










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 no.6 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i6.09
Responses of broiler chickens fed diets containing sunflower meal and wheat middlings pre-treated with enzymes
F. MuchiriI, II; C. K. GachuiriI; E. G. KiarieII, #
IDepartment of Animal Production, University of Nairobi, Kenya Postal address: 30197, 00100, GPO, Nairobi, Kenya
IIDepartment of Animal Biosciences, University of Guelph, ON, Canada Postal address: 50 Stone Road E, N1G 2W1 ON, CA
ABSTRACT
The effects of feeding diets containing sunflower and wheat middlings pre-treated with fibre-degrading enzymes were investigated. Based on hatch body weight (BW), 288 Ross-708 male broiler chicks were placed in cages (six birds/cage). Diets were: 1) positive control (PC), a maize-soybean positive control; 2) negative control (NC), PC plus untreated sunflower meal (USM) and wheat middlings (UWM); and 3) four test diets in which USM and UWM were replaced with pre-treated sunflower (TSM) and wheat middlings (TWM) at 25% (NC25), 50% (NC50), 75% (NC75) and 100% (NC100). The pre-treated feedstuffs were mixed with 1 % of FDE in a ratio of 1:2 w/w for feedstuff:water and incubated for 24 hours at 40 °C, and oven-dried before feed preparation. Diets were formulated for two phases (starter, 0-21 d) and finisher (22-42 d). Diets were allocated in a completely randomized design (eight replicates per diet) and birds had free access to feed and water. Bodyweight (BW) and feed intake (FI) were monitored by phase, and one bird per cage was sacrificed on days 21 and 42 for samples. The BW and BW gain (BWG) of NC up to NC50 were not different relative to PC, but NC75 and NC100 were lower than PC throughout the study. On day 21, liver weight had increased linearly and on day 42, tibia length and diameter linearly decreased with the inclusion of TSM and TWM. In conclusion, TSM and TWM at low levels did not affect performance while high inclusion levels reduced the performance of the broilers.
Keywords: cellulase, fibre-degrading enzymes, mannanase, pectinase, xylanase
Introduction
Agricultural by-products, such as those from sunflower seed meal and wheat processing, are abundant. They have been explored as the alternative source of protein and energy to replace a portion of soybean meal and maize, respectively (Alshelmani et al., 2021). The main challenges with using non-conventional feedstuffs are the anti-nutritional factors, such as non-starch polysaccharides (NSPs) (Mbukwane et al., 2022). The inclusion of ingredients that are high in dietary fibre in broiler chicken diets has been associated with gut health and growth performance problems (Hamedi et al., 2012). Feed enzymes such as phytase, carbohydrase, and proteases have been widely researched and used as additives to degrade anti-nutritional factors and other substances that are not digested by endogenous enzymes (Adeola & Cowieson, 2011; Godoy et al., 2018; Kiarie et al., 2018). Raza et al. (2009) supplemented broiler chicken diets containing 10% sunflower meal and 6% crude fibre with multienzymes, and broiler performance was improved relative to a negative control. In another study where grower and finisher broilers were fed diets with sunflower meal inclusion and supplemented with enzymes (xylanase, cellulase, and β-glucanase), the growers were more responsive than the finishers (Horvatovic et al., 2015). Wheat middlings can be included up to 30% in both broiler and layer chicken diets without affecting their performance (Ahmadi & Amini, 2014; Abudabos, 2016). However, these enzymes encounter physiological limitations inside the poultry gut, thus reducing their efficacy (Ravindran, 2013). To maximize enzyme efficacy, researchers are now focusing on pre-treating these fibrous materials outside the bird. For example, Pedersen et al. (2015) used xylanases with or without proteases to pre-treat maize dried distiller's grains with solubles (DDGS) and wheat DDGS, and they showed that the enzymes were able to increase the solubilization of crude protein and mono-sugars. Sathitkowitchai et al. (2018) also pre-treated palm kernel cake with 500 units of β-mannanase and showed an increase in protein solubilization and mono-sugars.
Most of the available data has focused on the inclusion of a single, untreated, high dietary fibre ingredient in broiler chicken diets, but with the increasing shortage of conventional ingredients, poultry nutritionists, especially in developing countries, will find themselves being forced to include more than one agricultural by-product in feed formulations. Most of the enzymatic pre-treatment studies have only been conducted in vitro. The novelty of the current study was that two ingredients high in dietary fibre were enzymatically pre-treated and used to replace untreated ingredients in practical broiler chicken diets. To investigate the effects of pre-treatment, a negative control (NC) diet with higher neutral and acid detergent fibre when compared with the positive control (PC), and four test diets with different inclusion levels of pre-treated ingredients, were formulated. It was hypothesized that pre-treatment would restore the performance of the negative control to that or better than the PC.
Materials and Methods
The guidelines established by the Canadian Code of Practice for the Care and Use of Animals for Scientific Purposes were followed (CCAC, 2009). The University of Guelph Animal Ethics Committee research approved the broiler study, and animal trials were conducted at the University of Guelph, Arkell Poultry Research Station (license number 4403).
