










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.53 n.2 Pretoria 2023
Nutritional regulation of gut health through the gastrointestinal tract microbiota in weaning piglets
L. Musungwe; L. Dong
Department of Animal Nutrition and Feed Science Technology, College of Animal Science and Technology, Yangzhou University, Yangzhou, Jiangsu Province 215009, People's Republic of China
ABSTRACT
Piglets experience stresses that impact intestinal homeostasis and result in a high incidence of diarrhoea and reduced growth performance during weaning. Recent studies have indicated that gastrointestinal tract microbiota can be nutritionally regulated in ways that can benefit and promote gut health in weaning piglets. In this review, we discuss different components of nutrition and their effects on microbial populations, diversity, and activity and, in turn, how these altered microbial structures and activities affect gut health in weaning piglets. In addition, we discuss some of the inner mechanisms through which microbiota regulate and exert their effects on gut health, focusing particularly on adaptive and innate immunity, inflammatory responses and intestinal barrier integrity. Most of the mechanisms involve the TLR and NF-kB signalling pathways, and miRNA regulation. An understanding of the effects of nutritional regulation of microbiota on gut health and its underlying mechanisms can be applied in developing nutritional regimens that promote beneficial microbial structures for gut health.
Keywords: dietary fibre, inner mechanisms, intestinal barrier integrity, organic acids, prebiotics, probiotics
1. The effects of weaning on the most common microbiota in the gastrointestinal tract of piglets.
The composition of natural microflora in the gastrointestinal tract is not absolute because it develops gradually over time, with many changes occurring during weaning and infections (Zhao et al., 2020), and it is influenced by the interaction between the host and the microbiota. Foetuses are devoid of microbial species and can be regarded as germfree. Neonates are exposed to diverse microbes in the vaginal canal during parturition and in the suckling environment during nursing, resulting in microbial seeding. The nursing piglets are also exposed to microbiota on the sow skin, faeces, and mucosal surfaces, resulting in the piglets developing an adult-like microbial structure due to seeding and microbial succession (Guevara et al., 2019). However, as illustrated by a previous study, dietary, environmental and social stresses induced by the weaning transition result in alterations in the gut microbiota with loss of microbial diversity (Gresse et al., 2019). In this section, these changes in gastrointestinal tract microbiota related to weaning in piglets are identified.
The microbiota in the gut of a piglet are mainly composed of Lactobacillus, Actinobacillus, Romboutsia, Escherichia Shigella, Bacteroides, Terrisporobacter, and Campylobacter genera at all gastrointestinal tract sites (Li et al., 2018; Adhikari & Kim et al., 2019; Zhao et al., 2020). However, during the weaning transition, a small number of dominant species constitute the gut microbiota. An increase in fermentative anaerobes and a decrease in the abundance of Lactobacillus group bacteria are observed (Gresse et al., 2019). The core bacterial phyla found all along the piglets' digestive tract during weaning are Firmicutes, Cyanobacteria, and Actinobacteria, with Firmicutes being the most abundant with a relative abundance of 97% (Li et al., 2017). These findings on the decrease in microbial diversity coincide with the findings of Kraler et al. (2016). In the research by Kraler et al. (2016) the phyla Firmicutes, Proteobacteria, and Bacteroidetes were abundant (Hu et al., 2016). Proteobacteria and Firmicutes constitute approximately 25% and 43.5% of the total mean microbiota relative abundance, respectively (Kraler et al., 2016; Wang et al., 2019). The reduction in microbial diversity in the gastrointestinal tract of piglets during weaning gives rise to opportunistic pathogens such as E. coli, which is a predominant group in both the small intestine and the large intestine in piglets during weaning but is in relatively low abundance under normal health conditions (Chen et al., 2013). The increase in the relative abundance of E. coli during weaning further disrupts the piglets' gut microbiome, leading to diarrhoea and growth reduction experienced through the post-weaning period.
The abundance of beneficial bacterial species such as Lactobacillus and Bifidobacteria over pathogenic ones in the entire digestive tract is preferable for the stability of the intestinal immune system and the body. The health of organisms depends primarily on the composition of intestinal microflora, which is a key area for further development in piglets during weaning to enhance health benefits to the organism. The microbiota composition across the gastrointestinal tract in the weaning piglets is shown in Table 1. In conclusion, weaning alters the gut microbiome in piglets by decreasing microbial diversity, downregulating the abundance of beneficial bacteria, and increasing the proliferation of pathogenic bacteria.
2. Influences of microbiota on weaning piglets' gastrointestinal function and health
Intestinal microbiota composition and metabolic activities are important parameters affecting gastrointestinal health in animals (Blachier et al., 2013). The composition and diversity of the microbiota affects gut epithelial integrity, barrier function, and immunity (Su et al., 2018). In this section, the influences of microbiota on these gastrointestinal functions and health in weaning piglets are identified.
