










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 no.6 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i6.14
Effect of microRNA, miR-215, on gene expression and metabolism of porcine intestinal epithelial cells
Hongyi LiI, II; Mao ZhangI, #; Wenjuan LiuIII
IHenry Fok College of Biology and Agriculture, Shaoguan University, Guangdong,China, 512005
IICollege of Life Science, Longyan University, Fujian, 364012, China
IIIShenzhen Kingsino Technology Co., Ltd, Guangdong, China, 518107
ABSTRACT
MiR-215 maintains a high level of expression in healthy pig intestine and is decreased in lesions or injuries. To clarify the influence of miR-215 on the porcine intestine, miR-215 mimic/inhibitor was transfected into the intestinal porcine epithelial cell line, J2 (IPEC-J2) to overexpress or suppress the expression of miR-215. The treated cells were subjected to various functional assays. The effect of miR-215 on global gene expression was detected using high-throughput sequencing on the BGISEQ-500 platform; cell proliferation was evaluated using a Cell Counting Kit-8 (CCK-8); and the role of miR-215 on cellular metabolism was tested using non-targeted metabonomics and liquid chromatography with tandem mass spectrometry (LC-MS/MS) analysis. Our results revealed that miR-215 promoted cell proliferation in IPEC-J2, up-regulated the expression of the ribosomal protein family, and down-regulated the expression of oncogenesis-related genes and tumour-suppressor genes. Results of the RNA sequencing and the metabonomics detection reveal that miR-215 affects the PI3K-Akt signalling pathway. Our results contribute to further research on miR-215 in the regulation of barrier function and metabolism in the porcine intestine.
Keywords: PI3K-Akt, proliferation, RNA-Sequencing
Introduction
Pigs are considered to be ideal biomedical models for research because of their many physiological similarities with humans (Sharbati et al., 2010). The intestine is one of the most architecturally and functional organs in mammals. It plays a key role in digesting and absorbing nutrients, as well as eliminating waste and protecting from infection (Du P et al., 2016). It is also the fastest self-renewing tissue in the body (Zou et al., 2018). Therefore, keeping intestinal function normal is essential for the maintenance of health.
MicroRNAs (miRNAs) are small, non-coding RNAs of 19-25 nucleotides in length. They play a key role in post-transcription by binding to the 3'-untranslated region (3'UTR) of target mRNA (Bartel, 2009). Recent studies describe the crucial role of miRNAs in cell damage repair (Bhanja et al., 2018), cell self-renewal (Andriatsilavo et al., 2018), nutrient metabolism (Lesjak et al., 2014), and immune regulation (Sawant et al., 2013; Murer et al., 2019). Our previous studies utilizing pig models revealed substantial differences in miRNA expression between an Lawsonia intracellularis-infected porcine intestine and the control pigs, with miR-215 expression being 49.4-fold lower than that of control group (Li et al., 2017).
Results of recent reports show that miR-215 is related to the pathogenesis of multiple diseases in multiple tissues. In papillary thyroid cancer (PTC), miR-215 inhibits PTC proliferation and metastasis by targeting ARFGEF1 (Han et al., 2019); in multiple myeloma (MM), miR-215-5p inhibits MM cell apoptosis via targeting RUNX1 and suppressing the activation of the PI3K/AKT/mTOR pathway (Liu et al., 2020); and miR-215 promotes cell migration and invasion of gastric cancer by directly targeting RB1 (Chen et al., 2017). In the intestine, miR-215 was found to affect neuron migration and proliferation through the target gene, SIGLEC-8, in Hirschsprung's disease (Lei et al., 2016); it also suppressed colon cancer cell proliferation, migration, and invasion by directly targeting YY1 (Chen et al., 2016), and worked against the effects of hypoxia in colon tumour-initiating cells (Ullmann et al., 2019).
Interestingly, expression levels of miR-215 in the above reported tumours were substantially lower than in normal tissues, similar to our previous report in pigs. Coincidentally, miR-215 is also expressed less in the small intestine of weaned piglets than in normal suckling piglets (Tao & Xu, 2013). To investigate whether the expression of miR-215 was reduced in pigs with intestinal injury or structural changes, we used two existing models to detect miR-215 expression levels and discovered that miR-215 was highly expressed in normal intestinal tissues, indicating that miR-215 plays an important role in the regulation of the porcine intestine. In order to clarify the influence of miR-215 on the porcine intestine, we focused on the role of miR-215 in regulating intestinal metabolism and global gene expression in this study to elucidate the regulatory role of miR-215.