Wheat middlings (UWM) were obtained from Floradale Feed Mill Limited (Floradale, ON, Canada) and sunflower meal (USM) from Persall Fine Foods Co. (Waterford, ON, Canada). Both products were used without any further processing. Wheat middlings (UWM) and sunflower meal (USM) were pre-treated separately by mixing with enzyme cocktails (at the rate of 1% w/w) made of xylanase/cellulase/ß-glucanase for UWM and cellulase/ß-glucanase/ß-mannanase and pectinase for USM. This was then mixed with tap water in a ratio of 1:2 (Water: UWM/USM). The enzyme inclusion rate was based on the manufacturer's recommendations. The wet mixture was then incubated for 24 h at 40 °C in an incubator shaker (Controlled Environment Incubator Shaker, New Brunswick Scientific, Enfield, CT) without shaking, but hand-mixed using a wooden spatula three times within 24 h. The pre-treated product (treated wheat middlings (TWM) and treated sunflower meal (TSM)) was then oven-dried at 65 °C for three days. This procedure was repeated until enough material was obtained for experimental diet formulation. The enzyme target activity was 60,000 u/g xylanase, 8,000 u/g glucanase, 38,000 u/g cellulase for wheat middlings; and 40,000 u/g glucanase, 45,000 u/g cellulase, 12,000 u/g ß-mannanase and 25,000 u/g pectinase for sunflower meal. All the enzymes were obtained from Canadian BioSystems, Calgary, Canada (Rho et al., 2020). These pre-treatment conditions were optimized and previously validated (Rho et al., 2020).
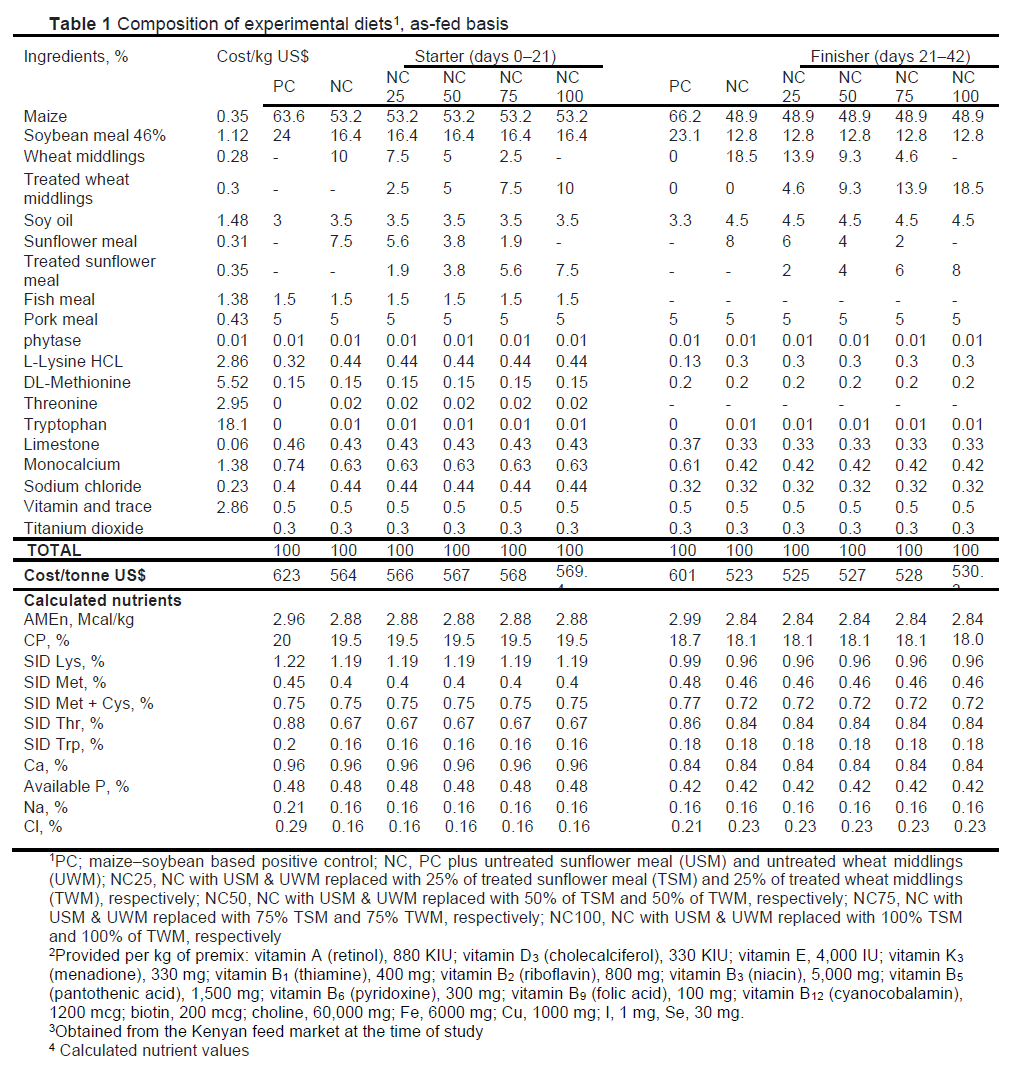
Six dietary treatments were formulated to meet or exceed the NRC nutrient requirements for broiler chickens (NRC, 1994). Birds were fed in two phases: starter (0-21 d) and finisher phase (21-42 d). The ingredient specifications were obtained from the INRA feed tables (INRA CIRAD AFZ, 2022). The diets were: Treatment 1) PC, a maize-soybean positive control; Treatment 2) NC (negative control), PC plus inclusion of untreated sunflower meal (USM) and wheat middling (UWM); and Treatments 36) four test diets in which USM and UWM were replaced with pre-treated sunflower (TSM) and wheat middlings (TWM) at 25% (NC25), 50% (NC50), 75% (NC75), and 100% (N100) (Table 1). The NC and TD were formulated to have a low nutrient density with a lower AMEn. The metabolisable energy (ME) of the negative control (NC) and test diets was reduced by 80 kcal/kg and 150 kcal/kg in the starter and finisher phases, respectively, against the PC. This reduction was done by hypothesizing that pre-treatment would unlock fibre-bound nutrients in SM and WM, which would enable the birds to utilize them, thus restoring the performance of test diets to that of the PC. The NC was expected to perform poorly. All diets were formulated based on equal, total, standardized ileal digestible amino acids (AA). To meet phosphorous requirements, all diets were supplemented with phytase (750 FTU/kg) obtained from CBS Bio Platform Inc. (Alberta, Calgary, Canada).
