










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 n.6 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i6.08
Dietary supplementation with vegetable oils with low n-6:n-3 polyunsaturated fatty acid ratios improves the intramuscular fat and fatty acid composition of growing-finishing pigs
S.S. JiangI, II; F. WeiI; D.L.Y.E. AbulahaitiI; C.C. TianI; L. FuI; J.X. LuI, #; G.H. ZhangI, #
ICollege of Life Science and Engineering, Northwest Minzu University, Lanzhou 730030, China
IIDepartment of Animal Science and Technology, Gansu Agriculture Technology College, Lanzhou 730000, China
ABSTRACT
This study investigated the effects of different n-6:n-3 polyunsaturated fatty acid (PUFA) ratios in pig diets using vegetable oil supplementation on growth performance, carcass characteristics, meat quality, and fatty acid composition in the longissimus dorsi muscle (LM) and subcutaneous adipose tissue (SAT) of pigs. Fifty-four cross-bred growing pigs (Large White χ Landrace χ Duroc; 45.03 ± 1.72 kg) were randomly assigned to one of three isoenergetic diets: the 1) basal diet (control), 2) RAPO diet (diet supplemented with 4.5% rapeseed oil), and 3) MIXO diet (diet supplemented with 2.25% rapeseed oil and 2.25% linseed oil), with n-6:n-3 PUFA ratios of approximately 13:1, 7:1, and 2:1, respectively. The experiment lasted 42 days. The RAPO and MIXO diets did not affect the growth performance and carcass characteristics of pigs, while the content of low-density lipoprotein (LDL), triglycerides (TGs), and glucose (GLU) in the serum decreased substantially. Intramuscular fat (IMF) content increased by 19.25% and 20.11% in the LM of pigs fed the RAPO and MIXO diets, respectively, and lower cooking loss and drip loss of meat were observed only in pigs fed the MIXO diet. The RAPO and MIXO diets decreased the stearic acid and palmitoleic acid levels, increased the total PUFA levels (including those of α-linolenic and linoleic acid), and decreased the n-6:n-3 PUFA ratios in LM and SAT. However, the MIXO diet was more effective in improving the fatty acid composition. A mixed oil diet with an n-6:n-3 PUFA ratio of 2:1 is an effective measure for improving meat quality.
Keywords: meat quality, growth performance, intramuscular fat, fatty acids, pig
Introduction
Fat mass and distribution are essential factors determining the economic traits of pigs. Reduction in subcutaneous fat content increases carcass value, and intramuscular fat (IMF) is known for "marbling" and is positively associated with various meat quality traits, such as juiciness, flavour, and tenderness (Dannenberger et al., 2012). Previous studies have proposed that a minimum level of 2.5% IMF is needed to positively influence the eating quality and sensory quality traits, thereby increasing consumer acceptance of pork (Alfaia et al., 2019). In addition, the type and content of fatty acids in pork have received much attention due to the nutritional value and health benefits of fatty acids in the human diet, especially unsaturated fatty acids (Puig-Oliveras et al., 2014). However, genetic selection for increased meat yield has resulted in a less than 1.5% IMF content in most modern pig breeds and has altered the fatty acid composition of meat (Hernández-Sánchez et al., 2013; Ba et al., 2019), and typical corn-soybean meal-based diets lead to a high ratio of n-6:n-3 PUFA in pork (Palmquist, 2009). A high n-6:n-3 PUFA ratio in the human diet could be a factor inducing cardiovascular, inflammatory, and metabolic disorders (Martínez-Fernández et al., 2015). There is growing interest in producing pork with a high IMF content, balanced fatty acid composition, and low subcutaneous fat content (Alfaia et al., 2019).
