










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 n.6 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i6.13
Crambe cake protein as a replacement for soybean meal protein: Intake, digestibility, and ruminal parameters in lamb diets
A.R. Poveda-ParraI, #; E. S. PereiraII; O. P. Prado-CalixtoI; G. L. GuerraI; K. A. PiresI; M. R. LopesI; E. R. RodriguesI; E. R. Cavalheiro JuniorI; J. P. M. do CarmoIII; I. Y. MizubutiI
IDepartment of Animal Science, State University of Londrina, Londrina, Paraná, Brazil
IIDepartment of Animal Science, Federal University of Ceará, Fortaleza, Ceará, Brazil
IIIDepartment of Medicine Veterinary, State University of Londrina, Londrina, Paraná, Brazil
ABSTRACT
The objective was to evaluate different levels of crambe cake protein (0, 250, 500, 750 and 1000 g/kg of DM) to replace soybean meal protein on the intake, apparent digestibility coefficient (ADC), blood glucose, ruminal fermentation, and protozoal population in lambs. Two experiments were performed. In experiment one, 40 crossbred male lambs weighing 20 ± 3.45 kg were used, distributed in a completely randomized design, to study intake and ruminal parameters. In experiment two, 25 crossbred male lambs weighing 23.44 ± 1.04 kg were used to study digestibility. The intake of dry matter and total carbohydrate (TC) (kg/lambs/day and g/kg0.75), crude protein (kg/lambs/day) and final body weight decreased with increase of crambe cake protein as a replacement for soybean meal protein, while ether extract ( EE) intake (kg/lambs/day and g/kg075) increased. The ADC of EE presented a positive linear effect and the ADC of TC and non-fibrous carbohydrates presented a negative linear effect. There was no interaction within treatments and collection period (0, 3, 6 and 9 h) on ruminal fluid characteristics. Blood glucose presented a quadratic effect for levels of crambe cake protein in diets (max. P = 35.18%), and linear increase for collection period. Ruminal pH showed a quadratic effect for sampling hours (max. P = 5.43 h). For protozoa evaluation, the total amount, Entodinium, and Eudiplodium presented linear decreasing effects with treatment, while Isotricha, Eudiplodium, Ostracodinium and the total amount of protozoa had a linear decrease with sampling hours. Soybean meal protein can be replaced by crambe cake protein at up to 250 g/kg DM.
Key-words: by-product, Crambe abyssinica Hochst, nutritive value, protozoa, sheep
Introduction
The competition between humans and animals for grains threatens the sustainability of food systems in general, and it therefore becomes necessary to develop new ingredients, especially non-conventional ingredients, that can be used in animal feed (Yang et al., 2021). The renewable sources for biodiesel production in Brazil are soybean, cotton, maize grains, sunflower, canola, and animal fat (Ramos et al., 2017). However, some non-edible oleaginous plants have aroused interest as alternative raw materials, among which is crambe (Crambe abyssinica Hochst). In the production of biodiesel, some by-products are generated, which, although considered pollutants, are available and suitable for use in ruminant nutrition (Mendonça et al., 2015).
Crambe is a subtropical plant, native to the Mediterranean, tolerant to drought and acid soils, with grains that have a high protein content (460 to 580 g/kg), energy (5994 kcal/g) and ether extract (260 to 440 g/kg) (Souza et al., 2009; Goes et al., 2010; Goes et al., 2016). During the extraction of the oil, by-products such as crambe cake and bran are obtained.
Crambe cake is obtained after the mechanical extraction of the oil by pressing the seeds, presenting a residual oil content higher than that contained in the crambe meal (Canova, 2015). The crambe cake contains 870-940 g/kg dry matter, 240-340 g/kg crude protein, 180-290 g/kg ether extract, 300-550 g/kg neutral detergent fibre (NDF), and 190-244 g/kg acid detergent fibre (ADF) (Brás et al., 2014; Canova et al., 2015; Silva et al., 2015; Goes et al., 2016; Pegoraro et al., 2017; Goes et al., 2018). The crambe cake contains 50.0-77.4 mmol/kg DM of glucosinolates and 56% erucic acid of total fatty acids (Böhme et al., 2005). These characteristics make it possible to use it as a protein source in the feeding of lambs.
Knowledge of the basic nutritional principles of the ingredients and their chemical composition and nutritional value allows adjustment of the amount in the diet, based on nutritional requirements, to maximize performance and avoid loss of production. Among the techniques used to evaluate ruminant diets is apparent digestibility, which evaluates the efficiency of utilization of nutrients in the digestive tract of animals through the balance of nutrients consumed and faecal excretion (Van Soest, 1994).
Ruminal parameters and metabolic profile techniques are commonly used to describe byproducts and identify both potential and efficiency when replacing traditional feed in ruminant diets (Mizubuti et al., 2011). Concentrate levels are high in feedlot systems and may affect ruminal fermentation traits like pH and the production of short chain fatty acids, changing ruminal microbiota activity (Palmonari et al., 2010).
Studies on ruminal microorganisms usually only consider bacteria, but new findings on ciliate protozoa metabolism has associated them w i t h a reduction in methanogenesis, improvements in efficiency, and biotechnology development of biomass degradation processes (Newbold et al, 2015). Rumen protozoa can use all the plant contents through cellulolytic and fermentative pathways. They are a continuous source of nitrogen to bacteria even after their death and total breakdown (Salas et al., 2012; Patel & Ambalam, 2018).
Thus, the goal of this study was to evaluate the intake, digestibility of nutrients, ruminal fermentation, blood parameters and protozoal population in lambs fed crambe cake as a replacement for soybean meal.
Materials and Methods
Two experiments were carried out on the experimental farm of the State University of Londrina (UEL), Brazil, and were approved by the university's Ethics Committee (number 7748.2014.28).
In both experiments, the evaluated diets were formulated to contain increasing levels of crambe cake protein (0, 250, 500, 750, and 1000 g/kg) as a replacement for soybean meal protein in the concentrate ration. All diets were formulated to be isonitrogenous and met the nutrient requirements of growing and finishing lambs (NRC, 2007); a variation of up to 0.5% protein was allowed among them. The roughage:concentrate ratio was 30:70, using maize silage as a roughage (Table 1). To formulate the diets, chemical compositions of ingredients determined at the Laboratory of Animal Nutrition were used, according to the methodologies described by Association of Official Analytical of Chemists (AOAC) (2000, 2006).
The TDN (%) was calculated using the Weiss formula (Weiss, 1993) and used to determine the digestible energy (DE) where:
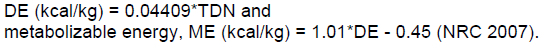
The diets were provided twice a day at 07h30 and 16h30. Mineral supplementation and water were provided ad libitum. The feed supply were adjusted every morning, before the first meal, using the weights of food and leftovers.