A total number of 288 Ross-708-day-old male broiler chicks were purchased from a local hatchery (Maple Leaf Foods, New Hamburg, ON, Canada), weighed, and placed in cages (six birds per cage). The trial was conducted in two phases: starter (0-21 d) and finisher phases (21-42 d). The cages were housed in a temperature and light-regulated room. The temperature was maintained at 32 °C for the first 7 d and then gradually reduced by 3 °C on a weekly basis to 24 °C for the first 3 w, then maintained at this temperature until the end of the experiment, while the lighting was maintained at 17 h light for the entire trial period. The six diets were allocated to cages in a completely randomized design to give eight replicates per diet. Birds had free access to feed and water throughout the trial.
Feed intake and body weight of the chicken were determined on days 21 and 42 for calculating body weight gain (BWG) and feed conversion ratio (FCR). Mortality was recorded as it occurred and used to calculate the corrected-FCR. One bird per cage was randomly selected and sacrificed by cervical dislocation on days 21 and 42 for organ weight and tibia sample collection.
Before analyses, the samples (ingredients and mixed feed) were ground using a coffee grinder (CBG5 Smart Grind, Applica Consumer Products, Inc., Shelton, CT). Gross energy (GE), AA, and crude fat were determined at the University of Guelph, whereas dry matter (DM), starch, crude protein, neutral detergent protein, soluble crude protein, neutral detergent fibre, acid detergent fibre, ash, calcium, and phosphorous were determined in a commercial lab (SGS Canada Inc., Agricultural Services Agri-Food Laboratories, Guelph, ON, Canada). An adiabatic bomb calorimeter was used to determine gross energy (IKA Calorimeter System C 6000; IKA Works, Wilmington, NC). The dietary particle size was analysed using US sieves and a pan stuck together in ascending order of numbers 6, 8, 12, 16, 20, 30, 40, 50, 70, 100, 140, 200, 270.
The cage was the experimental unit. The data was analysed using the PROC GLIMIXX of SAS v9.4 with diet as the fixed factor in the model. The linear statistical model used was:

where: Yij = observations recorded such BWG, BW, FI, FCR; μ = overall population mean; α = treatment effect; ɛij = random error effect associated with observations recorded.
Least Square Means were separated using Tukey's test, while pre-planned orthogonal and polynomial contrast statements were used to compare the performance of NC against PC (PC vs NC), and the response of diets containing the test feedstuffs. The level of statistical significance was pre-set at P <0.05.
Results and Discussion
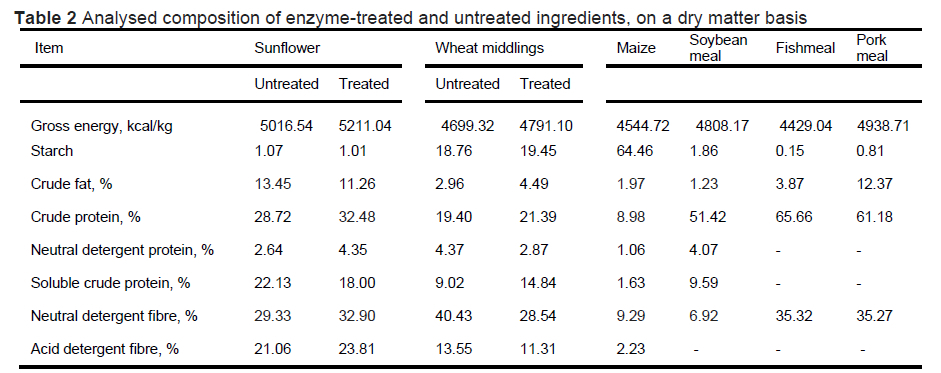
The ingredient analysis results are presented in Table 2. Relative to untreated sunflower meal, pre-treated sunflower meal (TSM) had more gross energy, less starch, less crude fat, more crude protein, more neutral detergent protein, less soluble crude protein, more neutral detergent fibre (NDF) and more acid detergent fibre (ADF). It was expected that enzymatic pre-treatment of sunflower meal would increase soluble protein and reduce neutral detergent protein and NDF relative to untreated sunflower meal, but the opposite happened. Although the current study did not analyse for Maillard reaction products, when ingredients rich in free amino acids (especially lysine and carbohydrates) are thermally processed, they react with mono-sugars to form complex products known as Maillard reaction products (Teodorowicz et al., 2018). The solubilised crude protein in the current study might have been involved in Maillard reactions. When Almeida et al. (2014) autoclaved sunflower meal at 130 °C, the analysed neutral detergent fibre (NDF) and acid detergent fibre (ADF) increased. The Maillard reaction products are captured as part of the NDF during laboratory analysis (Oliveira et al.,2021). This explains the observed increase in NDF in pre-treated sunflower meal (TSM) in the current study. When Knipfel et al. (1983) heated alfalfa, heating led to an increase in NDF and nitrogen content in NDF and ADF. In the current study, relative to untreated wheat middlings, pre-treated wheat middlings (TWM), had more gross energy, starch, crude fat, and crude protein, less neutral detergent protein, more soluble crude protein, less NDF, and less ADF. These results indicate that enzymatic pre-treatment of wheat middlings was effective in solubilizing trapped crude protein and hydrolysing non-starch polysaccharides. The analysis of both pre-treated sunflower meal (TSM) and pre-treated wheat middlings (TWM) after freeze-drying indicated an increase in crude protein, soluble protein, and a reduced NDF relative to untreated SM and WM (data not shown). It can therefore be hypothesized that it is oven-drying that affects the nutritional composition of TSM negatively.