During weaning, the abundance of Lactobacillus group bacteria in the gastrointestinal tract of piglets is very low. Normally, Lactobacillus predominates the small intestine and has a positive effect on piglets' gut health, being vital itself in disease prevention. However, the relatively low abundance of Lactobacillus in the gut of weaning piglets decreases mucin production (secreted by intestinal goblet cells), thus disrupting the gut barrier through a permeable, thin mucus layer (Fouhse et al., 2016). The relatively low abundance of Lactobacillus downregulates the gene expression of occludin and cingulin (Anderson et al., 2010), which are paramount tight junction proteins in Caco-2 cells. Furthermore, the abundance of lactic acid-producing bacteria (Lactobacillus and Bifidobacteria) decreases during weaning, which increases intestinal pH and in turn, increases disease susceptibility (Konstantinov et al., 2004) since acidic gut pH is bactericidal. The decreased abundance of Bifidobacterium inhibits the expression of an ABC transporter responsible for the stimulation of fructose metabolism to acetate. Acetate promotes the integrity and barrier function of epithelial cells, preventing the passage of Shiga toxins past the barrier, a function that is absent in weaning piglets. The reduced number of Lactobacilli and Bifidobacteria in weaning piglets also reduces protein bacteriocins, which increases the growth of related bacterial species (Kawai et al., 2004; Kantas et al., 2014). Lactobacillus on its own adheres to ileal epithelial cells and prevents the adhesion of other pathogenic microbes to the epithelial cells in a mechanism of competitive exclusion. Prevotella is abundant in the gut of piglets during weaning.
The abundance of Prevotella in the gut and the accompanying microbial fermentation stimulates the secretion of mucosal immunoglobulins (IgA), restricting pathogen access through intestinal epithelial cells (Fouhse et al., 2016), which also improves gut immunity in weaning piglets. On the other hand, E. coli, an infectious pathogen with negative effects on piglet gut health, is in relatively high abundance in the gut of weaning piglets. Escherichia coli cells inhabit most sectors of the gastrointestinal tract, resulting in tight junction disruption through the destabilization and dissociation of tight junction proteins (occludin and cingulin) from epithelial cells, which affects intestinal barrier integrity and increases the translocation of pathogens and toxins penetrating the barrier (Chen et al., 2013).
The gut microbiota that are altered during weaning also caused changes in metabolites such as short-chain fatty acids (SCFAs) through microbial fermentation in the colon and caecum of the gastrointestinal tract. Butyrate is one of the SCFAs that inhibit the growth of both gram-positive and gram-negative bacteria in the colon. Butyrate is utilized as an energy source for colonic epithelial cell proliferation and differentiation, which improves colonic barrier function (Kinoshita et al., 2002). Butyrate at high concentrations in the gut also enhances intestinal barrier function and increases the mRNA expression of tight junction proteins (occludin, zona occludin 1, and claudin 1) in the duodenal, jejunal, and ileal mucosa of weaning piglets (Huang et al., 2015; Zhao et al., 2019).
SCFAs also protect the mucosal immune system from inflammation, especially in cells harbouring the SCFA receptor, GPR43, which has numerous positive effects on gut health, including increasing epithelial integrity, decreasing bacterial translocation, decreasing inflammation, increasing epithelial barrier repair, increasing mucus secretion, decreasing neutrophil chemotaxis, and decreasing reactive oxygen species. Feng et al. (2018), concluded that SCFAs were able to prevent disruption of the intestinal barrier by lipopolysaccharides through the inhibition of autophagy and the NLRP3 inflammasome. SCFAs can alleviate the effects of the NLRP3 inflammasome and autophagy in the intestinal mucosa. These microbial metabolites prevent intestinal mucosal atrophy and decrease apoptosis. The intestinal microbiota plays a pivotal role in the regulation of intestinal barrier function, intestinal immunity, and overall piglet gut health. Impaired microbiota barrier function of the gastrointestinal tract leads to compromised piglet health.
3. Effects of nutritional supplements on gut health through the gastrointestinal tract microbiota in weaning piglets
3.1 Prebiotics - fermentable carbohydrates
Short-chain fructooligosaccharide (SCFO) supplementation has been shown to have modulatory effects on the microbiota composition in the hindgut of weaning piglets (Huang et al., 2015). Administration of SCFOs in previous studies improved the health of weaning piglets by selectively stimulating and promoting the growth of beneficial microbiota in the gastrointestinal tract of weaning piglets (Tsukahara et al., 2003; Xue et al., 2018; Zhao et al., 2019). SCFO supplementation had positive effects on microbiota composition in weaning piglets by increasing the relative abundance of beneficial bacteria (Lactobacillus, Prevotella, and Bifidobacterium species) in the colon (Zhao et al., 2019). This coincides with the effects of fructooligosaccharides (FOSs) (Tsukahara et al., 2003). Konstantinov et al., 2004, supplemented the diet of weaning piglets with a combination of fermentable carbohydrates and reported a prevalence of Lactobacillus reuteri and Lactobacillus amylovorous-like populations in the ileum and colon of the piglets. The increased abundance of Prevotella in the colon of weaning piglets resulted in an increased luminal SCFA level (acetate, butyrate, and propionate) (Xue et al., 2018). The acetate produced through microbial fermentation has anti-inflammatory effects that maintain intestinal homeostasis and promote intestinal barrier function (Tsukahara et al., 2003; Huang et al., 2015; Zhao et al., 2019). The collective effects of microbiome enhancement by different prebiotic supplements confers beneficial health effects such as increased intestinal barrier function and intestinal barrier integrity; suppression of mucosal inflammation, indicated by low diarrhoea indices; low serum D-lactate, IL-1 beta, IL-6, and tumour necrosis factor alpha concentrations; and increased tight junction protein mRNA expression in the ileal and jejunal mucosa. Based on these studies, prebiotics modulate the gut microbiota by increasing the abundance of beneficial microbial species such as Lactobacillus, Prevotella, and Bifidobacteria, which produce metabolites that promote intestinal barrier integrity and function and create an environment that substantially enhances weaning piglet gut health.