Materials and Methods
The protocol for the research project was approved by the guidelines of Guangdong Province on the Review of Welfare and Ethics of Laboratory Animals, which was approved by the Guangdong Province Administration Office of Laboratory Animals (GPAOLA, 2020-llsc-004).
Model of piglets with diarrhoea and spleen deficiency: Intestinal tissue of piglets with diarrhoea and spleen deficiency was kept by the laboratory. The model was established according to the diagnostic reference standard of spleen deficiency syndrome in traditional Chinese medicine, using the method of cervical intramuscular injection of reserpine and abnormal hunger and satiety. Twenty, healthy 25-day-old, weaned piglets (Duroc χ Landrace χ Yorkshire) with similar body weight were randomly divided into two groups, each group with 10 piglets. The experimental group was injected with reserpine (0.10 mL/kg) every morning and alternately receiving no feeding or full feeding every other day. The control group was injected with 0.10 mL normal saline/kg every day. After three days, the experimental piglets had loose stools, followed by symptoms such as limb weakness, reduced activity, withered or erect hair, poor colour, supine, sleepy, slow movements, and slow responses, indicating that the spleen deficiency model was successfully copied.
Cell line and cell culture: Intestinal porcine epithelial cell J2 (IPEC-J2) were preserved in our laboratory. Cells were maintained in Dulbecco's Modified Essential Medium: Nutrient Mixture F-12 (DMEM/12, GBICO, Grand Island, New York, USA) with 10% foetal bovine serum (FBS, GBICO), 100 U penicillin/mL, and 100 mg streptomycin/mL, and cultured in a humidified atmosphere with 5% CO2 at 37 °C.
RNA sequencing and analysis: Cell lines transfected with the miR-215 mimic or the negative control sequence (NC, GenePharma, Nanjing, China) were seeded into a 6-well plate in triplicate at a concentration of 5 *105 cells per well. After 48 h, cells were collected for total RNA extraction. Library construction and sequencing were performed on a BGISEQ-500 platform by Beijing Genomic Institution (www.genomics.org.cn, BGI, Shenzhen, China). Clean reads were mapped to the reference genome and available genes using HISAT (Kim et al., 2015). For gene expression analysis, the mapping fragments were calculated and normalized to FPKM (fragments per kilobase per million mapped fragments) using RESM software (Li & Dewey, 2011). Pearson's correlation coefficient was used to measure the repeatability between parallel samples. The significance of the differential expression of genes (DEGs) was defined according to the combination of the absolute value of log2-ratio (miR-215/control) >1, and false discovery rate (FDR) <0.01. Hierarchical cluster analysis of differentially expressed genes was performed with the heatmap function in the R package. Genes with the same expression pattern among samples were clustered using the Mfuzz package. The classification and enrichment of the gene function and pathway annotation were performed based on the Gene Ontology (GO) Database (http://www.geneontology.org/) and KEGG (Kyoto Encyclopedia of Genes and Genomes) pathway database (http://www.genome.jp/kegg/), respectively.
Cell proliferation assay: While transfected with the miR-215 mimic or inhibitor, the IPEC-J2 cell line was seeded and incubated in a 96-well plate (4 χ 103 cells per well) for 24 h, 48 h, 72 h, and 96 h. At the indicated time, cell proliferation activity of cell was examined using a Cell Counting Kit-8 (CCK8, Beyotime, Shanghai, China) in accordance with the manufacturer's instructions. In brief, cells were added to 20 μL CCK-8 solution to incubate for 2h, and the absorbance of the cells at 450 nm was examined using a microplate reader (Bio-Tek, USA).