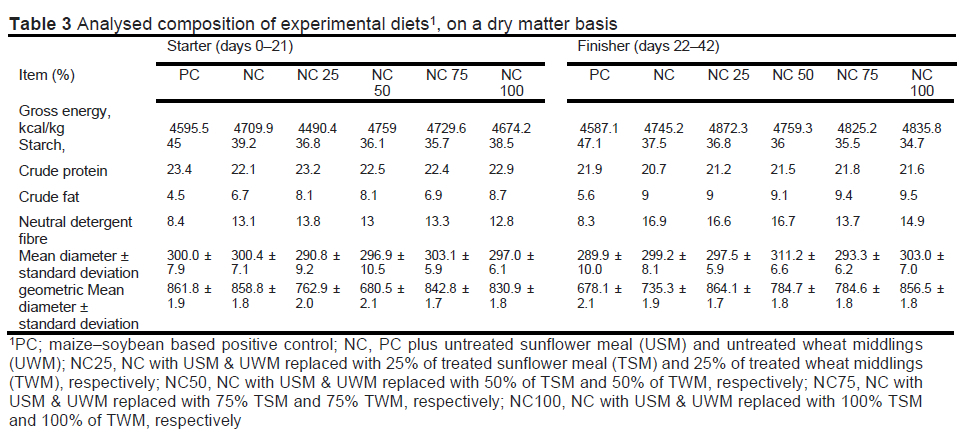
Analysis of the treatment diets is in Table 3. Compared with the PC, negative control diets had more gross energy, less starch, less crude protein, more crude fat, and more NDF in both the starter and finisher diets. The mean diameter ± standard deviation for PC, NC, N25, N50, N75, and N100 starter diet particle size was 300 ± 7.91, 300.43 ± 7.09, 290.78 ± 9.23, 296.91 ± 10.47, 303.08 ± 5.9, and 296.95 ± 6.11 μm, respectively; their geometric mean diameter (GMD) ± standard deviation was 861.83 ± 1.90, 858.79 ± 1.83, 762.85 ± 2.01, 680.54 ± 2.07, 842.79 ± 1.73, and 830.91 ± 1.76 μm, respectively. The mean diameter ± standard deviation for PC, NC, N25, N50, N75, and N100 finisher diet particle size was 289.93 ± 10.02, 299.2 ± 8.1, 297.47 ± 5.89, 311.21 ± 6.57, 293.26 ± 6.21, and 302.95 ± 6.98, respectively; their geometric mean diameter ± standard deviation was 678.14 ± 2.05, 735.25 ± 1.90, 864.11 ± 1.74, 784.69 ± 1.77, 784.57 ± 1.77, and 856.51 ± 1.82, respectively. According to Nir et al. (1994), young broiler chickens fed diets in mash form require feed with a GMD of 700-900 μm for optimal performance. All diets in the current study were in this range.
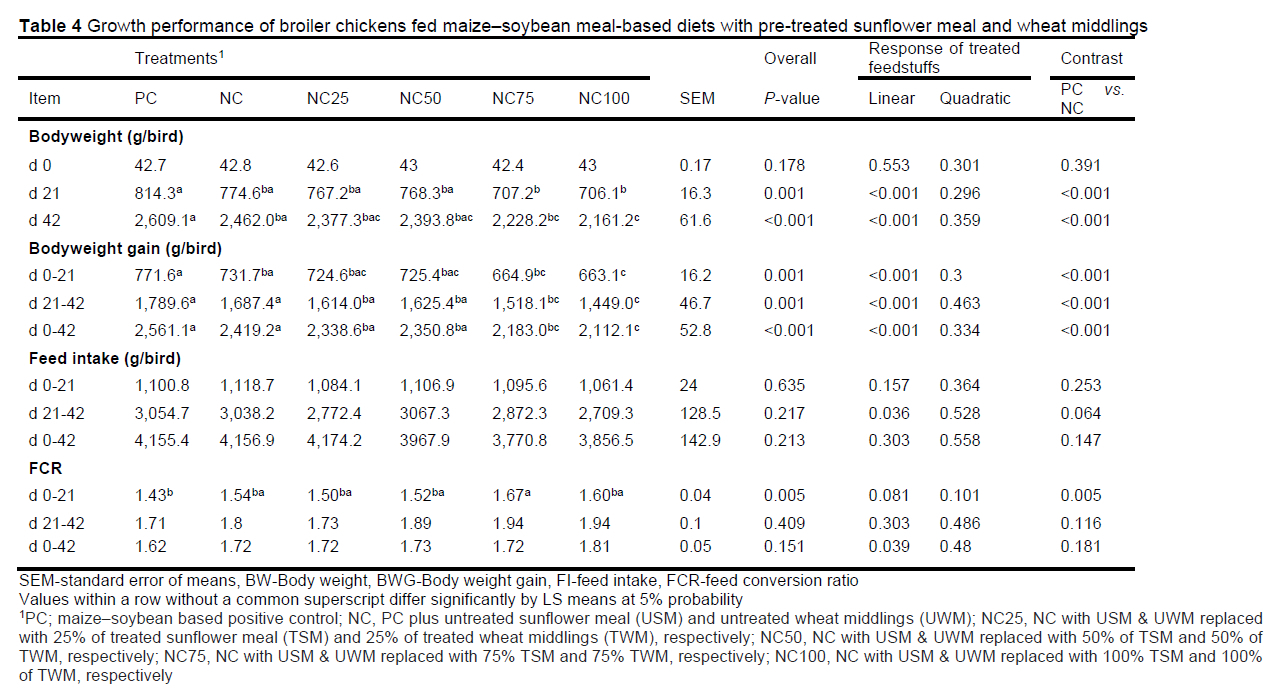
Relative to PC, the NC orthogonal contrast BW and BWG was lesser throughout the study (P <0.001), but FI was not different (P >0.064) (Table 4). The reason as to why the BW and BWG were different despite their FI being not different is due to high crude fibre in NC, and the birds were unable to utilize the extra crude fibre. The NC had 4.39% more NDF than PC (Table 3). The current study was designed such that the NC and test diets were deficient in energy (80 kcal/kg in starter and 150 kcal/kg in finisher diets) relative to the respective PC diets (Table 1) to determine whether birds would increase their feed intake to compensate; however, there seems to be a physiological limitation in intake capacity. This agrees with Tejeda & Kim (2020), who observed similar FI between the control and a high fibre diet (8% soy hulls), despite the two diets being isonitrogenous and isocaloric. Diets with soluble fibre increase gut viscosity and reduce feed movement rate and performance thus reducing FI (Kiarie et al., 2014; Perera et al., 2019). The NC feed conversion ratio (FCR) was higher during the starter phase (P = 0.005), while during the finisher and the overall period, it was not different from PC (P >0.116). The observed starter phase FCR can be due to the young birds being unable to utilize the high dietary fibre due to physiological limitations. In the current study, inclusion of pre-treated sunflower (TSM) and wheat middlings (TWM) in the test diets at higher levels (NC75 and NC100) linearly reduced BW and BWG throughout the study (P <0.001) but did not affect their FI (P >0.213) relative to PC and NC. The reduction in BW and BWG, especially at the high inclusion rates of the pre-treated ingredients (NC75 and NC100), can partly be attributed to 1) the reduced soluble protein