In experiment one, 40 crossbred whole male lambs, with an average initial body weight of 20 ± 3.45 kg, were used. The animals were housed in individual stalls with slatted floors, equipped with feeders, mineral supplementation troughs, and drinkers. Animals were allocated in a completely randomized experimental design to five treatments (crambe cake protein at levels 0, 250, 500, 750, and 1000 g/kg), with eight replications per treatment.
The blood samples from all animals were collected at the 15th day for glucose determination by placing 0.5 mL in a portable glucose meter (ACCU CHEK, Roche). Ruminal fermentation parameters (pH and N-NH3 concentration) were evaluated at the 19th day and samples were collected to quantify and identify ciliate protozoa. Ruminal fluid sampling was performed after 12-h fasting (collection period zero) and then at 3, 6, and 9 h after feeding (Zeoula et al., 2003). Approximately 100 mL of ruminal fluid was sampled from each animal using an oesophageal tube.
The pH determination was performed immediately after ruminal fluid sampling with a digital potentiometer calibrated with buffer solutions of pH 4,0 and 7,0. The liquid was filtered with double gauze and placed into hermetically sealed containers within eight drops of sulfuric acid (50% v/v) and frozen at -18 °C for later determination of N-NH3. N-NH3 content was determined by distilling 2 mL of ruminal fluid added to 10 mL 155 KCl solution and 250 mg of magnesium oxide (P.A.) in a micro Kjeldahl distiller and using titration with H2SO4 at 0.01N. The following formulas were used for calculation:
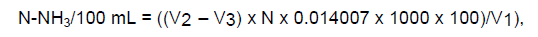
where N-NH3/100mL = ammoniacal nitrogen concentration in 100 mL of ruminal fluid sampled; V1 = volume in mL of ruminal fluid used in the analysis; V2 = sulfuric acid volume used in titration, in mL; V3 = sulfuric acid volume used in reference titration, in mL; N = sulfuric acid normality.
The colour, odour, and consistency of ruminal fluid were observed right after sampling by trained technicians. Colour scale followed a pattern according to animal feeding: olive green, dark brownish green, light yellowish green, and milky/grey green. Abnormal colours of ruminal fluid indicate acidosis (milky/grey) or alkalosis and decomposition (dark green).
The odour was ranked as aromatic, sour acid, and ammoniacal rotting, while consistency was described as aqueous, viscous, and pasty. To measure bacterial activity, 0.5 mL of 0.03%-methylene blue solution was added to 10mL of ruminal fluid without any colouring for the same lamb.
Time was measured until full breakdown of the sample after the addition of colouring, in other words, until it became similar to the control. The following criteria were considered: normal microbiota (3-6 min), simple indigestion (more than 8 min) and acute acidosis (more than 30 min) (Radostits et al., 2002).
In order to quantify and identify the protozoal population, ruminal fluid was filtered with gauze, and 20 mL was fixed with the same volume of 18.5% formalin solution, according to Dehority (1984), adapted by D'Agosto and Carneiro (1999). Then samples were storage in jars in the refrigerator for later analysis. Both identification and quantitative evaluations were performed on a Sedgewick-Rafter counting chamber, according to methodology described by D'Agosto & Carneiro (1999), which uses lugol staining instead of bright green staining. The average count of 100 fields was considered, and the procedure for identification was based on the method of Ogimoto & Imai (1981).
In experiment two, 25 crossbred, entire, male lambs, with an average initial body weight of 23.44 ± 1.04 kg, were used. The animals were housed in individual stalls with slatted floors, equipped with feeders, mineral supplementation troughs, and drinkers. Animals were allocated in a completely randomized experimental design to five treatments (crambe cake protein at levels 0, 250, 500, 750, and 1000 g/kg), with five replications per treatment.
There was an initial period of 10 days for the lambs to adapt to their diets and management routines. The digestibility evaluation lasted 19 d. The first 14 d were for adaptation of animals to the facilities, management routines, and food, followed by 5 d of collecting food (offered and leftovers) and faeces for the determination of the apparent digestibility of nutrients.
Napa collecting bags attached to the animals were used to collect total faeces. Total faecal production was weighed daily and, from the total, an aliquot of approximately 20%/animal/day was taken to provide samples for each animal during the evaluation period.
During the collection period, daily weighing of the food provided, the leftovers, and total faeces was performed, and an aliquot of each was taken to provide representative composite samples for each animal. Later, all samples were analysed in the Laboratory of Animal Nutrition of the State University of Londrina (UEL).
After collection, the samples (diet, leftovers, and faeces) were placed in plastic bags, appropriately identified, and frozen at a temperature of approximately -18 °C. At the end of the experimental period, the samples were thawed and pre-dried in an oven at 55 °C for 72 h. Subsequently, they were ground in a Wiley mill with a 1-mm sieve. All samples were submitted for chemical analyses in order to determine dry matter (DM, method 930,15), mineral matter (MM, method 923,03), crude protein (CP, method 990,03), ether extract (EE, method 920,39), hemicellulose (HEM) according to AOAC (2000); organic matter (OM, method 942,05) following AOAC (2006); and neutral detergent fibre (NDF), acid detergent fibre (ADF), and lignin (LIG) as reported by Detmann et al. (2012). The apparent digestibility coefficient (ADC) of nutrients was determined from the results of nutrients ingested, nutrients in the leftover food, and nutrients excreted in the faeces according to the equations described by Coelho da Silva & Leão (1979). The results of intake, digestibility, and ruminal parameters (protozoal counting was log transformed) were submitted to analysis of variance, and when significant, to regression analysis, setting a 5% level of significance, using the R statistical package (version 3.1.4, 2017). Multivariate analysis using R Studio was performed for the blood glucose analysis.
Results and Discussion
The intake of dry matter (IDM) and total carbohydrates (TC) (kg/lamb/day and g/kg0.75) showed a linear decreasing (P <0.05J with the increase in crambe cake protein as a replacement for soybean meal protein (Table 2). The levels of crambe cake protein affected the intake of ether extract (EE) (kg/lamb/day and g/kg0.75) with a positive linear effect (P <0.05, Table 2). This could be due to the higher EE content of crambe cake when compared to soybean meal (275.54 vs 11.90 g/kg of DM, respectively, Table 1), thus the diets with higher levels of crambe cake had a higher content of EE (Table 1).
The intake of crude protein (kg/lamb/day) showed a linear decrease (P <0.05) with the increase in crambe cake protein to replace the soybean meal protein (Table 2). The diets with 750 and 1000 g/kg DM of crambe cake protein had a lower crude protein content (179.9 and 180.9 g/kg DM, respectively) than the diet without crambe cake (184 g/kg DM). These differences influenced the decrease in the intake of crude protein expressed in kg/lamb/day, but did not affect (P >0.05) the intake of crude protein in g/kg075 (Table 2). Despite the decrease in intake, the protein requirement, as recommended by the NRC (2007), was not affected.