Metabonomics preparation and detection: Cells were seeded into 6-well plates at a concentration of 5 χ 105 cells per well, and transfected with miR-215 mimic and NC respectively, with eight replicates each treatment. Cells were collected 48 h after transfection and stored at -80 °C until ultra-high performance liquid chromatography and mass spectroscopy (LC-MS/MS). Before LC-MS/MS analysis, the cells were left to thaw at 4 °C for approximately 30 min and an equal volume of frozen methanol and water (800 μL) was added. After fragmentation, cells were precipitated at -20 °C for 2 h and centrifuged at 4 °C for 15 min. An amount of 800 μL of supernatant was sent for further preparation and the remaining samples were reserved for future use. LC-MS/MS was performed using a UPLC system (Waters, Milford, Massachusetts, USA) coupled to a Xevo G2-XS quadrupole-time-of-flight spectrometer (Waters). For liquid chromatography, samples were analysed using a 2.1 mm χ 100 mm ACQUIY UPLC BEH 1.7-μm column at a flow rate of 400 μl/min, with water containing 25 mM ammonium acetate and 25 mM ammonium hydroxide as mobile phase A, while acetonitrile was used as mobile phase B in the gradient elution mode. The gradient composition was 95% of B for 0.5 min, linearly reduced to 65% in 7 min, then reduced to 40% in 2 min. After maintaining this condition for 1 min, it was increased to 95% in 1 min, with 3 min left for column re-equilibration. For mass spectrometry, data were acquired in positive and negative modes. The electrospray ionisation source parameters were set as follows: source temperature of 150 °C, desolvation temperature of 450 °C, sampling cone of 40 V, cone gas flow of 50 L/h, and desolvation gas flow of 900 L/h. Capillary voltage was set to 3500 V for the positive mode and 2500 V for the negative mode. The data acquisition rate was set to 0.4 s with a 0.1 s interscan delay. All analyses were acquired using the lock spray to ensure accuracy and reproducibility. Additionally, quality control (QC) samples were prepared by pooling aliquots of all samples that were under analysis to use for data normalization and to ensure data quality for metabolic profiling. QC samples were prepared and analysed using the same procedure as that for the experimental samples.
LC-MS/MS data processing: Peak detection, alignment, deconvolution, data filtering, and ion annotation were performed using Progenesis QI (Waters), as previously reported (Franco et al., 2020). Metabolites with a P-value less than 0.05 were considered to be statistically significant. Fold change was calculated as the logarithm of the average mass area ratio between the treatment group and the control. Furthermore, the identified differential metabolites were used to perform cluster analyses using the R software package. KEGG ID mapping was performed on the differential metabolites and submitted to the KEGG website for related pathway analysis. Enriched KEGG pathways were nominally statistically significant at P <0.05.
Statistics: Statistical analyses and drawing of figures were done using GraphPad Prism 5.0 software. All descriptive data were expressed as mean ± standard deviation (SD) and the significance of difference was calculated using paired t-tests. Values of P <0.05 were considered significant.
Results and Discussion
MiR-215 was highly expressed in normal intestinal tissues: In our previous report, the expression of miR-215 was 49.4-fold lower in Lawsonia intracellularis-infected porcine intestine (Li et al., 2017),which would cause porcine proliferative enteropathy (PPE). PPE is an intestinal hyperplastic disease characterized by the thickening of the intestinal mucosa due to enterocyte proliferation (Lawson & Gebhart, 2000). In order to verify whether miR-215 was down-regulated in the case of intestinal injury or structural changes, the expression of miR-215 was detected using the existing models of spleen-deficiency diarrhoea (using reserpine and poor feeding methods) piglets and a pAPN (porcine aminopeptidase N) knockout cell line. The results showed that miR-215 all maintained a high level of expression in the intestine of the control group (P <0.01, Figure 1). Diarrhoea due to a spleen deficiency can damage intestinal epithelial cells and change villus structure. IPEC-J2 with pAPN knockout also alters the structure of intestinal cells. From the report of Tao et al. (Tao & Xu, 2013), the expression of miR-215 was also found to be decreased in piglets with intestinal stress caused by weaning. Therefore, it is critical to study the comprehensive effects of miR-215 on intestinal gene expression.
MiR-215 affects gene expression in the pig intestine: miR-215 was highly expressed in normal intestinal tissues in the current study, indicating that miR-215 may play a critical role in intestinal health. To identify the effects of miR-215 on the comprehensive expression profile in the porcine intestine, the miR-215 mimics was transfected into the IPEC-J2 cell and RNA-sequencing (RNA-Seq) was conducted. The correlation coefficients of the three biological replicates in the experimental and the NC groups were all above 0.99 (Figure 2A), indicating that the data has excellent repeatability and were suitable for the subsequent analysis. The correlation coefficients of gene expression between transfected miR-215 and the NC cell were in the range of 0.97 to 0.99, indicating that miR-215 altered the expression of just a small number of genes.