in pre-treated sunflower meal, which might have caused an amino acid imbalance; 2) although non-significant, NC75 and NC100 consumed 9.29% (21-42 d) and 10.82% (0-42 d) less when compared with PC, respectively; and 3) enzymatic pre-treatment of sunflower meal and wheat middlings led to an increased solubilization of xylose, as TWM and TSM had 59.44 and 29.40 μmol/ml of xylose, respectively, and 4 μmol/ml arabinose in TWM. Xylose and arabinose have been shown to reduce broiler chicken performance, as they are directly absorbed in the small intestine, where they affect glucose and lipid utilization by the liver (Regassa et al., 2017). The test diets in the starter phase and overall FCR showed a linear increment with increased inclusion of TSM and TWM, an indication that these pre-treated ingredients were not utilised by the broiler chickens. As C-5 sugars, xylose and arabinose would only be beneficial to the broiler chicken if fermented to organic acids in the ceca.
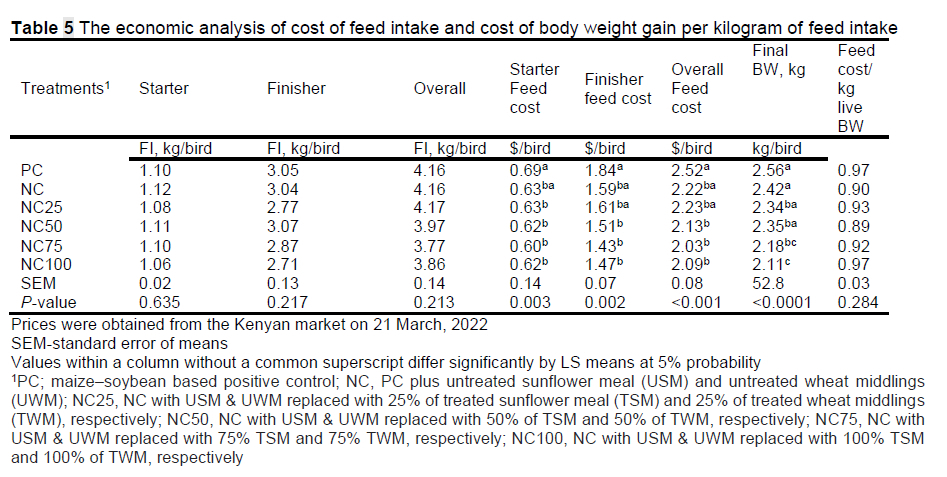
The NC and test diets had 16.35% maize and 31.67% of soybean replaced with cheaper SM and WM and the mean performance of NC, NC25 and NC50 was not different (P >0.05) compared with PC (Table 5), which showed that use of NC and our test diets up to NC50 could save costs for a farmer. The price of starter diets was 623.4, 564.3, 565.6, 567.2, 568.1, and 569.4 USD/ton for PC, NC, NC25, NC50, NC75, and NC100, respectively. The corresponding prices for finisher diets were 601.4, 522.7, 524.6, 526.8, 528.4, and 530.3 USD/ton, respectively (based on ingredient prices in Nairobi at the time of the study). The treated ingredients were given a 10% premium value over the untreated. Compared with PC, the test diets were the cheapest throughout the study (P <0.003). The N50 BWG/kg FI was cheaper during the starter phase (P = 0.020); NC and NC25 were cheapest during the finisher phase and overall study period (d0-42) (P <0.001); cost/kg FI performance of NC, NC25, and NC50 throughout the trial were not different (P >0.05) (Table 5). Therefore, NC and N25 could be economically integrated into the feed formulation for the BW gain comparable to PC. The by-products used in the current study are also readily available all year round and in large quantities in most developing countries (Alshelmani et al., 2021; Mbukwane et al., 2022)
While the efficacy of any feed enzyme is based on performance data obtained from feeding trials (Aftab & Bedford, 2018), it is important to note that pre-treatment has been shown to improve digestibility in in vitro studies. For example, pre-treatment of maize-soybean-based diets with probiotics and subsequently, oven drying and pelleting them, has been shown to work and improve growth performance (Yeh et al., 2018). Therefore, more research is needed to develop an optimum heat processing standard for different pre-treated, non-conventional ingredients as our study has suggested that oven drying of TSM and TWM at 65 °C reduces their nutritional value, thus affecting broiler performance.