The final body weight decreased linearly (P <0.05) with the increase in crambe cake protein as a replacement for soybean meal protein (Table 2). These results are a consequence of the lower intake of dry matter, total carbohydrates, and crude protein. It was observed that animals feed 1000 g/kg DM of crambe cake protein were 6.49 kg lighter than lambs not fed crambe cake protein (Table 2).
Factors such as energy balance regulate feed intake. This mechanism is regulated by the maintenance and production requirements (Mertens, 1997), and the high content of EE in the diets can be assumed to be the main factor controlling intake. According to Palmquist & Mattos (2006), values above 60.0 g/kg DM in the diet can limit the DM intake and negatively affect the nutrient intake. Another factor is the high content of fatty acids that can increase cholecystokinin hormone release, causing less rumen and reticular motility, as well as reduced intake (Nicholson & Omer, 1983). The level of fatty acids in the blood suppresses the desire for intake in the central nervous system (Medeiros et al., 2015b), which may explain the decreased intake of DM and total carbohydrates. Similar results were obtained by Issakowicz et al. (2017), who evaluated different levels of crambe cake (0%, 22%, 44%, 66%) in replacing dietary soybean meal protein and observed a linear decrease in DM intake (kg/lamb/day).
A lower intake of DM (kg/lamb/day) and high intake of EE (kg/sheep/day) was observed by Brás et al. (2014), who evaluated different co-products from the extraction of oils in sheep diets. However, Mizubuti et al. (2016) observed a higher intake of DM and EE in lambs fed diets containing 70% crambe cake and 30% sorghum silage. Goes et al. (2018) evaluated four levels of crambe cake (0, 50, 100, and 150 g/kg DM) and observed no effect on DM intake but observed an increase in the intake of EE.
The results of the crude protein intake can be due to the lower CP content of crambe cake than soybean meal (280.5 and 455.5 g/kg DM, respectively) (Table 1), as well as the quality of the crambe cake protein. The decrease in final body weight was a consequence of the lower DM intake, total carbohydrates, and crude protein.
The high EE intake resulted in greater absorption of lipids by lambs fed crambe cake. Although lambs fed crambe cake were lighter, these animals may have been modulated in terms of fatty acid profile and fat deposition in the carcass. According to Roh et al. (2016), the brain modulates food intake, energy expenditure, insulin secretion, and glucose / fatty acid metabolism in adipose tissue and skeletal muscle, so when there is an accumulation of fat in the animal, it indicates that the levels of energy metabolites in the animal's blood are high, activating the centre of satiety in the hypothalamus and inhibiting hunger.
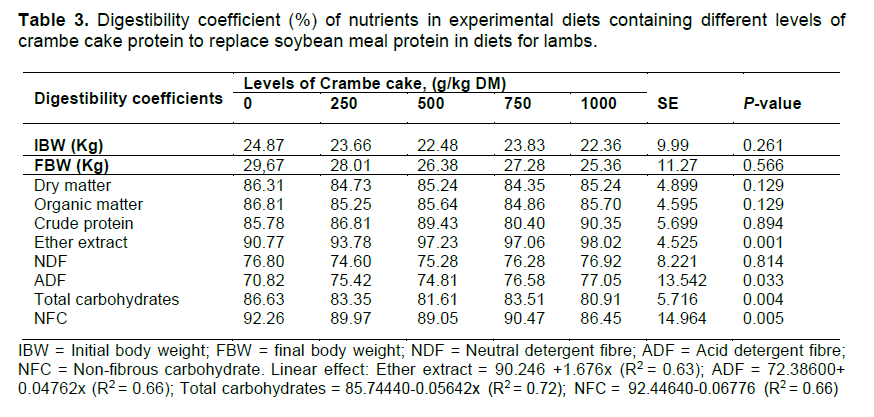
The levels of crambe cake protein influenced the digestibility coefficients (P <0.05) of EE, ADF, total carbohydrates, and NFC (Table 3). The digestibility coefficient of EE and ADF presented a positive linear effect (P <0.05, Table 3). The digestibility coefficient of total carbohydrates and NCF presented a negative linear effect (P <0.05) with a n increase in crambe cake protein as a replacement for soybean meal protein (Table 3).
The results of the digestibility coefficient of total carbohydrates and the NCF may be due to the intake of diets with high EE, which can reduce fibre digestion and the rate of passage of the digest through the gastrointestinal tract. The effect that long-chain fatty acids have on fibre digestion is through coating the food particles, which would make colonization by rumen bacteria difficult (Palmquist & Jenkins, 1980; Maczulak et al., 1981; NRC, 2001).
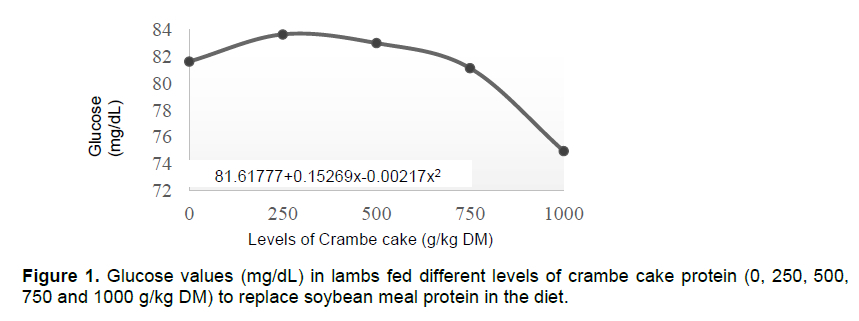
Glucose levels presented a n interaction (P <0.05) within treatments and collection periods. A quadratic effect (P <0.05) was observed as a function of the levels of crambe cake protein, peaking at 351,8 g/kg DM (Figure 1).
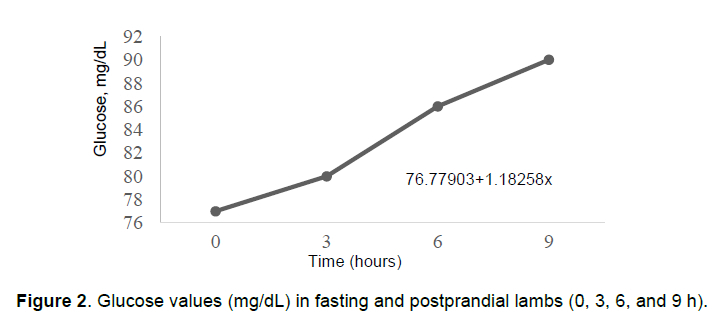
There was positive linear effect (P <0.05) of glucose values over time (0, 3, 6, and 9 h), indicating that after the first feeding, glucose increased and remained high for up to 9 h postprandial, due to its metabolism (Figure 2).