3.2 Probiotics
Probiotics are live microorganisms administered in sufficient amounts to confer health benefits (Hill et al., 2014). As natural bioregulators, probiotics help maintain the balance of the gastrointestinal tract ecosystem and prevent colonization of the gastrointestinal tract with pathogenic bacteria. Supplementation with probiotic complexes in weaning piglets increases the abundance of lactic acid bacteria, including Lactobacillaceae, Bifidobacteriaceae, total anaerobes and total aerobes, while decreasing Clostridiaceae, Peptostrepcoccaceae and Enterobacteriaceae faecal counts (Collington et al., 1990; Kantas et al., 2014; Luise et al., 2019). In a study by Luise et al. (2019, supplementation with probiotics containing Bacillus subtilis and B. amyloliquefaciens reduced the abundance of Enterobacteriaceae, and this effect on microbial diversity decreased the risk of pathogen development and promoted gut health in weaning piglets. The Enterobacteriaceae family is regarded as a biomarker of gut health since an increase in its prevalence is associated with intestinal dysbiosis in weaning piglets (Trevisi et al., 2018). The same results were observed after piglets were orally administered a mixture of probiotic strains Lactobacillus plantarum, L. acidophilus, L. casei, and Streptococcus faecium (Collington et al., 1990).The overall health of weaning piglets was improved through the reduction of enteric pathogenic microbes and the increase in lactic acid bacteria in the gut after supplementation with a probiotic (Kantas et al., 2014). Campylobacter jejuin and Enterobactericeae were the least abundant, Bifidobacterium was the most abundant, and Salmonella, L. monocytogenes, and E. coli were absent (Kantas et al., 2014). Probiotic supplementation increased the relative abundance of Ruminococcaceae in the gut of piglets, resulting in increased gut SCFA concentrations, which affected gut health, inhibited Salmonella growth, and exhibited anti-inflammatory effects.
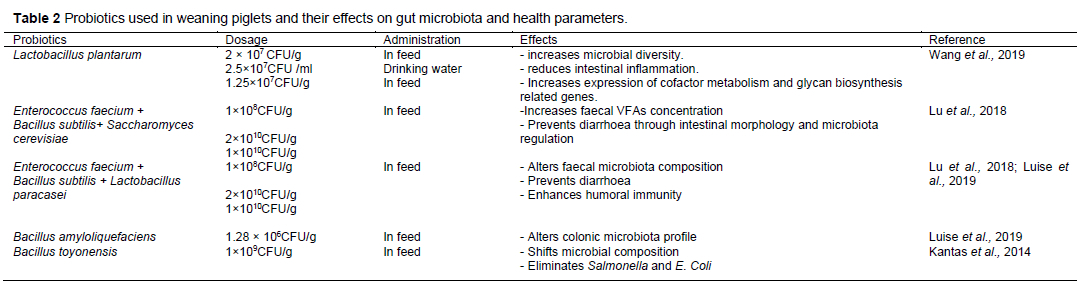
Other studies have explored the supplementation of probiotics in combination with prebiotics such as fructooligosaccharides, maltodextrins, and polyunsaturated fatty acids (Nemcova et al., 2007), which requires more research and can be implemented to further improve weaning piglet gut health. In conclusion, probiotics as feed components alter the gastrointestinal tract microbial composition, increase the relative abundance of beneficial microbes, and decrease the abundance of pathogenic microbes in weaning piglets. The microorganisms used as probiotics in weaning piglets and their effects on gut microbiota and health parameters are shown in Table 2.