Dietary fat sources and their n-6:n-3 PUFA ratios play an essential role in altering the fatty acid composition in tissues, maintaining healthy lipid metabolism, and promoting animal production performance. Studies have focused on altering the fatty acid composition of pork by supplementing the diet with natural sources of PUFAs, such as fish oil or fish meal, different linseed products, rapeseed oil, and soybean oil (Realini et al., 2010; Okrouhla et al., 2018), which could be suitable fat sources for enriching n-3 PUFAs in animal products. For example, the inclusion of 3% extruded linseed or linseed oil in the diet improved the fatty acid profile of pork without affecting growth performance (Hăbeanu et al., 2014; Tarricone et al., 2020). However, different vegetable oils or n-6:n-3 PUFA ratios in diets had inconsistent effects on the IMF content and fatty acid composition of pigs. A meta-analysis showed that dietary linseed supplementation increased the levels of n-3 PUFAs, such as a-linolenic acid, docosapentaenoic acid (DPA), docosahexaenoic acid (DHA), and eicosapentaenoic acid (EPA), in muscle and adipose tissue (Corino et al., 2014). Okrouhlá et al. (2018) reported that rapeseed oil was better than soybean oil in improving the fatty acid composition and reducing the n-6:n-3 PUFA ratio in subcutaneous fat. Dietary n-6:n-3 PUFA ratios of 4:1 and 2:1 substantially increased the proportions of essential fatty acids and reduced the cooking loss ratio, but low expression of acetyl COA carboxylase (ACC) with a dietary n-6:n-3 PUFA ratio of 2:1 indicated a reduced capacity for de novo fatty acid synthesis (Song et al., 2020). Palmitic, oleic, and linoleic acids, as dietary fat sources, have been confirmed to promote lipid accumulation in porcine adipocytes through different metabolic pathways, which could be related to the phosphorylation of peroxisome proliferator-activated receptor γ (PPARy) (Yu et al., 2017). Moreover, an appropriate n-6:n-3 PUFA ratio in the diet is beneficial in improving lipid metabolism and the inflammatory system to ensure the efficient utilization of energy and nutrients (Duan et al., 2014). Given the role of dietary n-6 and n-3 PUFAs and their ratio in regulating fat metabolism and health benefits, their impact on the improvement of meat quality needs to be elucidated.
The purpose of this study was to investigate whether the addition of vegetable oils with different n-6:n-3 PUFA ratios to the diet as a partial carbohydrate replacement could improve the production performance, health status, and meat quality in finishing pigs; and to further clarify the effects of dietary n-6:n-3 PUFA ratios on fatty acid metabolism in longissimus dorsi muscle (LM) and subcutaneous adipose tissue (SAT).
Materials and Methods
This study was approved by the Animal Care and Use Committee of Northwest Minzu University (Lanzhou, China; ethical clearance number: 20180307). A total of 54 barrows (Large White χ Landrace χ Duroc) at approximately 13 weeks of age, with an initial body weight of 45.03 ± 1.72 kg, were used. The pigs were provided by Lanzhou Ruiyuan Agricultural Technology Co. Ltd (Lanzhou, China). Pigs were allotted randomly to three treatments, and each treatment had six replicates and three pigs per pen. Pigs were housed in an enclosed building with a partially-slatted concrete floor. Each pen provided 4.68 m2 floor space with a size of 2.6 m χ 1.8 m and was equipped with a stainless-steel feeder and nipple drinkers. Feed and fresh water were provided adíibitum. During the experimental period, the average temperature inside the building was 20-28 °C (average 24 °C), and the relative humidity was 56 ± 6%. All pigs were fed twice daily (at 08:00 and 17:00). Pigs were given a 7-day adaptation period before the experiment. The experiment lasted 42 days.