Glucose values in fasting ruminants may vary from 55 to 65 mg/dL (Church, 1993). Although fasting values in this trial were greater, blood composition can range as a function of breed, age, production system, herd management, weather condition, fasting duration, and physiological state of the animal, inter alia (Polizopoulou, 2010). However, postprandial values (3, 6, and 9 h) were considered normal (50-80 mg/dL) (Kaneco et al., 2008). In ruminants, blood glucose is controlled by insulin and glucagon, and levels are kept within a normal range through cortisol (Reece, 2006). Moreover, ruminants differ from monogastrics biochemically, as they are able to use several metabolic pathways for liver gluconeogenesis in order to maintain blood glucose during the postprandial and fasting periods (Kozloski, 2009; Balaro et al., 2012).
Despite homeostatic glucose imbalance, its levels can be altered by glucose precursors, where low consumption of metabolic energy can induce a propionate decrease in the rumen, which is a key factor for glycaemic reduction (Reynolds et al., 2003). However, changing levels and sources of proteins hardly ever affect the plasma glucose concentration (Santos et al., 1998). Increasing levels of crambe cake (0, 22, 44, and 66%) fed to an ovine herd presented blood glucose values ranging from 74.30 to 86.71 mg/dL on the 34th experimental day (Canova et al., 2015). Brás et al. (2014) recorded 66.25 mg/dL after fasting at the last day of their study (18th day).
There was no interaction (P >0.05) within treatments and collection periods (0, 3, 6, and 9 h) in odour, colour, consistency, reductive activity, and ammoniacal nitrogen (NH3). Ruminal fluid presented a n aromatic odour and light yellowish green colour, which was considered normal for feed supplied to the animals. The mean value for ammoniacal nitrogen was 20.66 mg/dL. The optimal ammonia level is ~10 mg/dL (Van Soest, 1994), but it may vary once bacteria are able to both produce and capture ammonia. Therefore, maximum values close to 24 mg/dL are suggested to promote greater substrate breakdown (Mehrez & Orskov, 1977). When evaluating the nutritive value of oilseed crushing byproducts in lamb diets, Brás et al. (2014) found 29.10 mg/dL of ammonia in crambe cake, greater than that stated in literature, but lower than the toxic concentration (100 mg/dL). However, Medeiros et al. (2015a) observed in vitro ammoniacal nitrogen from a myriad of biofuel byproducts replacing maize silage and reported increasing ammonia as replacement levels were higher.
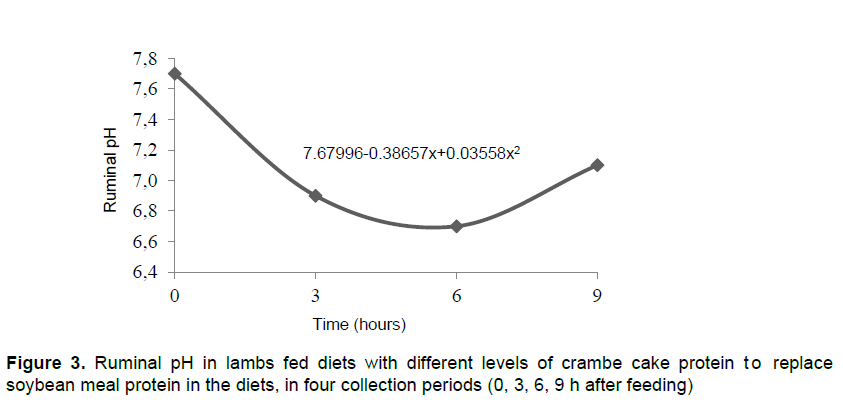
The pH showed a quadratic effect as a function of collection period, reaching a peak at 5.43 h (Figure 3). Rumen pH may vary from 5-8, according to diet and time after feeding (Kozloski, 2009). Before the first meal, pH values might be high due to limited availability of nutrients to ruminal microorganisms and rumination, which stimulates saliva production and promotes a buffering effect on ruminal fluid. After the first meal, the pH is reduced because fermentation reaches its peak 3 to 4 h after feeding (Marino et al., 2011). High-concentrate diets can lead to a rapid pH reduction (Church, 1993). Similar to Van Soest (1994), figures lower than 6.2 suppress intake rate and increase the time to bacterial colonization to the plant cell wall.
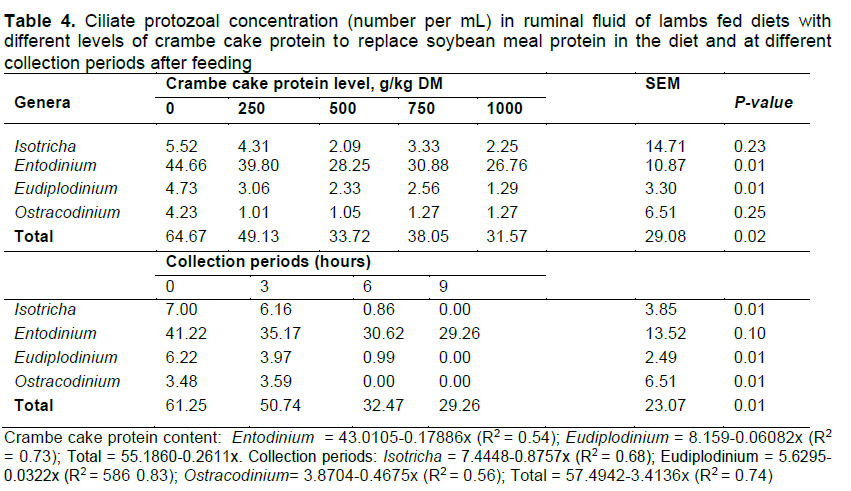
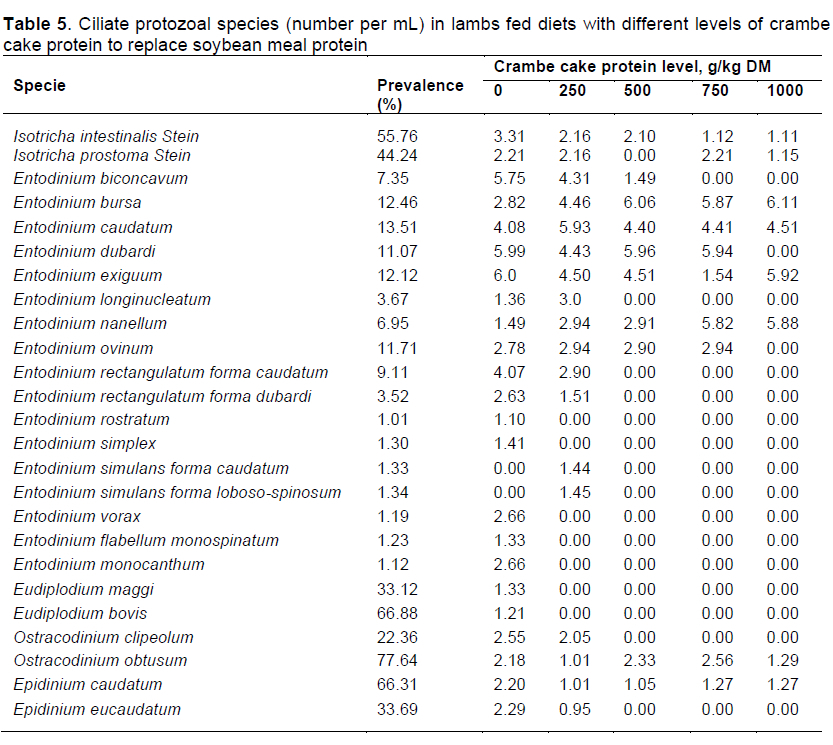
The protozoal population showed a negative linear effect for genera Entodinium, Eudipíodium, and total content as a function of treatment, but there was no effect on genera Isotricha and Ostracodinium within treatments (Table 4). For collection periods, there was negative linear effect of genera Isotricha, Eudipíodium, Ostracodinium, and total count (Table 4). Prevalence is reported in Table 5. Species with greater population were Isotricha intestinaíis Stein (55.76%), Entodinium caudatum (13.51%), Eudipíodium bovis (66.88%), Ostracodiniumobtusum (77.64%), and Epidinium caudatum (66.31%).