3.3 Organic acids and fatty acids (acidifiers)
Dietary acidifiers are a major alternative to antibiotics for improving weaning piglet performance and inhibiting the proliferation of pathogenic microbes in the gastrointestinal tract (Kim et al., 2005; Kil et al., 2011). The addition of organic acids to diets lowers the pH of the stomach and gastrointestinal tract, prohibiting invasion and proliferation of pathogenic bacteria (Kil et al., 2011). In a study by Han et al. (2018), piglets fed a diet supplemented with a combination of organic acids and medium fatty acids replacing chlortetracycline decreased the relative abundance of Proteobacteria, increased that of Firmicutes, and markedly increased that of Lactobacillus and Faecalibacterium. The relative increase in Firmicutes, Lactobacillus, and Faecalibacterium can be attributed to the reduction in E. coli, Salmonella, Bacteroidaceae, and Enterococcus in the duodenum and jejunum of weaning piglets (Roth & Kirchgessner, 1998; Han et al., 2018). Roth & Kirchgessner (1998) found the same results when they supplemented piglet diets with formic acid, which exhibited antibacterial effects, reducing the abundance of Bacillus, E. coli, and Salmonella while increasing Bifidobacterium counts in the duodenum and jejunum. An experimental diet fed to piglets reduced the proportion of coliform bacteria and increased that of Lactobacillus. Furthermore, Cole et al. (1968) found that there was a reduction in haemolytic E. coli counts in the duodenum and jejunum of weaning piglets fed 0.8% lactic acid in drinking water. However, in another study, no effect of Lactobacillus and total aerobic bacterial count was found after organic acid supplementation (Lang et al., 2018). Supplementation with organic acids yields a microbial population dominated by Firmicutes, Bacteroidetes, and Proteobacteria. These acidifiers are both bactericidal and bacteriostatic, and undissociated organic acids are lipophilic and can penetrate the bacterial cell, reduce internal pH, and disrupt normal physiology. Formic acid, for example, dissociates into cations and anions after passing the bacterial cell wall, disrupting bacterial protein synthesis. The effects of dietary acidifiers on microbial populations, however, are not clear in most studies and require further research involving modern molecular techniques for microbial count accuracy. Most researchers found no marked effect, either positive or negative, of acidifiers on Lactobacillus counts in the gastrointestinal tract of weaning piglets (Roth & Kirchgessner 1998; Kim et al., 2005; Kil et al., 2011; Lang et al., 2018). However, from past research analysis, it is noted that there are limited data on the effects of acidifiers on microbial populations in weaning piglets; hence, there is a need for more research to verify these effects.
3.4 Dietary fibre
Dietary fibre in weaning piglet diets may be fermented by gut microbiota to produce health-promoting SCFAs that modify intestinal barrier function and microbiota composition (Chen et al., 2013). Feeding dietary fibre in the form of wheat bran fibre or pea fibre in diets increased the relative abundance of ileal Lactobacillus and colonic Bifidobacterium, while decreasing that of E. coli in the ileum and colon of piglets (Chen et al., 2013). Dietary fibre supplementation increased acetate, propionate, butyrate, and total VFA concentrations in the colon. In recent studies, the inclusion of wheat bran as a dietary fibre combined with ZnO in diets increased E. coli counts (Molist et al., 2011), contrary to the results observed under the inclusion of wheat bran only, which decreased E. coli counts (Molist et al., 2010) and improved gut health by modulating the activity and composition of gut microbiota. The negative interaction of wheat bran and ZnO raises the consideration of including phytase enzymes to reduce the levels of therapeutic ZnO (Molist et al., 2011). Wheat bran in early weaning piglets promoted intestinal fermentation, reduced the incidence of diarrhoea, and reduced the attachment of E. coli K88 to the ileal mucosa. It was observed that the addition of ZnO decreased the contents of SCFAs, increased coliform bacteria, and decreased some of the counts of microbial groups promoted by wheat bran supplementation (Molist et al., 2010). Dietary fibre therefore has effects on gastrointestinal tract digestive processes by increasing feed bulkiness and gut retention time and promoting beneficial microbial species, enhancing weaning piglet gut health.
4. Inner mechanisms
Different nutrients in the diets of swine alter the gastrointestinal tract microbial structure. The resultant gut microbial populations affect the overall gut health of pigs through various pathways and mechanisms. The underlying mechanisms mainly involve the regulation of enzymes, genes, micro-RNA (miRNA), and certain metabolic pathways.
Signalling pathways: The interaction and crosstalk between microbes and the host system is transmitted through signalling pathways involving different molecules and body organs (Nicholson et al., 2012). In these interactions, the primary and secondary metabolites involved include SCFAs, antimicrobial compounds, medium- and long-chain fatty acids, neurotransmitters, and bile acid metabolites, which are all associated with different microbial species activities in the gut. Microbes (Lactobacillus, Bifidobacteria, Enterobacter, Bacteroides) in abundance in the gastrointestinal tract of weaning piglets participate in bile acid metabolism through their hydrolase enzymes. The enzymes are involved in bile salt deconjugation and dehydroxylation, producing transformed metabolites (cholate, hyocholate, deoxycholate, taurocholate, lithocholate, taurohyocholate) that are easily absorbed in the gut and act as interaction molecules with the host in crosstalk, activating biological functions that maintain intestinal barrier function and integrity in weaning piglets (Ridlon et al., 2006). The host gut senses these microbial metabolites via G-protein- or microbiota-associated molecular pattern (MAMP) receptor recognition on microbial cell surfaces (Frosali et al., 2015). Microbial cell surface antigens activate pathogen-recognition receptors (PRRs) expressed on the mucosa and immune cells (Metzler-Zebeli et al., 2018). As a result, transcriptional and intracellular signalling cascades are induced, leading to inflammatory responses; recruitment of phagocytic cells; mobilization of antigen-presenting cells; and activation of genes that code for proinflammatory cytokines, antimicrobial peptides and anti-apoptotic factors in promotion and maintenance of gastrointestinal health (Watt et al., 2020). The cascades are regulated by histone modification, miRNAs, and DNA methylation and chromatin remodelling at the gene level.