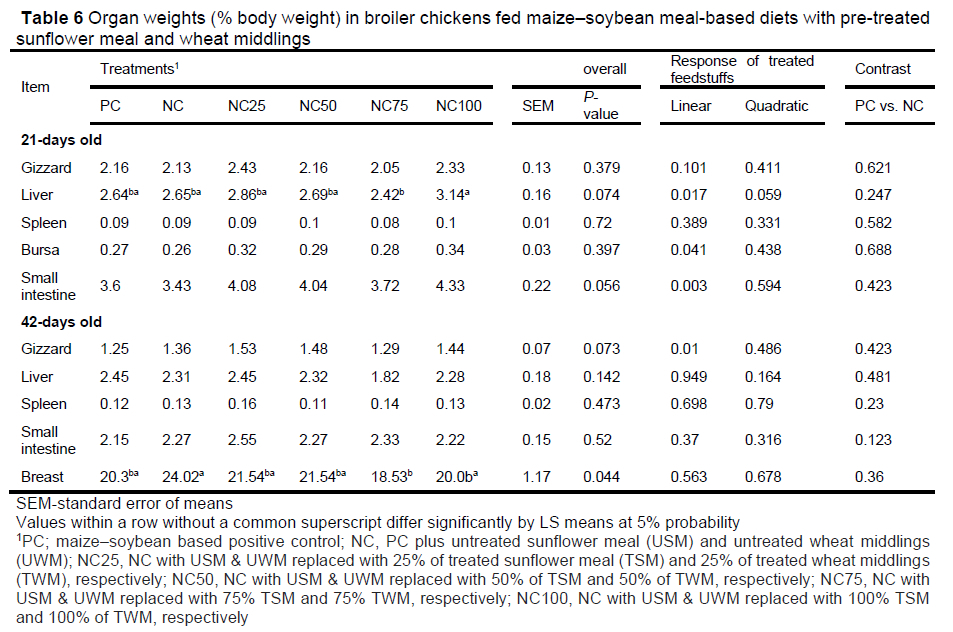
Relative to PC, NC spleen, bursa, gizzard, and small intestine weights were not different throughout the study (P >0.05). This shows that untreated sunflower meal and wheat middlings had no effect on broiler chicken organ weights. On day 21, relative to PC, the gizzard, spleen, and bursa weights on the test diets were not different (P >0.074). However, the liver and small intestine weights increased linearly with the inclusion of pre-treated sunflower meal (TSM) and wheat middlings (TWM) (P = <0.017). On day 42, the gizzard, liver, spleen, and small intestine weights on the test diets were not different when compared with PC (P >0.073), while the breast muscle of the test diets was heavier than the PC (P = 0.044). The increased liver weight in the test diets can be attributed to a higher concentration of xylose and arabinose in TSM and TWM. Inclusion of these mono-sugars in broiler chicken diets has been shown to affect liver hepatic cells, leading to its enlargement (Regassa et al., 2017). The linear increase in the relative weight of the intestine can be due to the fact that the test diets were lower in nitrogen corrected apparent metabolizable energy (AMEn) when compared to PC, and the birds might have responded to this energy deficiency by increasing their mucosal surfaces. Röhe et al. (2020) showed that when dual- purpose laying hens were fed a high dietary fibre diet 2 MJ/kg lower that the control, they responded by increasing their mucosal surface area.
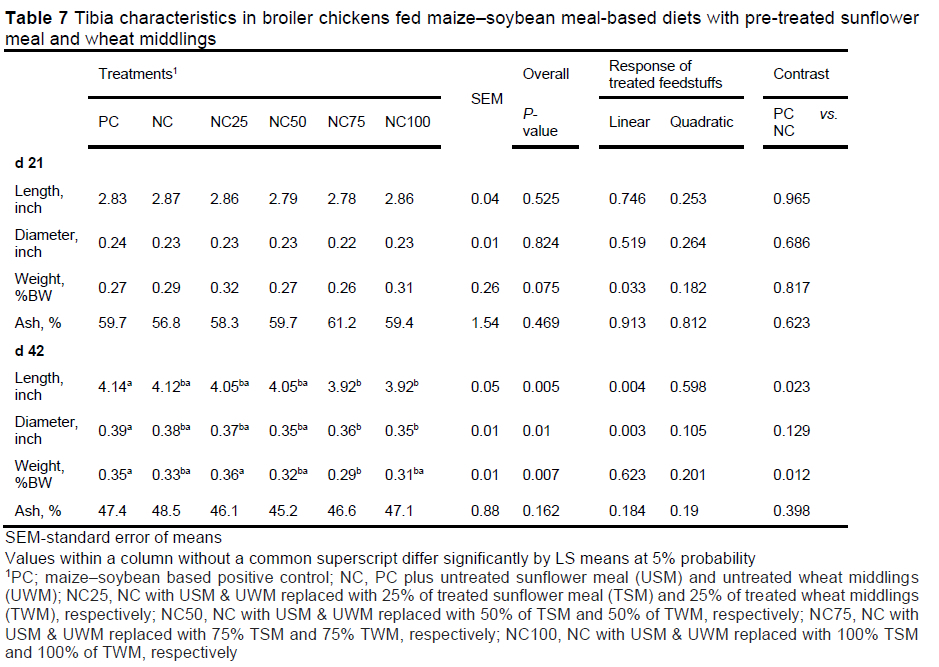
Results of tibia characteristics are shown in Table 7. Tibia has been shown to be the longest and the most mineralised bone in broiler chickens, relative to femur and metatarsus (Han et al., 2014). In the current study, relative to PC, on day 21, NC tibia length, diameter, weight, and ash content were not different (P >0.623). On day 42, tibia length and weight were less than the PC (P <0.023), but tibia diameter and ash were not different (P >0.129). The difference in tibia length and weight could be due allometric growth differences, as PC birds grew faster and were heavier than NC. Relative to PC, the tibia length, diameter, and ash content on the test diets were not different (P >0469), but relative weight decreased linearly with the inclusion of pre-treated sunflower meal and wheat middlings (P =0.033) on day 21. On day 42, tibia length and diameter on the test diets decreased linearly with the inclusion of pre-treated ingredients, while tibia weight was lower, especially at higher inclusion rates (NC75 and NC100) (P =0.007). The ash was not different (P =0.162). These differences can also be attributed to allometric growth rate difference as PC birds were fed diets higher in AMEn, which supported a faster growth. These results are similar to the ones observed by Oikeh et al. (2019), when they suppressed broiler chicken growth rate by diluting their energy with lignocellulose. The tibia length of the birds fed a low energy diet was shorter than the control.