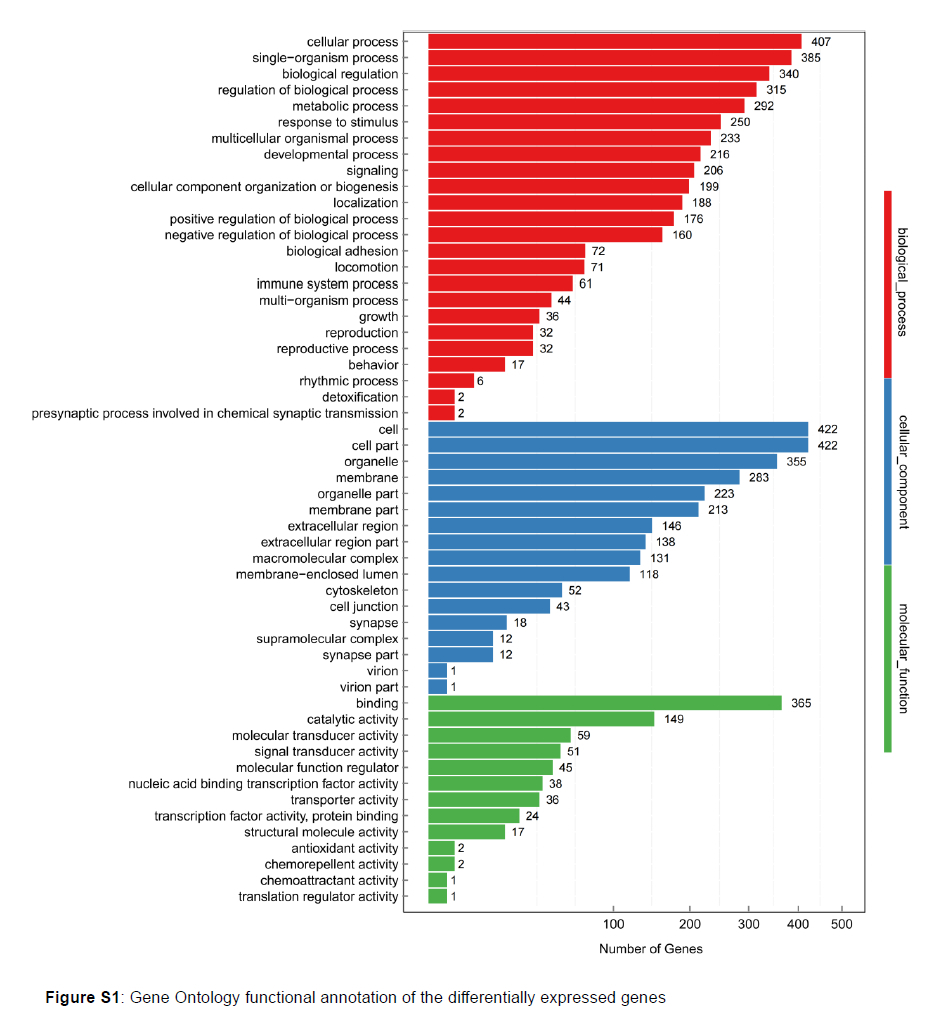
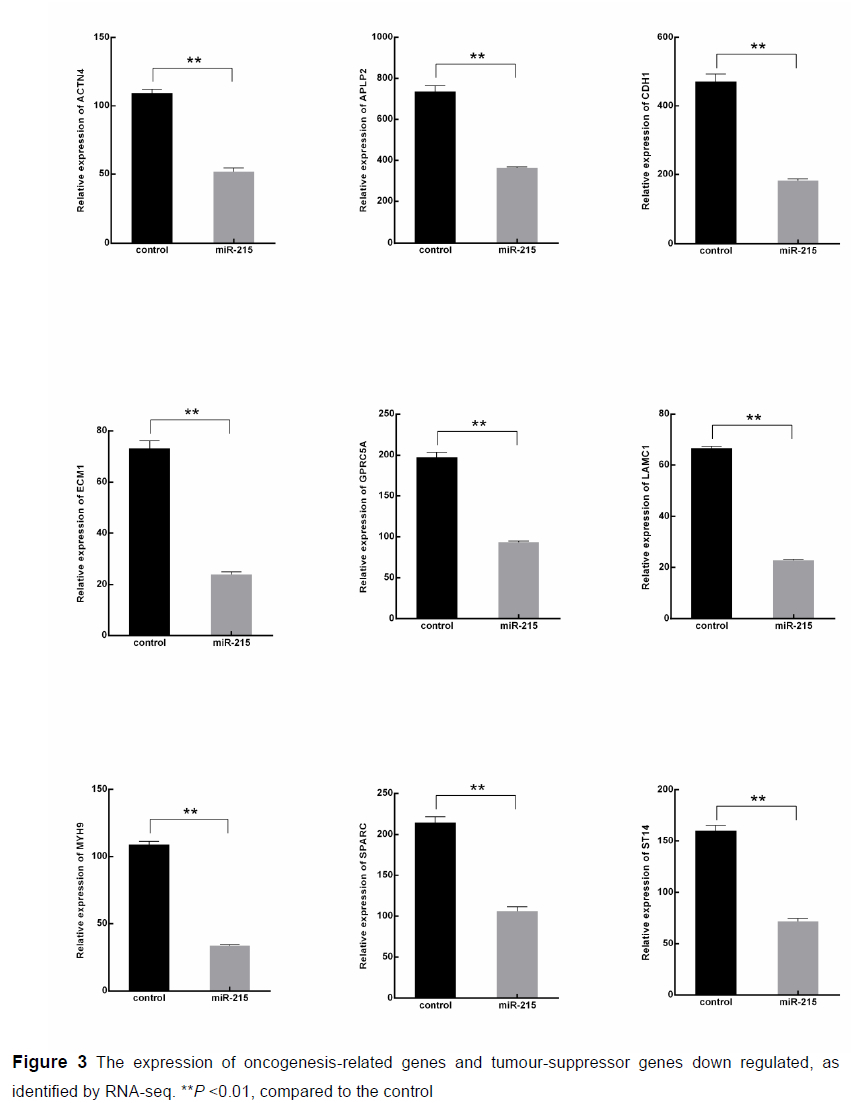
An average of 23.89 million clean reads were obtained that mapped on the porcine genome at an average rate of 94.80%, representing an average of 20349 genes that were expressed in each sample. From the mapping genes, we found that miR-215 decreased the quantity of high expressed genes (FPKM >10, Figure 2B). Compared with the NC group, a total of 31 up-regulated genes and 685 down-regulated genes were detected in the miR-215 transfected cells (Figure 2C). GO functional annotation of the DEGs indicated that the up- and down-regulated genes could be classified into three parts of 54 categories, such as cellular processes, metabolic processes, cell parts, and binding (Figure S1). From the DEGs, we found that the expression of oncogenesis-related genes, such as myosin-9 (MYH9), actinin alpha 4 (ACTN4), secreted protein, acidic and rich in cysteine (SPARC), mucin-4 (MUC4) were down regulated in the miR-215 over-expressed cells (P <0.01, Figure 3). What interested us was that the expression of the tumour-suppressor genes, such as suppression of tumorigenicity 14 (ST14), were also down-regulated, indicating that moderately high expression of miR-215 may reduce cell tumorigenicity.
Pathway enrichment analysis of DEGs was conducted according to KEGG functional annotation to identify signalling pathways possibly regulated by miR-215-5p in IPEC-J2. We found that the DEGs were focussed on the PI3K-Akt signalling pathway, focal adhesion, ECM-receptor interaction, pathways in cancers, inter alia (Figure 4).