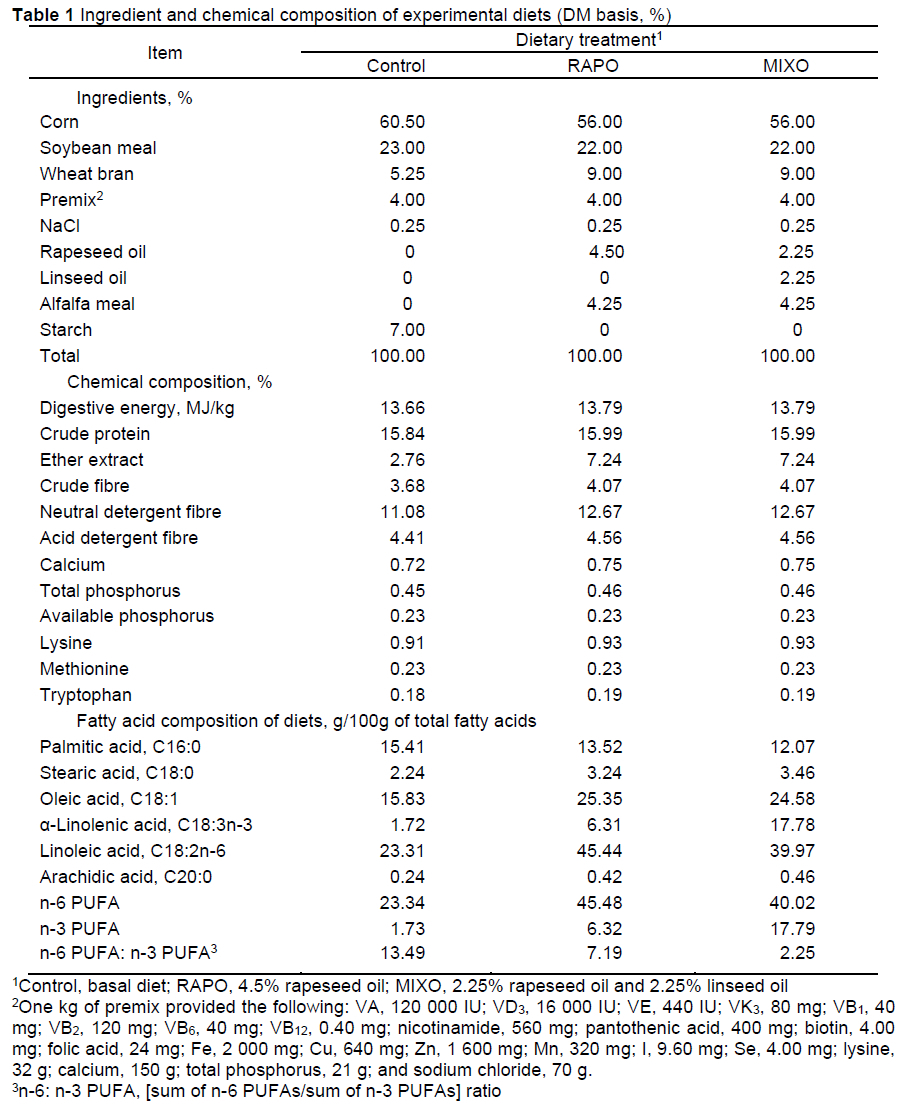
Each group was fed one of three isoenergetic diets: the control diet (high-carbohydrate diet), RAPO diet (diet supplemented with 4.5% rapeseed oil), and MIXO diet (diet supplemented with 2.25% rapeseed oil and 2.25% linseed oil) with an n-6:n-3 PUFA ratio of approximately 13:1, 7:1 and 2:1, respectively. Diets were formulated to meet the requirements proposed by the NRC (2012). The contents of digestible energy (DE), crude protein (CP), and acid detergent fibre (ADF) in the diets were approximately equal. Feed samples were taken from each dietary treatment for chemical analysis. The ingredients, chemical composition, and fatty acid composition of each diet are shown in Table 1.
Feed consumption was recorded daily per pen. Data were collected to calculate the initial body weight, final body weight, average daily gain (ADG), average daily feed intake (ADFI), and feed conversion ratio (FCR).