There were five genera and 25 species of protozoa (Table 5); animals that were not fed crambe cake protein presented the greatest number of species, however, the protozoal population decreased as levels of crambe cake protein increased in the diets. From genera Entodinium, species E. biconcavum, E. bursa, E. caudatum, E. dubardi, E. exiguum, and E. nanello were observed at all levels of crambe cake supplementation, proving that protozoa have amylolytic activity and a low sensitivity to ruminal pH change and to high concentrate diets (Williams & Coleman, 1992; Dehority, 2003).
Several factors cause changes in the ciliate population, such as digesta retention time, pH, and metabolic characteristics of the host (Michalowski, 1977). As stated by Van Soest (1994), rumination declines with higher concentrate levels, promoting lower saliva excretion, and consequently affecting pH control and buffering activity. Franzolin and Dehority (2010) reported that long periods at low ruminal pH are harmful to ciliate protozoa survival. The protozoal population is driven by several factors, like diet (Booyse et al., 2014) and ruminal pH. Animals receiving diets with more concentrate usually present ruminal pHs ranging from 5.5 to 6.2, and those receiving diets rich in roughage have values between 6.3 and 7.0. A pH reduction may have a negative effect on some protozoal species, such as Eudipíodium maggi et bovis and Epidinium eucaudatum when animals are fed levels above 250 and 500 g/kg DM, respectively (Williams and Coleman, 1992; Dehority, 2003).
The genus, Isotricha, can promote soluble carbohydrate fermentation and storage of amylopectins, while producing acetic, butyric, lactic and, in smaller amounts, propionic acids (Patel & Ambalam, 2018). The enzymatic profile of such genera contains amylase, invertase, pectin esterase, and polygalacturonase in great quantities for starch, pectin, and soluble sugar breakdown as a source of energy, and that seems to be the reason of the unchanged concentration among treatments (Table 4 and 5). There are also enzymes able to break cellulose and hemicellulose, but in smaller amounts (Kamra, 2005).
Isotricha populations depend on diet, intake frequency, and collection period (Dehority and Tirabasso, 1989). In agreement with Williams and Coleman (1992), Isotricha tends to increase as intake frequency is greater, but Entodinium protozoa diminish in the same situation.
In line with Salvio & D'Agosto (1999) and D'Agosto et al. (2001), Isotrichia migrate to the reticulum after fasting, and that is the reason for protozoal absence 9-h post-feeding. Such migration probably occurs as a static chemical response to soluble nutrients associated with strong reticulum contractions during feeding (Dehority & Tirabasso, 1989). Another possible explanation for this specific protozoon disappearance is the high amount of absorbed starch, so they become heavier and tend to reach the bottom (ventral) of the rumen (Franzolin and Franzolin, 2000).
Entodinium prevailed in every treatment due to their wide occurrence in ruminants, and some species are considered rumen colonizers (Ogimoto & Imai, 1981; Kamra, 2005). The Entodinium protozoal population decreases as dietary concentrate increases, similar to cellulolytic protozoa (Franzolin and Dehority, 1996). There was a decrease in Entodinium in this trial as a result of high levels of EE in crambe cake protein to replace soybean meal at 500, 750, and 1000 g/kg DM. Jenkins (1993) stated that EE may have an antimicrobial effect in the rumen by affecting cell membrane biological functions, such that other mechanisms can occur, such as oxidative phosphorylation and Acetyl-CoA transferase inhibition (Faciola et al., 2013).
From Entodinium, species E. biconcavum, E. bursa, E. caudatum, E. dubardi, E. exiguum, and E. nanello were found in all replacement ranges, supporting that protozoa perform amylolitic breakdown, with low sensitivity to changes in ruminal pH and high concentrate diets (Williams & Coleman, 1992; Dehority, 2003).
In a trial to evaluate the prevalence and abundance of ciliate protozoa in Brazilian lambs fed different levels of concentrate (20, 40, 60, and 80%), Cedrola et al. (2016) did not find Isotricha intestinalis in treatments with 20, 40, and 80% and there was a reduction of Entodinium when more concentrate was supplied to animals. These results support the findings of the current study.
The presence of protozoa is crucial for fermentation and to control substrate availability, promoting a constant fermentative process during feeding intervals (Teixeira, 1991; Patel & Ambalam, 2018); the lack of protozoa may lead to intake restriction and inefficiency in feed conversion (Hristov et al., 2013). The reduction of protozoa in treatments with greater ranges of replacement is a consequence of the high EE content in such diets, curbing protozoal capacity of lipidic transformation and absorption, which leads to the rupture of their cells and decreased methanogenesis (Hook et al., 2010). Similar results were observed by Abubakr et al. (2013) when evaluating the digestibility of ruminal protozoa in goats fed palm oil extraction byproducts.
Conclusion
The increasing levels of crambe cake protein in the feeding of lambs as a replacement for soybean meal protein resulted in greater intake and digestibility of EE and ADF, lower intake and digestibility of total carbohydrates, and lambs with a lower final live weight. The crambe cake protein did not impact ruminal and blood parameters in lambs. However, the protozoal population decreased with greater amounts of crambe cake protein and during long periods without feeding. Therefore, it is recommended that soybean meal protein only be replaced with crambe cake protein up to 250 g/kg DM.