MiR-215 up-regulated the expression of the ribosomal protein (RP) family and promoted cell proliferation: Detected genes with the same expression pattern among samples were clustered using Mfuzz software, and one of the clusters caught our attention. The membership of most genes in this cluster was close to 1, which showed that the trend of the gene was consistent with the change of the cluster (Figure 5). In this cluster, we found most members of the RP family were 1.5-2.0-fold up-regulated in the miR-215 transfected group (Table S1), indicating that miR-215 may be related to the proliferation of porcine intestinal cells. The CCK-8 assay was performed to detect the effect of miR-215 on the cell proliferation. Twenty-four hours after treatment, there was no significant difference among the miR-215 overexpressed group, miR-215 suppressed group, and the control group (P >0.05). The miR-215 mimics began to promote cell proliferation from 48 h (P <0.05) and the miR-215 inhibitor had the opposite effect (P <0.05), which confirmed our hypothesis that miR-215 promoted cell proliferation in IPEC-J2 (Figure 6).
Recent research indicates that the expression of miR-215 decreases in almost all cancer tissues, except in some tumour cells (Lei et al., 2016; Wang et al., 2017) and miR-215 inhibits cell proliferation in most of cancer cells (Cai et al., 2017; A. Liu et al., 2020; Wu et al., 2020; X. Xu et al., 2020). In our results, miR-215 promoted IPEC-J2 cell proliferation, which may indicate that miR-215 will accelerate the self-renewal of porcine intestinal cells by promoting cell proliferation in normal conditions and may inhibit cell progression through self-feedback in the case of abnormal proliferation, thereby inhibiting further deterioration of the tumour. The acceleration of cell proliferation will lead to an increase in protein synthesis. Most cellular proteins are manufactured on ribosomes in the cytoplasm. Therefore, the expression of ribosome family genes almost indicates an upward trend to adapt to the increase in protein synthesis.