Conclusions
Inclusion of untreated sunflower meal (SM) and wheat middlings (WM) did not affect body weight gain relative to a maize-soybean meal control diet. Inclusion of enzymatically pre-treated sunflower meal and wheat middlings at moderate levels (N25 and N50) in a maize-soybean meal broiler chicken diet did not affect the performance while high levels (N75 and N100) linearly decreased the performance. Based on this study, the cost of producing 1 kg of the broiler chicken meat was the same for all treatments with N50 being numerically cheaper.
Acknowledgments
Financial support from the Federal Government of Canada through the NSERC program, the Ontario Government through Agri-Food Innovation, and the Canadian Bio-systems is appreciated.
Authors contributions
E.G. Kiarie conceptualized and designed the experiment. E.G. Kiarie and C K. Gachuiri read, edited the manuscript, and approved it for publication. F. Muchiri conducted the investigation, analysed the data, and wrote the manuscript.
Conflict of interest declaration
The authors declare that there is no conflict of interest.
References
Abudabos, A. M., 2016. Effect of enzyme supplementation and wheat middlings as an alternative to maize on laying hens performance. http://dx.doi.org/10.4081/ijas.2011.e57 2011, 254-259. https://doi.org/10.4081/IJAS.2011.E57. [ Links ]
Adeola, O., & Cowieson, A. J., 2011. BOARD-INVITED REVIEW: Opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J Anim Sci 89, 3189-3218. https://doi.org/10.2527/JAS.2010-3715. [ Links ]
Aftab, U., & Bedford, M. R., 2018. The use of NSP enzymes in poultry nutrition: Myths and realities. Worlds Poult Sci J 74, 277-286. https://doi.org/10.1017/S0043933918000272. [ Links ]
Ahmadi, K., & Amini, B., 2014. Determination of chemical composition and suitable level of wheat middlings in broiler diets. Int. J. Plant Animal Env. Sci. 4, 454-459. [ Links ]
Almeida, F. N., Htoo, J. K., Thomson, J., & Stein, H. H., 2014. Digestibility by growing pigs of amino acids in heat-damaged sunflower meal and cottonseed meal. J Anim Sci 92, 585-593.https://doi.org/10.2527/JAS.2013-6769. [ Links ]
Alshelmani, M. I., Abdalla, E. A., Kaka, U., & Basit, M. A., 2021. Non-traditional feedstuffs as an alternative in poultry feed. Advances in Poultry Nutrition Research. https://doi.org/10.5772/INTECHOPEN.95946. [ Links ]
CCAC guidelines on the care and use of farm animals in research, teaching and testing. Canadian Council on Animal Care in Science. https://ccac.ca/Documents/Standards/Guidelines/Farm_Animals.pdf [ Links ]
F Oliveira, M. S., Espinosa, C. D., & Stein, H. H., 2021. Heat damage, Maillard reactions, and measurement of reactive lysine in feed ingredients and diets. Proceedings of the Arkansas Nutrition Conference 2021(7).https://scholarworks.uark.edu/panc/vol2021/iss1/7 [ Links ]
Godoy, M. G., Amorim, G. M., Barreto, M. S., & Freire, D. M. G., 2018. Agricultural residues as animal feed. Current Developments in Biotechnology and Bioengineering, 235-256. https://doi.org/10.1016/B978-0-444-63990-5.00012-8. [ Links ]
Hamedi, S., Rezaian, M., & Shomali, T., 2012. Histological changes of small intestinal mucosa of cocks due to sunflower meal single feeding. Am J Anim Vet Sci 6, 171-175. https://doi.org/10.3844/AJAVSP.2011.171.175. [ Links ]
Han, J.C., Qu, H.X., Wang, J.G., Chen, G.H., Yan, Y.F., Zhang, J.L., Hu, F.M., You, L.Y., Cheng, Y.H., 2015. Comparison of the growth and mineralization of the femur, tibia, and metatarsus of broiler chicks. Rev. Bras. Cienc. Avic. 17(3). https://doi.org/10.1590/1516-635x1703333-340. [ Links ]
Horvatovic, M. P., Glamocic, D., Zikic, D., & Hadnadjev, T. D., 2015. Performance and some intestinal functions of broilers fed diets with different inclusion levels of sunflower meal and supplemented or not with enzymes. Braz. J. Poult. Sci. 17, 25-30. https://doi.org/10.1590/1516-635X170125-30. [ Links ]
INRAE-CIRAD-AFZ Feed tables. Tables of composition and nutritional values of feed materials. https://www.feedtables.com/ [ Links ]
Kiarie, E., Romero, L. F., & Ravindran, V., 2014. Growth performance, nutrient utilization, and digesta characteristics in broiler chickens fed maize or wheat diets without or with supplemental xylanase. Poult Sci 93, 1186-1196. https://doi.org/10.3382/PS.2013-03715. [ Links ]
Kiarie, E., Walsh, M., Nyachoti, C.M., 2016. Performance, digestive function, and mucosal responses to selected feed additives for pigs, J Anim Sci, 94(3), 169-180. https://doi.org/10.2527/jas.2015-9835 [ Links ]