Enzyme and gene regulation: Butyrate and propionate are produced in large amounts due to the abundance of anaerobic bacteria in the gut of weaning piglets, and they exert direct anti-inflammatory and anti-apoptotic effects on the gut mucosa. These SCFAs lower inflammation by inhibiting the expression of the enzyme, histone deacetylase, or by downregulating proinflammatory mediator activities (IL-6, IL-12, and nitric oxide). The absence of histone deacetylase affects histone deacetylation, which regulates nuclear gene expression towards host immunity. Histone deacetylase is responsible for cleaving the acetyl groups of lysine amino acid residues in histone subunits, allowing DNA to wrap tightly around histones (Watt et al., 2020). As a result, DNA transcription is repressed and inflammatory response pathways are inhibited in the gut of weaning piglets.
Histone deacetylase inhibition occurs simultaneously with gene acetylation. Intestinal bacteria-derived SCFAs promote transcriptionally-permissive acetyl modifications at gene promoters and enhancers through their histone deacetylase inhibitory activities (Watt et al., 2020). While histone deacetylase inhibition represses DNA transcription, gene acetylation triggers the expression of specific genes within the DNA. Acetylation at immune gene promoters improves host innate immune memory and improves the ability of piglets to resist infection by pathogenic microbes. Furthermore, in the presence of gut Firmicutes and Bacteroidetes, the SCFA, butyrate, is absorbed by the colon mucosa, leading to increased mRNA levels of E-cadherin and ZO-1 in colonic tissues (Kawai et al., 2004). Increases in Firmicutes and Bacteroidetes are associated with increases in the mRNA levels of the anti-inflammatory cytokines, IL-4, IL-10, and TGF-B1. Low-grade inflammation in the gut is triggered by bacteria via the TLR4-NFkB signalling pathway (McKenzie et al., 2017).
miRNA regulation: The gut microbiota can modulate host intestinal miRNAs and regulate intestinal epithelial tight junction protein expression. These ultimately function in regulating innate and adaptive intestinal immunity (Bi et al., 2020). The intestinal microbiota negatively regulate the expression of mR-10a in dendritic cells by interacting with the TLR signalling pathway to promote intestinal immunity. In humans, the development of chronic Inflammatory bowel disease is suppressed by targeting IL12/IL-23p40 expression. Listeria specifically inhibits miR-192, miR-215, and miR-200b expression, avoiding immunity in the intestine for infection to progress, whereas Lactobacillus casei BL23 and Lactobacillus paracasei reverse these effects of Listeria on miRNA levels under infection in the gut (Bi et al., 2020).
Metabolic pathway regulation: Microbial metabolites (e.g., n-butyrate) and structural components (LPS, CpG, DNA motifs) induce intestinal alkaline phosphatase (IAP) and inducible heat shock protein (iHSP) expression in the gut. IAP dephosphorylates microbial components and downregulates microbe-induced inflammation through inhibition of the NF-kB proinflammatory pathway in enterocytes (Lalles, 2016). IAP also detoxifies proinflammatory free luminal ATP, which inhibits inflammation and propels the differentiation of Th17 lymphocytes responsible for the production of IL-17 and IL-22 cytokines that promote neutrophil tissue infiltration and stimulate antibacterial peptide production, respectively (Lalles, 2016). Pigs have three IAP gene copies that are strongly inhibited in piglets post early weaning, contributing to post-weaning disorders and enhanced piglet sensitivity to enteric infection. IAPs and iHSPs indirectly control intestinal barrier function through the stimulation of tight junction protein gene expression (zona occludens and occludin) and their correct localization in cells. Bacteroides, Faecalibacterium, Lactobacillus, and Bifidobacteria strains were reported to induce iHSP production in epithelial cells (Rakoff-Nahoum et al., 2004). LPS also directly activates TLR4/TLR2/TLR9 gene expression to increase gut immunity gene expression (Lalles, 2016; Sun et al., 2017; Gardiner et al., 2020).
Increased abundance of anaerobic bacteria in the gastrointestinal tract of weaning piglets leads to high levels of SCFAs produced by microbial fermentation. SCFAs activate the expression of GPRs (GPR43, GPR41, GPR109A), which trigger mucosal pathways that moderate inflammatory responses and influence epithelial integrity and immunoglobulin-A and macrophage antibody responses (Frosali et al., 2015; McKenzie et al., 2017). Regulatory T cell and dendritic cell activities are also influenced by GPRs (Sun et al., 2017).
5. Conclusions and Outlook
Weaning alters the composition of the most common microbiota in the gastrointestinal tract of piglets, leading to a minimal number of dominant species in the gastrointestinal tract microbiota. Microbiota diversity and composition influences gastrointestinal function and health by affecting gut epithelial integrity, barrier function, immunity, and metabolic activities in weaning piglets. The weaning transition induces stress in weaning piglets, which is reflected by shifts in the gastrointestinal tract microbiota composition, resulting in ill health or no adverse effects, depending on the nutritional interventions employed. Dietary acidifiers and probiotics affect the immune system of piglets, reducing the risk of pathogen development and modifying gene expression to maintain gut health in weaning piglets by altering the gut microbial profile, inhibiting the proliferation of pathogenic microbes, and increasing that of beneficial microbes. Prebiotics improve gut health in weaning piglets by maintaining intestinal barrier integrity, suppressing mucosal inflammation, and increasing tight junction protein mRNA expression through modulatory effects on microbiota composition in the hindgut of weaning piglets. Dietary fibre promotes gut health by modifying intestinal barrier function through its influence on microbial composition. All these dietary effects on microbiota and gut health occur through a series of inner mechanisms, including the regulation of enzymes, genes, miRNA, and metabolic pathways in weaning piglets. However, studies have been centred mostly on butyrate, which is not quite representative of all SCFAs; therefore, more research on SCFAs in general is necessary, and at present, it is not clear how different sources of dietary fibre alter gut microbiota in weaning piglets. More research on these effects is necessary to pave the way for dietary fibre as a nutritional supplement in piglets during the weaning transition. Future studies can also address the interaction between different nutritional regimens and environmental factors in regulating gut health in piglets during the weaning transition.