MiR-215 affects intestinal cell metabolism in pigs: GO functional annotation of the DEGs indicated that miR-215 also regulated the metabolic process. To elucidate the regulation of miR-215 of the metabolism of pig intestinal cells, the metabolomics were also analysed between the miR-215-treated cells and the control cells. A total of 5223 and 4682 ions were detected in positive and negative modes, respectively, of which the ion numbers of relative standard deviation (RSD) less than 30% were 3093 and 2772, including 160 and 205 differentially expressed ions, respectively. On the basis of the KEGG pathway database, perturbed biological pathways between miR-215-mimic transfected cells and the control cells were identified. Differential expression of ions was mainly concentrated in 27 pathways and most of them were involved in glucose, amino acid, and nucleic acid metabolism. What interested us was that the test result was consistent with the sequencing; the differential metabolites were the same affected by the PI3K-Akt signalling pathway (Figure 7). The PI3K-AKT signalling pathways are crucial to many physiological and pathological conditions, such as cell proliferation, angiogenesis, metabolism, differentiation, and survival (Porta et al., 2014). Current research indicates that PI3K-AKT is associated with various cancers, such as neurodegenerative tumours (Xu et al., 2020), prostate cancer (Shorning et al., 2020), non-small cell lung cancer (Tan, 2020), breast cancer (Miricescu et al., 2020). MiRNAs were also involved in the regulation process: among them, PI3K-Akt can negatively regulate miR-192/215 expression in colon cancer (Zhao et al., 2018). MiR-215 can target EREG, NIPAL1, and PTPRU genes to regulate the resistance of piglets to Escherichia coli F18 (Dai et al., 2020), indicating that besides cancer, miR-215 also plays an important regulatory role in the intestinal tract of pigs.
Conclusions
In conclusion, this study revealed that miR-215 promoted cell proliferation in IPEC-J2 and caused a series of changes in gene expression and metabolites. Both transcriptome sequencing and metabonomic detection indicated that miR-215 may regulate porcine intestinal function via the PI3K/AKT pathway. However, the detailed regulatory mechanism of miR-215 in regulating the porcine intestine via the PI3K/AKT pathway requires further investigation.
Acknowledgements
This work was supported by Guangdong Basic and Applied Basic Research Foundation (Grant No. 2021A1515010483), the Doctoral Initiation Fund of Shaoguan University (Grant No.432-99000633), the University Outstanding Young Scientific Research Talents Program of Fujian province (2018), and the Project of Guangdong Provincial Department of Education (2021ZDZX4040).
Author contributions
Hongyi Li contributed to experimental operation and data analysis; Mao Zhang contributed to experimental design and article writing; Wenjuan Liu contributed to graphics production.
Conflict of interest declaration
Authors declare no conflict of interests for this article.
References
Andriatsilavo, M., Stefanutti, M., Siudeja, K., Perdigoto, C. N., Boumard, B., Gervais, L., Gillet-Markowska, A., Al, Z. L., Schweisguth, F., & Bardin, A. J. 2018. Spen limits intestinal stem cell self-renewal. Plos Genet, 14(11), e1007773. doi 10.1371/journal.pgen.1007773 [ Links ]
Bartel, D. P. 2009. MicroRNAs: Target recognition and regulatory functions. Cell, 136(2), 215-233. doi 10.1016/j.cell.2009.01.002 [ Links ]
Bhanja, P., Norris, A., Gupta-Saraf, P., Hoover, A., & Saha, S. 2018. BCN057 induces intestinal stem cell repair and mitigates radiation-induced intestinal injury. Stem Cell Res Ther, 9(1), 26. doi 10.1186/s13287-017-0763-3 [ Links ]
