaines, W. L. 1928. The energy basis of measuring milk yield in dairy cows. University of Illinois Agricultural Experiment Station. Bulletin 308. [ Links ]
Gallo, L., Carnier, P., Cassandro, M., Mantovani, R., Bailoni, L., Contiero, B., & Bittante, G., 1996. Change in body condition score of Holstein cows as affected by parity and mature equivalent milk yield. J. Dairy Sci. 79, 1009-1015. doi.org/10.3168/jds.s0022-0302(96)76452-4. [ Links ]
Gorewit, R.C., 1988. Lactation biology and methods of increasing efficiency. In designing foods: Animal product options in the marketplace. National Research Council (United States) Committee on Technological Options to Improve the Nutritional Attributes of Animal Products. National Academies Press, Washington DC. [ Links ]
Grainger, C. and Goddard, M. E., 2004. A review of the effects of dairy breed on feed conversion efficiency - an opportunity lost? Anim. Prod. Aust. 25, 77-80. [ Links ]
Hall, M. B., Larson, C. C., & Wilcox, C. J., 2010. Carbohydrate source and protein degradability alter lactation, ruminal, and blood measures. J. Dairy Sci. 93, 311-322. doi.org/10.3168/jds.2009-2552. [ Links ]
Ingvartsen, K.& Weisbjerg, M., 1993. Jersey cows have a higher feed intake capacity and higher rate of passage than Friesian cows. Archiv. Fuer. Tierzucht. 5:495-498. [ Links ]
Knowlton, K. F., Wilkerson, V. A., Casper, D. P., & Mertens, D. R., 2010. Manure nutrient excretion by Jersey and Holstein cows. J. Dairy Sci. 93, 407-412. doi.org/10.3168/jds.2009-2617. [ Links ]
Kristensen, T., Jensen, C., 0stergaard, S., Weisbjerg, M. R., Aaes, O., & Nielsen, N. I., 2015. Feeding, production, and efficiency of Holstein-Friesian, Jersey, and mixed-breed lactating dairy cows in commercial Danish herds. J. Dairy Sci. 98, 263-274. doi.org/10.3168/jds.2014-8532. [ Links ]
Lee, J. Y., & Kim, I. H., 2006. Advancing parity is associated with high milk production at the cost of body condition and increased periparturient disorders in dairy herds. J. Vet. Sci. 7, 161-166. doi.org/10.4142/jvs.2006.7.2.161. [ Links ]
Lin, Y., Sun, X., Hou, X., Qu, B., Gao, X., & Li, Q., 2016. Effects of glucose on lactose synthesis in mammary epithelial cells from dairy cow. BMC Vet. Res. 12, 81. doi.org/10.1186/s12917-016-0704-x. [ Links ]
Liu, H., Zhao, K., & Liu, J., 2013. Effects of glucose availability on expression of the key genes involved in synthesis of milk fat, lactose, and glucose metabolism in bovine mammary epithelial cells. PLoS One doi.org/10.1371/journal.pone.0066092. [ Links ]
Mackle, T. R., Parr, C. R., Stakelum, G. K., Bryant, A. M., & MacMillan, K. L. 1996., Feed conversion efficiency, daily pasture intake, and milk production of primiparous Friesian and Jersey cows calved at two different liveweights. New Zeal. J. Agric. Res. 39, 357-370. doi.org/10.1080/00288233.1996.9513195. [ Links ]
Macrae, A.I., Burrough, E., Forrest, J., Corbishley, A., Russell, G. & Shaw, D.J., 2019. Risk factors associated with excessive negative energy balance in commercial United Kingdom dairy herds. The Veterinary Journal, 250, 1523. doi.org/10.1016/j.tvjl.2019.06.001 [ Links ]
Mepham, T. B., 1982. Amino acid utilization by lactating mammary gland. J. Dairy Sci. 65, 287-298. doi.org/10.3168/jds.s0022-0302(82)82191-7. [ Links ]
Muller, C. J. C., & Botha, J. A., 1998. The comparative performance of primiparous Holstein Friesland and Jersey cows on complete diets during summer in a temperate climate. S. Afr. J. Anim. Sci. 28, 161 - 166. [ Links ]
National Research Council, 2001. Nutrient Requirements of Dairy Cattle. Seventh Revised Edition. The National Academies Press, Washington DC. doi.org/10.17226/9825. [ Links ]
Nutritional Dynamic System (NDS) Professional Software (version 6.5, 2008 to 2018). RUM&N., Reggio Emilia, Italy. [ Links ]
Olijhoek, D. W., Lovendahl, P., Lassen, J., Hellwing, A. L. F., Höglund, J. K., Weisbjerg, M. R., Noel, S. J., McLean, F., H0jberg, O., & Lund, P., 2018. Methane production, rumen fermentation, and diet digestibility of Holstein and Jersey dairy cows being divergent in residual feed intake and fed at 2 forage-to-concentrate ratios. J. Dairy Sci. 101, 99269940. doi.org/10.3168/jds.2017-14278. [ Links ]
Prendiville, R., Pierce, K. M., & Buckley, F., 2009. An evaluation of production efficiencies among lactating Holstein-Friesian, Jersey, and Jersey * Holstein-Friesian cows at pasture. J. Dairy Sci. 92, 6176-6185. doi.org/10.3168/jds.2009-2292. [ Links ]