Acknowledgements
To Araucária Foundation and Higher Education Personnel Improvement Coordination (CAPES) for funding (A.F./CAPES n°12/2013) this research project and for financing the post-doctoral research grant of AR. Poveda-Parra, and research fellowship of I.Y. Mizubuti; to the Agricultural Engineer, C. Pitol; to the MS Foundation for the Research and Diffusion of Agricultural Technologies, to the Industry and Commerce Matsuda Importer and Exporter Ltd for the financial resources and the donation of the raw materials used in this study.
Authors' Contributions
IYM proposed the project and wrote the manuscript; ESP & OPPC wrote the manuscript; ARPP & ERR executed the experiment, statistically analysed the collected data, and wrote the manuscript; LSC, GLG, KAP, MRL, ERCJ & JPMC worked on the project and laboratory analysis.
Conflict of interest
The authors declare that there is no conflict of interest.
References
Abudakr, A.R, Alimon, A.R, Yaakub, H., Abdullah, N. & Ivan, M., 2013. Digestibility, rumen protozoa, and ruminal fermentation in goats receiving dietary palm oil by-products. J Saudi Soc Agric Sci.12, 147-154. Doi: 10.1016/j.jssas.2012.11.002 [ Links ]
AOAC, 2000. Official Methods of Analysis (17th ed.), Association of Official Analytical Chemists International, Gaithersburg, MD. [ Links ]
AOAC, 2006. Official Methods of Analysis (18th ed.), Association of Official Analytical Chemists International, Gaithersburg, MD. [ Links ]
Balaro, M.F.A, Cardoso, E.C. & Peneiras, A.B.V, 2012. Ganho de peso e perfil metabólico sanguíneo de cordeiros alimentados com dietas contendo gordura protegida. Agroecossistemas, 4(1), 42-49. [ Links ]
Brás, P., Possentti, R.A., Bueno, M.S., Canova, E.B. & Schammas, E.A, 2014. Avaliação nutricional de coprodutos da extração de óleos vegetais em dietas de ovinos. B. Industri. Anim. 71(2),160- 175. doi:10.17523/bia.v71n2p160 [ Links ]
Böhme, H., Kampf, D., Lebzien, P. & Flachowsky, G., 2005. Feeding value of crambe press cake and extracted meal as well as production responses of growing-finishing pigs and dairy cows fed these by-products Arch. Anim. Nutr. 59(2), 111- 122. doi:10.1080/17450390512332387927 [ Links ]
Booyse, D.G., Dehority, B.A. & Reinighaus, B., 2014. Rumen ciliates in the African (Cape) buffalo (Syncerus caffer caffer) living in the vicinity of the Orper Gate entrance into Kruger National Park, South Africa. Zootaxa, 3846(1), 138-144. doi:10.11646/zootaxa.3846.1.8 [ Links ]
Canova, E.B., Bueno, M.S., Moeira, H.L., Possenti, R. & Brás, P., 2015. Crambe cake (Crambe abyssinica hochst) on lamb diets. Cienc.Agrotec. 39(1),75-81. doi:10.1590/S1413-70542015000100009 [ Links ]
Cedrola, F., Martinele, I., Dias, R.J.P. & D'Agosto, M., 2016. Rumen ciliates in Brazilian sheep (Ovis aries) with new records and redescription of Entodinium contractum (Entodiniomorphida: Ophryoscolecidae). Zootaxa, 4088(2), 292-300. doi:10.11646/zootx.4088.2.10 [ Links ]
Church, D.C., 1993. El Rumiante: fisiología digestiva y nutrición. Acribia: Zaragoza Spain. [ Links ]
Coelho Da Silva, J.F. & Leão, M.I., 1979. Fundamentos de nutrição dos ruminantes. Piracicaba: Ed. Livroceres. [ Links ]
D'Agosto, M., Siqueira, I.C.V. & Espírito-Santo, N.B., 2001. Comportamento e distribuição de protozoários ciliados (Protista, Ciliophora) no rúmen e no retículo de bovinos submetidos ao jejum. Rev Bras Ciênc Vet. 8(1),16-18. doi: 10.4322/rbcv.2015.326 [ Links ]
D'Agosto, M. & Carneiro, M.E., 1999. Evaluation of lugol solution used for counting rumen ciliates. Rev Bras Zool. 16(3), 725-729. doi:10.1590/S0101-81751999000300011 [ Links ]
Dehority, B.A., 1984. Evaluation of subsampling and fixation procedures used for counting rumen protozoa. Appl Environ Microbiol. 48(1),182-185. doi:10.1128/aem.48.1182-185.1984 [ Links ]
Dehority, B.A., 2003. Rumen Microbiology. Nottingham University Press. Nottingham, UK. [ Links ]
Dehority, B.A. &Tirabasso, P.A., 1989. Factors affecting the migration and sequestration of rumen protozoa in the family Isotrichidae. J Gen Microbiol. 135, 539-548. doi:10.1099/00221287-135-3-539 [ Links ]
Detmann, E., Souza, M.A., Valadares Filho, S.C., Queiroz, A.C., Berchiolli, T.T., Saliba, E.O.S., Cabral, L.S., Pina, D.S., Ladeira, M.M. & Azevedo, J.A.G., 2012. Métodos para análise de alimentos. Visconde de Rio Branco. MG: UFV. [ Links ]
Faciola, A.P., Broderick, G.A., Hristov, A. & Leão, M.I., 2013. Effects of lauric acid on ruminal protozoal numbers and fermentation pattern and milk production in lactating dairy cows. J Anim Sci. 91,363-373. doi:10.2527/jas.2012-5168 [ Links ]
Franzolin, R. & Dehority, B.A., 1996. Effect of prolonged high-concentrate feeding on ruminal protozoa concentrations. J. Anim Sci. 74, 2803-2809. doi:10.2527/1996.74112803x [ Links ]
Franzolin, R. & Dehority, B.A., 2010. The role of pH on the survival of rumen protozoa in steers. Rev Bras Zootec. 39(10), 2262-2267. doi:10.1590/S1516-35982010001000023 [ Links ]
Franzolin, R. & Franzolin, M.H.T., 2000. População protozoários ciliados e degradabilidade ruminal em búfalos e bovinos zebuínos sob dieta à base de cana-de-açúcar. Rev Bras Zootec. 29(6),1853-1861. doi:10.1590/S1516-35982000000600034 [ Links ]