Cai, X., Peng, D., Wei, H., Yang, X., Huang, Q., Lin, Z., Xu, W., Qian, M., Yang, C., Liu, T., Yan, W., & Zhao, J. 2017. MiR-215 suppresses proliferation and migration of non-small cell lung cancer cells. Oncol Lett, 13(4), 2349-2353. doi 10.3892/ol.2017.5692 [ Links ]
Chen, Z., Han, S., Huang, W., Wu, J., Liu, Y., Cai, S., He, Y., Wu, S., & Song, W. 2016. MicroRNA-215 suppresses cell proliferation, migration and invasion of colon cancer by repressing Yin-Yang 1. Biochem Biophys Res Commun, 479(3), 482-488. doi 10.1016/j.bbrc.2016.09.089 [ Links ]
Chen, Z., Liu, K., Li, L., Chen, Y., & Du S. 2017. miR-215 promotes cell migration and invasion of gastric cancer by targeting Retinoblastoma tumor suppressor gene 1. Pathol. Res. Pract., 213(8), 889-894. doi 10.1016/j.prp.2017.06.006 [ Links ]
Dai, C. H., Wang, F., Wang, S. Q., Wu, Z. C., Wu, S. L., & Bao, W. B. 2020. miR-215 targeting novel genes EREG, NIPAL1, and PTPRU regulates the resistance to E. coli F18 in piglets. Genes (Basel), 11(9). doi 10.3390/genes11091053 [ Links ]
Du P, Paskaranandavadivel, N., Angeli, T. R., Cheng, L. K., & O'Grady, G. 2016. The virtual intestine: In silico modeling of small intestinal electrophysiology and motility and the applications. Wiley Interdiscip Rev Syst Biol Med, 8(1), 69-85. doi 10.1002/wsbm.1324 [ Links ]
Franco, E., Contesini, F. J., Lima, D. S. B., Alves, D. P. F. A., Wielewski, L. C., Goncalves, C. J., Bueno, C. P., & de Oliveira, C. P. 2020. Enzyme-assisted modification of flavonoids from Matricaria chamomilla: Antioxidant activity and inhibitory effect on digestive enzymes. J Enzyme Inhib Med Chem, 35(1), 42-49. doi 10.1080/14756366.2019.1681989 [ Links ]
Han, J., Zhang, M., Nie, C., Jia, J., Wang, F., Yu, J., Bi, W., Liu, B., Sheng, R., He, G., Kong, L., Zheng, L., Pang, R., Ding, Z., Chen, L., Guan, Q., Pan, S., Meng, X., Xu, J., Liu, L., & Zhang, J. 2019. miR-215 suppresses papillary thyroid cancer proliferation, migration, and invasion through the AKT/GSK-3beta/Snail signaling by targeting ARFGEF1. Cell Death Dis, 10(3), 195. doi 10.1038/s41419-019-1444-1 [ Links ]
Kim, D., Langmead, B., & Salzberg, S. L. 2015. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods, 12(4), 357-360. doi 10.1038/nmeth.3317 [ Links ]
Lawson, G. H., & Gebhart, C. J. 2000. Proliferative enteropathy. J. Comp. Pathol., 122(2-3), 77-100. doi 10.1053/jcpa.1999.0347 [ Links ]
Lei, H., Li, H., Xie, H., Du C, Xia, Y., & Tang, W. 2016. Role of MiR-215 in Hirschsprung's Disease pathogenesis by targeting SIGLEC-8. Cell. Physiol. Biochem., 40(6), 1646-1655. doi 10.1159/000453214 [ Links ]
Lesjak, M., Hoque, R., Balesaria, S., Skinner, V., Debnam, E. S., Srai, S. K., & Sharp, P. A. 2014. Quercetin inhibits intestinal iron absorption and ferroportin transporter expression in vivo and in vitro. Plos One, 9(7), e102900. doi 10.1371/journal.pone.0102900 [ Links ]
Li, B., & Dewey, C. N. 2011. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics, 12, 323. doi 10.1186/1471-2105-12-323 [ Links ]
Li, H., Zhang, M., & Zheng, E. 2017. Comprehensive miRNA expression profiles in the ilea of Lawsonia intracellularis-infected pigs. J. Vet. Med. Sci., 79(2), 282-289. doi 10.1292/jvms.16-0423 [ Links ]
Liu, A., Zhang, B., Zhao, W., Tu, Y., Wang, Q., & Li, J. 2020. MicroRNA-215-5p inhibits the proliferation of keratinocytes and alleviates psoriasis-like inflammation by negatively regulating DYRK1A and its downstream signalling pathways. Exp. Dermatol. doi 10.1111/exd.14188 [ Links ]