Rastani, R. R., Andrew, S. M., Zinn, S. A., & Sniffen, C. J., 2001. Body composition and estimated tissue energy balance in Jersey and Holstein cows during early lactation. J. Dairy Sci. 84, 1201 -1209. doi.org/10.3168/jds.s0022-0302(01)74581-x. [ Links ]
Retief, N., 2000. Breed differences in ruminal digestibility of forages in dairy cows receiving high concentrate diets. MSc. Thesis. University of Stellenbosch. pp32. [ Links ]
Rezaei, R., Wu, Z., Hou, Y., Bazer, F. W., & Wu, G., 2016. Amino acids and mammary gland development: Nutritional implications for milk production and neonatal growth. J. Anim. Sci. Biotechnol. 7, 20. doi.org/10.1186/s40104-016-0078-8. [ Links ]
Rodriguez, A., Mellado, R. & Bustamante, H., 2020. Prepartum fat mobilization in dairy cows with equal body condition and its impact on health, behavior, milk production, and fertility during lactation. Animals (Basel). 22, 10(9):1478. doi: 10.3390/ani10091478. [ Links ]
Ross, S. A., Chagunda, M. G. G., Topp, C. F. E., & Ennos, R., 2015. Biological efficiency profiles over the lactation period in multiparous high-producing dairy cows under divergent production systems. Arch. Anim. Breed. 58, 127-135. doi.org/10.5194/aab-58-127. [ Links ]
Statistical Analysis Systems, 2012. SAS Enterprise guide Software, Version 7.1. SAS® Institute Inc., Cary, North Carolina, USA. [ Links ]
Tedeschi, L. O., Cavalcanti, L. F. L., Fonseca, M. A., Herrero, M., & Thornton, P. K., 2014. An overview of dairy cattle models for predicting milk production: Their evolution, evaluation, and application for the Agricultural Model Intercomparison and Improvement Project (AgMIP) for livestock. CCAFS Working Paper no. 94. Copenhagen, Denmark: CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS). [ Links ]
Tessari, R., Berlanda, M., Morgante, M., Badon, T., Gianesella, M., Mazzotta, E., Contiero, B. & Fiore, E., 2020. Changes of plasma fatty acids in four lipid classes to understand energy metabolism at different levels of non-esterified fatty acid (NEFA) in dairy cows. Animals (Basel). 13, 10(8):1410. doi: 10.3390/ani10081410. [ Links ]
Thomson, N.A., Kay, J.K. & Bryant, M.O. 2001. Effect of stage of lactation on the efficiency of Jersey and Friesian cows at converting pasture to milk production or liveweight gain. Proceedings of the New Zealand Society of Animal Production. 61, 213-216. [ Links ]
Tylutki, T. P., Fox, D. G., Durbal, V. M., Tedeschi, L. O., Russell, J. B., Van Amburgh, M. E., Overton, T. R., Chase, L. E., & Pell, A. N., 2008. Cornell Net Carbohydrate and Protein System: A model for precision feeding of dairy cattle. Anim. Feed Sci. Technol. 143, 174-202. doi.org/10.1016/j.anifeedsci.2007.05.010. [ Links ]
Tyrrell, H.F. & Reid, J.T., 1965. Prediction of the energy value of cow's milk. J. Dairy Sci. 48,1215-1223. [ Links ]
VandeHaar, M. J., 2011. Increasing efficiency of nutrient use to enhance profit and environmental stewardship. Proceedings: 22nd Annu. Florida Rumin. Nutr. Symp. Gainesville, FL. [ Links ]
VandeHaar, M. J., Armentano, L. E., Weigel, K., Spurlock, D. M., Tempelman, R. J., & Veerkamp, R., 2016. Harnessing the genetics of the modern dairy cow to continue improvements in feed efficiency. J. Dairy Sci. 99, 4941 -4954. doi.org/10.3168/jds.2015-10352. [ Links ]
Varga, G. A., Kolver, E. S., & Suppl, M., 1997. Fiber utilization by ruminants microbial and animal limitations to fiber digestion and utilization. J. Nutr. 127, 819S-823S. [ Links ]
Walter, L.L., Gartner, T., Gernand, E., Wehrend, A. & Donat, K., 2022. Effects of parity and stage of lactation on trend and variability of metabolic markers in dairy cows. Animals (Basel). 12(8), 1008. doi: 10.3390/ani12081008. [ Links ]
Washburn, S. P., White, S. L., Green, J. T., & Benson, G. A., 2002. Reproduction, mastitis, and body condition of seasonally calved Holstein and Jersey cows in confinement or pasture systems. J. Dairy Sci. 85, 105-111. doi.org/10.3168/jds.s0022-0302(02)74058-7. [ Links ]
Wathes, D.C., Cheng, Z., Bourne, N., Taylor, V.J., Coffey, M.P. & Brotherstone, S., 2007. Differences between primiparous and multiparous dairy cows in the inter-relationships between metabolic traits, milk yield and body condition score in the periparturient period. Domest. Anim. Endocrinol. 33, 203-225. [ Links ]
Xue, B., Yan, T., Ferris, C. F., & Mayne, C. S., 2011. Milk production and energy efficiency of Holstein and Jersey-Holstein crossbred dairy cows offered diets containing grass silage. J. Dairy Sci. 94, 1455-1464. doi.org/10.3168/jds.2010-3663. [ Links ]
Submitted 18 January 2023
Accepted 26 February 2023
Published 9 July 2023
# Corresponding author: vuyo.bangani@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}