Acknowledgements
Not applicable
Authors' contributions
LM conceptualized the study, wrote the original draft, and edited the final manuscript. LD supervised the study, reviewed, and edited the final manuscript. Both authors have read and approved the final manuscript.
Conflict of Interest
The authors declare no potential conflict of interest relevant to this article.
References
Adhikari, B., Kim, S.W, Kwon, Y.M., 2019. Characterization of microbiota associated with digesta and mucosa in different regions of gastrointestinal tract of nursery pigs. Int. J. Mol. Sci. 20(7), 1630. doi.org/10.3390/ijms20071630 [ Links ]
Anderson, R.C., Cookson, A.L., McNabb, W.C., Park, Z., McCann, M.J., Kelly, W.J. & Roy N.C., 2010. Lactobacillus plantarum MB452 enhances the function of the intestinal barrier by increasing the expression levels of genes involved in tight junction formation. BMC Microbiology. 1471-2180. https://www.biomedcentral.com/1471-2180/10/316 [ Links ]
Bi, K., Zhang, X., Chen, W. & Diao, H., 2020. MicroRNAs regulate intestinal immunity and gut microbiota for gastrointestinal health: A comprehensive review. Genes. 11, 1075. doi:10.3390/genes11091075 [ Links ]
Blachier, F., Wu, G., Yin, Y., 2013. Nutritional and Physiological Functions of Amino Acids in Pigs, 1st edition. Springer-Verlag-Wien. [ Links ]
Chen, H., Mao, X., Yu, J.H.B., Huang, Z., Yu, J., Zheng, P. & Chen, D., 2013. Dietary fibre affects intestinal mucosal barrier function and regulates intestinal bacteria in weaning piglets. Br. J. Nutr. 110, 1837-1848. doi:10.1017/S0007114513001293 [ Links ]
Cole, D.J., Beal, R.M., Luscombe, J.R., 1968. The effect on performance and bacterial flora of lactic acid, propionic acid, calcium propionate, and calcium acrylate in the drinking water of weaned pigs. Vet Rec. 2, 83(18), 459-64. doi:10.1136/vr.83.18.459 [ Links ]
Collington, G.K., Parker, D.S. & Armstrong, D.G., 1990. The influence of either an antibiotic or a probiotic in the diet on the development of digestive enzymes activity in the pig. Br. J. Nutr. 64, 39-70. doi:10.1079/bjn19900009 [ Links ]
Feng, Y., Wang, Y., Huang, Y. & Wang, F., 2018. Short-chain fatty acids manifest stimulative and protective effects on intestinal barrier function through the inhibition of NLRP3 inflammasome and autophagy. Cell Physio Biochem. 49, 190-205. DOI:10.1159/000492853 [ Links ]
Fouhse, J.M., Zijlstro, R.T., Willing, B.P., 2016. The role of gut microbiota in the health and disease of pigs. Nutritional Science University of Alberta.10, 2527. doi:10.2527/af.2016-0031 [ Links ]
Frosali, S., Pagliari, D., Gambassi, G., Landolfi, R., Pandolfi, F., Cianci, R., 2015. How the intricate interaction among toll-like receptors, microbiota, and immunity can influence gastrointestinal pathology. J. Immunol Res. 489821. doi:101155/2015/489821 [ Links ]
Gardiner, G. E., Metzler-Zebeli, B.U. & Lawlor, P.G., 2020. Impact of intestinal microbiota on growth and feed efficiency in pigs: A review. Microorganisms, 8, 1886. doi:10.3390/microorganisms8121886 [ Links ]
Gresse, R., Durand, F.C., Duru, L., Blanquet-Diot, S. & Forano, E., 2019. Microbiota composition and functional profiling throughout the gastrointestinal tract of commercial weaning piglets. Microorganisms. 77, 343 doi:10.3390/microorganisms7090343 [ Links ]
Guevara, R.B., Lee, J.H., Lee, S.H., Seok, M.J., Kim, D.W., Kang, B.N., Johnson, T.J., Isaacson, R. & Kim, H.B., 2019. Piglet gut microbial shifts early in life: Causes and effects, J. Anim. Sci. Bio. 10, 1. doi.org/10.1186/s40104-018-0308-3 [ Links ]
Han, Y.S., Tang, C.H., Zhao, Q.Y., Zhan, J.F., Zhang, K., Han, Y.M. & Zhang, J.M., 2018. Effects of dietary supplementation with combinations of organic and medium chain fatty acids as replacements for chlortetracycline on growth performance, serum immunity, and fecal microbiota of weaned piglets. J. Liv. Sci. 216, 210-218. doi.org/10.1016/j.livsci.2018.08.013 [ Links ]