Goes, R.H.T.B., Souza, K.A., Patussi, R.A., Cornelio, T.C., Oliveira, E.R. & Brabes, K.C.S., 2010. Degradabilidade in situ dos grãos de crambe, girassol e soja e de seus coprodutos em ovinos. Acta Sci., Anim. Sci. 32(3), 271-277. doi:10.4025/actascianimsci.v32i3.7913 [ Links ]
Goes, R.H.T.B., Carneiro, M.M.Y., Brabes, K.C.S. & Lana, R.P., 2016. Coprodutos de crambe (Crambe abyssinica Hoeschst) na alimentação de ruminantes. Arch Zootec. 65(R), 7-16. doi:10.21071/az.v65i249.450 [ Links ]
Goes, R.H.T.B., Carneiro, M.M.Y., Osmari, M.P, Souza, K.A., Oliveira, R.T. & Souza, CJS, 2018. Intake, digestibility, performance, and carcass characteristics of ewes fed crambe replacing soybean meal in the diet. Acta Sci., Anim. Sci. 40, e37171. doi:10.4025/actascianimsciv40i1.37171 [ Links ]
Hook, S.E., Wright, A.D.G. & McBride, B.W., 2010. Methanogens: Methane producers of the rumen and mitigation strategies. Archaea. 100. doi:10.1155/2010/945785 [ Links ]
Hristov, N.A., Oh, J., Firkins, J.L., Dijkstra, J., Kebreab, E., Waghorn, G., Makkar, H.P.S., Adesogan, A.T., Yang, W., Lee, C., Gerber, P.J., Henderson, B. & Tricarico, J.M., 2013. Mitigation of methane and nitrous oxide emissions from animal operations: A review of enteric methane mitigations options. J Anim Sci. 11(11), 5045-5069. doi:10.2527/jas.2013-6583. [ Links ]
Issakowicz, J., Bueno, M.S., Barbosa, C.M.P., Canova, E.B., Moreira, H.L., Geraldo, A.T. & Sampaio, A.C.K., 2017. Crambe cake impairs lamb performance and fatty acid profile of meat. Anim. Prod. Sci. 57, 785-792. doi:10.1071/AN1536 [ Links ]
Jenkins, T.C., 1993. Lipid-metabolism in the rumen. J Dairy Sci. 76:3851-3863. Doi:10.3168/jds.S0022-0302(93)77727-9 [ Links ]
Kamra, D.N., 2005. Rumen microbial ecosystem. Curr Sci. 89, 124-134. http://www.jstor.org/stable/24110438 [ Links ]
Kaneko, J.J., Harvey, J.W. & Bruss, M.L., 2008. Clinical Biochemistry of Domestic Animals. Academic Express, Califórnia, USA. [ Links ]
Kozloski, G.V., 2009. Bioquímica dos Ruminantes. Ed. UFSM, Santa Maria, Brazil. [ Links ]
Maczulak, A.E., Dehority, B.A. & Palmquist, D.L., 1981. Effects of long-chain fatty acids on growth of rumen bacteria. Appl. Environ. Microbiol. 42, 856-862. doi:10.1128/aem.42.5.856-862.1981 [ Links ]
Marino, C.T., Otero, W.G., Rodrigues, P.H.M., Dicostanzo, A., Millen, D.D., Pacheco, R.L.D., Dilorenzo, N., Martins, C.L. & Arrigoni, M.D.B., 2011. Effects of adding polyclonal antibody preparations on ruminal fermentation patterns and digestibility of cows fed different energy sources. J. Anim Sci. 89, 3228-3235. doi:10.2527/jas.2010-3062 [ Links ]
Medeiros, F.F., Bezerra, L.R., Silva, A.M.A., Carneiro, H., Morais, R.K.O., Moreira, M.N. & Pereira Filho, J.M., 2015a. Greenhouse gases, short-chain fatty acids and ruminal pH in vitro of biodiesel byproducts to replace corn silage. Rev Bras Saúde Prod Anim. 16 (4), 935-947. doi:10.1590/S1519-99402015000400017 [ Links ]
Medeiros, S.R., Gomes, R.C. & Bungenstab, D.J., 2015b. Nutrição de bovinos de corte: Fundamentos e aplicações. Brasília, DF: Embrapa, 176 p. [ Links ]
Mehrez, A.Z. & Orskov, E.R., 1977. A study of the artificial fiber bag technique for determining the digestibility of feed in the rumen. J Agric Sci. 88, 645-665.460 doi:10.1017/S0021859600037321. [ Links ]
Mendonça, B.P.C., Lana, R.P., Detmann, E., Goes, R.H.T.B. & Castro, T.R., 2015. Torta de crambe na terminação de bovinos de corte em confinamento. Arq. Bras. Med. Vet. Zootec. 67(2),583-590. doi:10.1590/1678-6753 [ Links ]
Mertens, D.R., 1997. Creating a system for meeting the fiber requirements of dairy cows. J. Dairy Sci. 80, 1463-1481. doi:10.3168/jds. S0022-0302(97)76075-2 [ Links ]
Michalowski, T., 1977. Diurnal changes in concentration or rumen ciliates and in occurrence of dividing forms in water buffalo (Bubaíus bubaíus) fed once daily. Appl Environ Microbiol. 33(4), 802-804. doi:10.1128 /aem.33.4.802-804.1977 [ Links ]
Mizubuti, I.Y., Ribeiro, E.L.A., Pereira, E.S., Pinto, A.P., Franco, A.L.C., Syperreck, M.A., Dórea, J.R.R., Cunha, G.E., Capelari, M.G.M. & Muniz, E.B., 2011. Cinética de fermentação ruminal in vitro de alguns co-produtos gerados na cadeia produtiva do biodiesel pela técnica de produção de gás. Semin Cienc Agrar. 32 (suppl.1), 2021-2028. doi:10.1590/1678-4162-8618 [ Links ]
Mizubuti, I.Y., Syperreck, M.A., Ribeiro, E.L.A., Pereira, E.S., Pinto, A.P., Prado, O.P.P., Peixoto, E.L.T., Parra, A.R.P., Massaro Junior, F.L. & Guerra, G.L., 2016. Consumo, digestibilidade e balanço de nitrogênio em ovinos alimentados com rações contendo torta de Crambe. Arq. Bras. 476 Med. Vet. Zootec. 68(3),761-768. doi:10.1590/1678-4162-8618 [ Links ]
Newbold, C.J., Fuente, G., Belanche, A., Ramos-Morales, E. & McEwan, N.R., 2015. The role of ciliate protozoa in the rumen. Front Microbiol. 6, 1-14. Doi:10.3389/fmicb.2015.01313 [ Links ]
Nicholson, T. & Omer, S.A., 1983. The inhibitory effect of intestinal infusions of unsaturated long-chain fatty acids on forestomach motility of sheep. Br. J. Nutr. 50, 141-149. doi:10.1079/bjn19830081. [ Links ]
NRC, 2000. Nutrient Requirements of Beef Cattíe. Seventh Revised Edition. Washington, DC: The National Academies Press. http://doi.org/10.17226/9761. [ Links ]