Knipfel, J. E., McLeod, J. G., & McCaig, T. N., 1983. Nutritional value of foods and feeds of plant origin: Relationship to composition and processing. pp361-377. https://doi.org/10.1021/BK-1983-0215.CH018. [ Links ]
Mbukwane, M. J., Nkukwana, T. T., Plumstead, P. W., & Snyman, N., 2022. Sunflower meal inclusion rate and the effect of exogenous enzymes on growth performance of broiler chickens. MDPI 12, 253 https://doi.org/10.3390/ani12030253. [ Links ]
Nir, I., Hillel, R., Shefet, G., & Nitsan, Z., 1994. Effect of grain particle size on performance: 2. Grain-texture interactions. Poult Sci. 73, 781-791. https://doi.org/10.3382/PS.0730781. [ Links ]
Nutrient Requirements of Poultry: Ninth Revised Edition, 1994 - National Research Council, Board on Agriculture, Subcommittee on Poultry Nutrition - Google Books. [ Links ]
Oikeh, I., Sakkas, P., Taylor, J., Giannenas, I., Blake, D. P., & Kyriazakis, I., 2019. Effects of reducing growth rate via diet dilution on bone mineralization, performance, and carcass yield of coccidia-infected broilers. Poult Sci. 98, 5477-5487. https://doi.org/10.3382/PS/PEZ400. [ Links ]
Pedersen, M. B., Dalsgaard, S., Arent, S., Lorentsen, R., Knudsen, K. E. B., Yu, S., & Laerke, H. N. 2015. Xylanase and protease increase solubilization of non-starch polysaccharides and nutrient release of maize- and wheat distillers dried grains with solubles. Biochem Eng J. 98, 99-106. https://doi.org/10.1016/J.BEJ.2015.02.036. [ Links ]
Perera, W. N. U., Abdollahi, M. R., Zaefarian, F., Wester, T. J., Ravindran, G., & Ravindran, V., 2019. Influence of inclusion level of barley in wheat-based diets and supplementation of carbohydrase on growth performance, nutrient utilisation, and gut morphometry in broiler starters. Br Poult Sci. 60, 736-748. https://doi.org/10.1080/00071668.2019.1639142. [ Links ]
Ravindran, V. 2013. Feed enzymes: The science, practice, and metabolic realities. J Appl Poult Res. 22, 628-636. https://doi.org/10.3382/JAPR.2013-00739. [ Links ]
Raza, S., Ashraf, M., Pasha, T., Latif, F., 2009. Effect of enzyme supplementation of broiler diets containing varying level of sunflower meal and crude fibre. Pak. J. Bot. 41. [ Links ]
Regassa, A., Kiarie, E., Sands, J. S., Walsh, M. C., Kim, W. K., & Nyachoti, C. M. 2017. Nutritional and metabolic implications of replacing maize starch with D-xylose in broiler chickens fed maize and soybean meal-based diet. Poult Sci. 96, 388-396. https://doi.org/10.3382/PS/PEW235. [ Links ]
Rho, Y., Patterson, R., Joye, I., Martinez, M., James Squires, E., & Kiarie, E. G., 2020. Fibre degrading enzymes increased monosaccharides release and fermentation in maize distillers dried grains with solubles and wheat middlings steeped without or with protease. Transl Anim Sci 4, 1-16. https://doi.org/10.1093/TAS/TXAA153. [ Links ]
Röhe, I., Vahjen, W., Metzger, F., & Zentek, J., 2020. Effect of a "diluted" diet containing 10% lignocellulose on the gastrointestinal tract, intestinal microbiota, and excreta characteristics of dual-purpose laying hens. Poult Sci. 99, 310-319. https://doi.org/10.3382/PS/PEZ492. [ Links ]
Sathitkowitchai, W., Nitisinprasert, S., & Keawsompong, S. 2018. Improving palm kernel cake nutrition using enzymatic hydrolysis optimized by Taguchi method. 3 Biotech 8, 1-7. https://doi.org/10.1007/S13205-018-1433-6/TABLES/3. [ Links ]
Tejeda, O. J., & Kim, W. K. 2020. The effects of cellulose and soybean hulls as sources of dietary fibre on the growth performance, organ growth, gut histomorphology, and nutrient digestibility of broiler chickens. Poult Sci. 99, 6828-6836. https://doi.org/10.1016/J.PSJ.2020.08.081. [ Links ]
Teodorowicz, M., Hendriks, W. H., Wichers, H. J., & Savelkoul, H. F. J., 2018. Immunomodulation by processed animal feed: The role of Maillard reaction products and advanced glycation end-products (AGEs). Front Immunol. 9, 2088. https://doi.org/10.3389/FIMMU.2018.02088/BIBTEX. [ Links ]
Yeh, R. H., Hsieh, C. W., & Chen, K. L., 2018. Screening lactic acid bacteria to manufacture two-stage fermented feed and pelleting to investigate the feeding effect on broilers. Poult Sci. 97, 236-246. https://doi.org/10.3382/PS/PEX300. [ Links ]
Submitted 13 April 2022
Accepted 10 September 2022
Published 27 February 2023
# Corresponding author: ekiarie@uoguelph.ca


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}