Hill, C., Guarner, F., Reid, G., Gibson, G.R., Merenstein, D.J., Pot, B., Moreli, L., Canani, R. B., Flint, H.J., Salminen, S., Calder, P.C. & Sanders, M.E., 2014. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotics. Nat Rev Gastroenterol & Hepatol. 11, 506-514. https://doi.org/10.1038/nrgastro.2014.66 [ Links ]
Hu, J., Meyers, R.M., Dong. J., Panchakshari, R.A., Alt, F.W. & Frock, R.L., 2016. Detecting DNA double-stranded breaks in mammalian genomes by linear amplification mediated high-throughput genome-wide translocation sequencing, Nat Prot.11(5), 853-871. doi:10.3389/fmicb.2016.01727 [ Links ]
Kantas, D., Papatsiros, V.G., Tassis, P.D., Giavasis, I., Bouki, P. & Tzika, E.D., 2014. A feed additive containing Bacillus toyonensis (Toyocerin) protects against enteric pathogens in post-weaning piglets. J. Appl. Microbiol. 1364-5072. doi:10.1111/jam.12729 [ Links ]
Kawai, T., Sato, S., Ishii, K., Coban, C., Hemmi, H., Yamamoto, M., Terai, K., Matsuda, M., Inoue, J., Uematsu, S., Takeuchi, O., Akira, S., 2004. Interferon-a induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat Immunol. 5, 1061-1068. doi.org/10.1038/ni1118 [ Links ]
Kil, D.Y., Kwon, W.B., Kim, B.G., 2011. Dietary acidifiers in weanling pig diets: a review, Rev Colomb Cienc Pecu, 24:231-247. http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-06902011000300002&lng=en&nrm=iso [ Links ]
Kim, Y.Y., Kil, D.Y., Oh, H.K. & Han, In.K., 2005. Acidifier as an alternative material to antibiotics in animal feed. Asian-Aust J. Anim. Sci. 1048-1060. doi.org/10.5713/ajas.2005.1048 [ Links ]
Kinoshita, K., Habermann, B. & Hyman, A.A., 2002. XMAP215: a key component of the dynamic microtubule cytoskeleton. SGD. 12, 267-273. doi.org/10.1016/S0962-8924(02)02295-X [ Links ]
Kraler, M., Ghanbari, M., Domig, K.J., Schedle, K. & Kneifel, W., 2016. The intestinal microbiota of piglets fed with wheat bran variants as characterised by 16S rRNA next generation amplicon sequencing. Arch Anim Nutr. 70, 173-189. doi:10.1080/1745039X.2016.1160534 [ Links ]
Konstantinov, S.R., Awati, A., Smidt, H.S., Williams, B.A., Akkermans, A.D.I. & De Vos, W.M., 2004. Specific response of a novel and abundant Lactobacillus amylovorus-like phylotype to dietary prebiotics in the guts of weaning piglets, Appl Environ Microbiol. 70, 3821-3830. doi.org/10.1128/AEM.70.7.3821-3830.2004 [ Links ]
Long, S.F., Xu, T.Y., Pan, Q.Q., Wang, C.L., Wang, J.Y., Wu, Y.Y., Han, Y.M., Yu, C.H. & Piao, X.S., 2018. Mixed organic acids as antibiotics substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglet. An. Feed Science Tech. 235, 23-32. doi.org/10.1016/janifeedsci.2017.08.018 [ Links ]
Lalles, J.P., 2016. Microbiota-host interplay at the gut epithelial level, health and nutrition. J. Anim. Sci. Biotech. 7, 66. doi.org/10.1186/s40104-016-0123-7 [ Links ]
Li, Y., Guo, Y., Wen, Z., Jiang, X., Ma, X. & Han, X., 2018. Weaning stress perturbs gut microbiome and its metabolic profile in piglets, Sci. Rep. 8, 18063. DOI:10.10381s41598-018-33649-8 [ Links ]
Li, K., Xiao, Y., Chen, J., Chen, J2., He, X. & Yang, H., 2017. Microbial in different locations of weaning piglets receiving antibiotics, Asian-Australas J. Anim. Sci. 30(1), 78-84. doi.org/10.5713/ajas.16.0285 [ Links ]
Luise, D., Bertocchi, M., Motta, V., Salvarani, C., Bosi, P., Luppi, A., Fanelli, F., Mazzoni, M., Archetti, I., Maiorano, G., Nielsen B.K.K., Trevisi, P., 2019. Bacillus sp. probiotic supplementation diminish the Escherichia Coli F4ac infection in susceptible weaned pigs by influencing the intestinal immune response, intestinal microbiota, and blood metabolomics. J. Anim Sci. 10, 174. doi.org/10.1186/s40104-019-0380-3 [ Links ]
McKenzie, C., Tan, J., Macia, L. & MacKay C.R., 2017. The nutrition-gut microbiome-physiology axis and allergic diseases, Immunol. Rev.1, 277-295. doi:10.1111/imr.12556 [ Links ]
Metzler-Zebeli, B.U., Lawlor, P.G., Magowan, E. & Zebeli, Q., 2018. Interactions between metabolically active bacteria and host gene expression at the cecal mucosa in pigs of diverging feed efficiency, J. Anim. Sci. 96, 2249-2264. doi:10.1093/jas/sky118 [ Links ]