NRC, 2001. Nutrient Requirements of Dairy Cattíe. Seventh Revised Edition. Washington, DC: The National Academies Press. http://doi.org/10.17226/9825. [ Links ]
NRC, 2007. Nutrient Re quirements of S maíí R uminants: S heep, goats, cervids, and new world camelids. Washington, DC: The National Academies Press. http://doi.org/10.17226/11654. [ Links ]
Ogimoto, K. & Imai, S., 1981. Atlas of Rumen Microbioíogy. Japan Scientific Societies Press, Tokyo, Japan. [ Links ]
Palmonari, A., Stevenson, D.M., Mertens, D.R., Cruywagen, C.W. & Weimer, P.J. 2010. pH dynamics and bacterial community composition in the rumen of lactating dairy cows. J Dairy Sci. 93(1), 279-287. doi:10.3168/jds.2009-2207 [ Links ]
Palmquist, D.L. & Jenkins, T.C., 1980. Fat in lactation rations: Review. J. Dairy Sci. 63, 1-14. doi:10.3168/jds.S0022-0302(80)82881-5 [ Links ]
Palmquist, D.L. & Mattos, W.R.S., 2006. Metabolismo de lipídeos. In: Nutrição de Ruminantes. Ed. Berchielli TT, Pires AV, Oliveira SG. 615p. Jaboticabal: FUNEP. [ Links ]
Patel, S. & Ambalam, P., 2018. Role of rumen protozoa: Metabolic and fibrolytic. Adv Biotech & Micro. 10(4), 79-84. doi:10.19080/AIBM.2018/10.555793 [ Links ]
Pegoraro, M., Silva, L.D.F.,Fernandes Junior, F., Massaro Junior, F.L., Fortaleza, A.P.S., Grandis, F. A., Ribeiro, E.L.A., Castro, F.A.B. 2017. Avaliação nutricional e cinética de degradação in vitro de concentrados proteicos utilizados na alimentação de ruminantes. R. Bras. Ci. Vet., 24(1), 31-38. doi: 10.4322/rbcv.2017.007 [ Links ]
Polizopoulou, Z.S., 2010. Haematological tests in sheep health management. Small Rumin Res.92, 88-91. doi:10.1016/j.smallrumres.2010.04.015 [ Links ]
Radostits, O.M., Mayhew, I.G.J. & Houston, D.M., 2002. Exame clínico e diagnóstico em veterinária. Guanabara Koogan Guanabara Koogan, Rio de Janeiro, Brazil. [ Links ]
Ramos, L.P., Kothe, V., César-Oliveira, M.A.F., Muniz-Wypych, A.S., Nakagaki, S., Krieger, N., Wypych, F. & Cordeiro, C.S., 2017. Biodiesel: Matérias-primas, tecnologias de produção e propriedades combustíveis. Rev Virtual Quim. 9 (1), 317-369.doi:10.21577/1984-6835.20170020 [ Links ]
Reece, W.O., 2006. Fisioíogia dos Animais Domésticos. Guanabara Koogan Guanabara Koogan, Rio de Janeiro, Brazil. [ Links ]
Reynolds, C.K., Aikman, P.C., Lupoli, B., Humphries, D.J. & Beever, D.E., 2003. Splanchnic metabolism of dairy cows during the transition from late gestation through early lactation. J Dairy Sci. 86, 1201-1217. doi:10.3168/jds. S0022-0302(03)73704-7 [ Links ]
Roh, E., Song, D.E.K. & Kim, M.S., 2016. Emerging role of the brain in the homeostatic regulation of energy and glucose metabolism. Exp. Mol. Med. 48, e216. doi:10.1038/emm.2016.4 [ Links ]
Salas, R.Z., Builes, A.G. & Echeverry, D.P., 2012. Role of rumen ciliated protozoa in the synthesis of conjugated linoleic acid: A review. Rev Colom Cienc Pecua. 25(1), 135-149. [ Links ]
Sálvio, G.M.M. & D'Agosto, M., 1999. Avaliação do comportamento de escape e migração de ciliados Isotriquidae no sistema rúmen-reticulo de bovinos. Rev Bras Zoociências, 1 (1,2), 41-53. [ Links ]
Santos, F.A.P., Santos, J.E.P., Theurer, C.B. & Huber, J.T., 1998. Effects of rumen undegradable protein on dairy cow performance: A 12-year literature review. J Dairy Sci. 81 (12), 3182-3213. doi:10.3168/jds.S0022-0302(98)75884-9 [ Links ]
Silva, A.L., Marcondes, M.I., Veloso, C.M., Sousa, F.C. Knupp, L.S., 2015. Simulation of rumen fermentation kinetics of by-products from the biodiesel industry with in vitro gas production technique. Semin Cienc Agra 36 (6), 3851-3862. doi: 10.5433/1679-0359.2015v36n6p3851 [ Links ]
Souza, A.D.V., Fávaro, S.P., Ítavo, L.C.V. & Roscoe, R., (2009). Caracterização química de sementes e tortas de pinhão-manso, nabo-forrageiro e crambe. Pesqui. Agropecu. Bras. 44(10),1328-530. doi:10.1590/S0100-204X2009001000017. [ Links ]
Teixeira, J.C., 1991. Nutrição dos Ruminante, Lavras, MG: ESAL/ FAEPE. [ Links ]
Van Soest, P.J., 1994. Nutritional Ecology of the Ruminant. (2nd edn). Ithaca, Cornell University Press. [ Links ]
Weiss, W.P., 1993. Predicting energy values of feeds. In: Symposium: Prevailing concepts in energy utilization by ruminants. J. Dairy Sci., 76:1802-1811. doi:10.3168/jds.S0022-0302(93)77512-8 [ Links ]
Williams, A.G. & Coleman, G.S., 1992. The Rumen Protozoa. Springer-Verlag, New York Inc., London, UK. [ Links ]
Yang, K., Quing, Y., Yu, Q., Tang, X., Chen, G., Fangt, R. & Liu, H., 2021. By-product feeds: Current understanding and future perspectives. Agriculture,11207. doi:10.3390/agriculture11030207 [ Links ]
Zeoula, L.M., Caldas Neto, S.F., Geron, J.V., Maeda, E.M., Prado, I.N., Dian, P.H.M., Jorge, J.R.V. & Marques, J.J.A., 2003. Substituição do milho pela farinha de varredura de mandioca (Manihot esculentacrantz) em rações de ovinos: Consumo, digestibilidade, balanços de nitrogênio e energia e parâmetros ruminais. Rev. Bras. Zootec., 32 (2), 491-502. doi:10.1590/S1516-35982003000200030 [ Links ]
Submitted 1 September 2022
Accepted 19 October 2022
Published 6 March 2023
# Corresponding Author: angelpov@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}