Liu, S., Zhang, Y., Huang, C., & Lin, S. 2020. miR-215-5p is an anticancer gene in multiple myeloma by targeting RUNX1 and deactivating the PI3K/AKT/mTOR pathway. J. Cell. Biochem., 121(2), 1475-1490. doi10.1002/jcb.29383 [ Links ]
Miricescu, D., Totan, A., Stanescu-Spinu, I. I., Badoiu, S. C., Stefani, C., & Greabu, M. 2020. PI3K/AKT/mTOR Signaling pathway in breast cancer: From Molecular landscape to clinical aspects. Int. J. Mol. Sci., 22(1). doi 10.3390/ijms22010173 [ Links ]
Murer, A., Ruhl, J., Zbinden, A., Capaul, R., Hammerschmidt, W., Chijioke, O., & Munz, C. 2019. MicroRNAs of Epstein-Barr Virus attenuate T-cell-mediated immune control in vivo. Mbio, 10(1). doi 10.1128/mBio.01941-18 [ Links ]
Porta, C., Paglino, C., & Mosca, A. 2014. Targeting PI3K/Akt/mTOR signaling in cancer. Front Oncol, 4, 64. doi 10.3389/fonc.2014.00064 [ Links ]
Sawant, D. V., Wu, H., Kaplan, M. H., & Dent, A. L. 2013. The Bcl6 target gene: MicroRNA-21 promotes Th2 differentiation by a T cell intrinsic pathway. Mol. Immunol., 54(3-4), 435-442. doi 10.1016/j.molimm.2013.01.006 [ Links ]
Sharbati, S., Friedlander, M. R., Sharbati, J., Hoeke, L., Chen, W., Keller, A., Stahler, P. F., Rajewsky, N., & Einspanier, R. 2010. Deciphering the porcine intestinal microRNA transcriptome. BMC Genomics, 11, 275. doi 10.1186/1471-2164-11-275 [ Links ]
Shorning, B. Y., Dass, M. S., Smalley, M. J., & Pearson, H. B. 2020. The PI3K-AKT-mTOR Pathway and prostate cancer: At the Crossroads of AR, MAPK, and WNT signaling. Int. J. Mol. Sci., 21(12). doi 10.3390/ijms21124507 [ Links ]
Tan, A. C. 2020. Targeting the PI3K/Akt/mTOR pathway in non-small cell lung cancer (NSCLC). Thorac Cancer, 11(3), 511-518. doi 10.1111/1759-7714.13328 [ Links ]
Tao, X., & Xu, Z. 2013. MicroRNA transcriptome in swine small intestine during weaning stress. Plos One, 8(11), e79343. doi 10.1371/journal.pone.0079343 [ Links ]
Ullmann, P., Nurmik, M., Schmitz, M., Rodriguez, F., Weiler, J., Qureshi-Baig, K., Felten, P., Nazarov, P. V., Nicot, N., Zuegel, N., Haan, S., & Letellier, E. 2019. Tumor suppressor miR-215 counteracts hypoxia-induced colon cancer stem cell activity. Cancer Lett., 450, 32-41. doi 10.1016/j.canlet.2019.02.030 [ Links ]
Wang, C., Chen, Q., Li, S., Li, S., Zhao, Z., Gao, H., Wang, X., Li, B., Zhang, W., Yuan, Y., Ming, L., He, H., Tao, B., & Zhong, J. 2017. Dual inhibition of PCDH9 expression by miR-215-5p up-regulation in gliomas. Oncotarget, 8(6), 10287-10297. doi 10.18632/oncotarget.14396 [ Links ]
Wu, R. J., Zou, Y., Ma, X. D., & Zheng, R. J. 2020. [TL - Expression and clinical significance of MiR-215 and KDM1B in Patients with diffuse large B cell lymphoma]. Zhongguo Shi Yan Xue Ye Xue Za Zhi, 28(5), 1570-1577. doi 10.19746/j.cnki.issn.1009-2137.2020.05.024 [ Links ]
Xu, F., Na, L., Li, Y., & Chen, L. 2020. Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci, 10, 54. doi 10.1186/s13578-020-00416-0 [ Links ]
Xu, X., Ding, Y., Yao, J., Wei, Z., Jin, H., Chen, C., Feng, J., & Ying, R. 2020. miR-215 inhibits colorectal cancer cell migration and invasion via targeting stearoyl-CoA desaturase. Comput Math Methods Med, 2020, 5807836. doi 10.1155/2020/5807836 [ Links ]
Zhao, J., Xu, J., & Zhang, R. 2018. SRPX2 regulates colon cancer cell metabolism by miR-192/215 via PI3K-Akt. Am J. Transl Res, 10(2), 483-490. doi [ Links ]
Zou, L., Xiong, X., Wang, K., & Yin, Y. 2018. MicroRNAs in the Intestine: Role in renewal, homeostasis, and inflammation. Curr. Mol. Med., 18(3), 190-198. doi 10.2174/1566524018666180907163638 [ Links ]
Submitted 3 November 2021
Accepted 27 October 2022
Published 5 March 2023
# Corresponding author: zm18email@163.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}