Molist, F., Hermes, R.G., Segura, A.G., Martin-Orue, S.M., Gasa, J., Manzanilla, E.G. & Perez, J.F., 2011. Effect and interaction between wheat bran and zinc oxide on productive performance and intestinal health in post-weaning piglets. Br. J. Nutr. 105, 11592-1600. doi:10.1017/s0007114510004575 [ Links ]
Molist, F., de Segura, A.G., Perez, J.F., Bhandari, S.K., Krause, D.O. & Nyachoti, M. 2010. Effect of wheat bran on the health and performance of weaned pigs challenged with Escherichia coli K88+, Livest. Sci. 133(1), 214-217. doi.org/10.1016/j.livsci.2010.06.067 [ Links ]
Nemcova, R., Bomba, A., Gancarcikova, S., Reiffova, K., Guba, P., Kascova, J., Jonecova, Z., Scirankova, L., Bugarsky, A., 2007. Effects of the administration of Lactobacilli, maltodextrins, and fructooligosaccharides upon the adhesion of E. Coli 08:K88 to the intestinal mucosa and organic acid levels in the gut contents of piglets. Vet Res. Comm. 31, 791-800. DOI:10 1126/science. 1223813 [ Links ]
Nicholson, J.K., Holmes, E., Kinross, J., Burcelin, R., Gibson, G., Jia, W. & Patterson, S., 2012. Host-gut microbiota metabolic interactions. Sci. 336, 1262-7. doi:10.1126/science. 1223813 [ Links ]
Rakoff-Nahoum, S., Paglino, J., Eslami-Varzaneh, F., Edberg, S. & Medzhitow, R., 2004. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell. 118(2), 1229-41. doi:10.1016/j.cell.2004.07.002 [ Links ]
Ridlon, J., Kang, D. & Hylemon, P., 2006. Bile salt transformations by human intestinal bacteria. J. Lipid Res. 47, 241-259. doi.org/10.1194/jlr.R500013-JLR200 [ Links ]
Roth, F.X., Kirchgessner, M., 1998. Organic acids as feed additives for young pigs: Nutritional and gastrointestinal effects. J. Anim & Feed Sci, (Suppl. 1), 25-33. doi.org/1022358/jafs/69953/1998 [ Links ]
Sun, M., Wu, W., Liu, Z. & Cong, Y., 2017. Microbiota metabolite short chain fatty acids, GPR, and inflammatory bowel diseases, J. Gastroenterol. 252, 1-8. doi:10.1007/S00535-016-1242-9 [ Links ]
Trevisi, P., Priori, D., Jansman, A.J.M., Luise, D., Koopsmans, S. & Hynonen, U., 2018. Molecular networks affected neonatal microbial colonization in porcine jejunum, luminally perfused with enterotoxigenic Escherichia Coli F4ac fimbria or Lactobacillus amylovorus, PLoS One. 13, 0202160. doi:10.1371/journal.pone.0202160 [ Links ]
Tsukahara, T., Iwasaki, Y., Nakayama, K. & Ushida, K., 2003. Stimulation of butyrate production in the large intestine of weaning piglets by dietary fructooligossacharides and its influence on the histological variables of the large intestinal mucosa. J. Nutri Sci Vitaminol. 49, 414-421. doi:10.3177/jnsv.49.414 [ Links ]
Wang, T., Teng, K., Liu, Y., Shi, W., Zhang, J., Dong, E., Zhang, X., Tao, Y. & Zhong, J., 2019. Lactobacillus plantarum PFM 105 promotes intestinal development through modulation of gut microbiota in weaning piglets Front Microbiol. 10, 90. doi:10.3389/fmicb.2019.00090 [ Links ]
Watt, R., Parkin, K. & Martino, D., 2020. The potential effects of SCFAs on the epigenetic regulation of innate immune memory. Challenges. 11, 25. doi:10.3390/challe11020025 [ Links ]
Zhao, W., Yuan, M., Li, P., Yan, H., Zhang, H. & Liu, J., 2019. Short-chain fructo-oligossacharides enhances intestinal barrier function by attenuating mucosa inflammation and altering colonic microbiota composition of weaning piglets. It./. J. Anim. Sci. 18, 1, 976-986. doi.org/10.1080/1828051X.2019.1612286 [ Links ]
Zhao, Y., Wang, J., Wang, H., Huang, Y., Qi, M., Liao, S., Bin, P. & Yin, Y., 2020. Effects of GABA supplementation on intestinal SIgA secretion and gut microbiota in healthy and ETEC-infected weanling piglets. Hindawu. 7368483, 17. doi.org/10.1155/2020/7368483 [ Links ]
Submitted 6 September 2022
Accepted 9 January 2023
Published 14 May 2023
# Corresponding author: L. Dong, E-mail: donglijiayou@126.com


{kind=link}
{kind=link}