At the end of the experiment (42 days), all pigs were weighed individually before the morning feeding. Approximately 5 mL of blood was collected from the anterior vena cava of pigs, centrifuged at 2,000 χ g for 5 min, and then frozen at -20 °C until the assay was conducted. Ten pigs with weights closest to the average weight of each group were chosen, for a total of 30 pigs. The pigs were electrically stunned (225 to 380 V/0.5 A for 5 to 6 s), slaughtered, scalded (4 min, 63 °C), dehaired, and eviscerated. Samples of the LM were taken from the 3rd to 11th rib on the left half of the carcasses to determine meat quality parameters. Samples of the LM and SAT were rapidly collected from the 13th to 14th ribs on the right side of the carcass, frozen in liquid nitrogen and stored at -80 °C until fatty acid composition analysis and gene expression analysis. Backfat thickness was measured on the right side of the carcass as the average value for three points: the shoulder, the last rib, and the last lumbar vertebra. Loin eye area was measured with the maximum width (cm) and height (cm) of the exposed surface muscle area between the 12th and 13th ribs on the right side of the carcass and was calculated using the equation:

according to the Chinese Guidelines on Performance Measurement Technology and Regulations for Pigs (China, 2014).
After 24 h of refrigeration at 4 °C, the meat colour was measured using a colorimeter (CR-400, Minolta Co., Osaka, Japan), and the average values of lightness (L*), redness (a*), and yellowness (b*) were determined from five readings. The pH values were measured with a pH meter (MP 220, Mettler-Toledo, Schwarzenbach, Switzerland). Drip loss was determined using the suspension method. To measure drip loss, the meat samples were trimmed to remove the peripheral muscular membranes and cut into blocks (approximately 2 χ 2 χ 2 cm3) along the direction of muscle fibres. The meat pieces were suspended in plastic bags and stored at 4 °C for 24 h. The drip loss was calculated as a percentage of weight loss compared to the initial weight, using the average of three experiments. To evaluate the shear force, any visible external fat and connective tissue were removed from the samples, which were individually packed in plastic bags and heated in a water bath until the core temperature reached 70 °C. After the samples were cooled to room temperature (25 °C), each cooled sample was cut into three slices, parallel to the fibre orientation (1.27 cm in diameter and ~3 cm in length) and measured using a shear apparatus (C-LM3, Northeast Agricultural University, Harbin, China). After weighing, the samples were placed in plastic bags and boiled in water until the internal temperature reached 70 °C. Cooking loss was calculated as the percentage of weight loss after cooking. The water loss ratio was measured using a Model RH-1000 instrument (RH-1000, RunHu Co., Guangdong, China).
The samples were cut into a circular meat sample with an area of 5 cm2 and a thickness of 1 cm, weighed and placed between gauze and 18 layers of qualitative filter paper. Then, the circular meat samples were pressurized to 35 kg (stress of 138.8 kPa) for 5 min and weighed. The water loss percentage was calculated as the fluid lost compared to the initial meat weight.
IMF content was determined in LM using the Soxhlet extraction method. The fatty acid composition was determined using gas chromatography, as previously described (Xu, et al., 2019). Briefly, lipids were extracted from samples using 2:1 chloroform:methanol according to the Folch method (Folch et al., 1957). Then, total lipids were converted into fatty-acid methyl esters (FAMEs) and analysed using gas chromatography (GC-2010 Plus, Shimadzu Co., Kyoto, Japan) equipped with a flame ionization detector (FID) and a SP2560 capillary column (100 mm χ 0.25 mm, 0.2 μm). The results are expressed as a percentage of total fatty acids.
Serum levels of triglycerides (TGs), total cholesterol (TC), high-density lipoprotein (HDL), low-density lipoprotein (LDL), glucose (GLU), serum creatinine (Scr), and blood urea nitrogen (BUN) were analysed using an automatic biochemical analyser (7100 Automatic Analyzer, Hitachi, Tokyo, Japan). Fasting serum insulin (INS) levels were analysed using an enzyme-linked immunosorbent assay (ELISA).
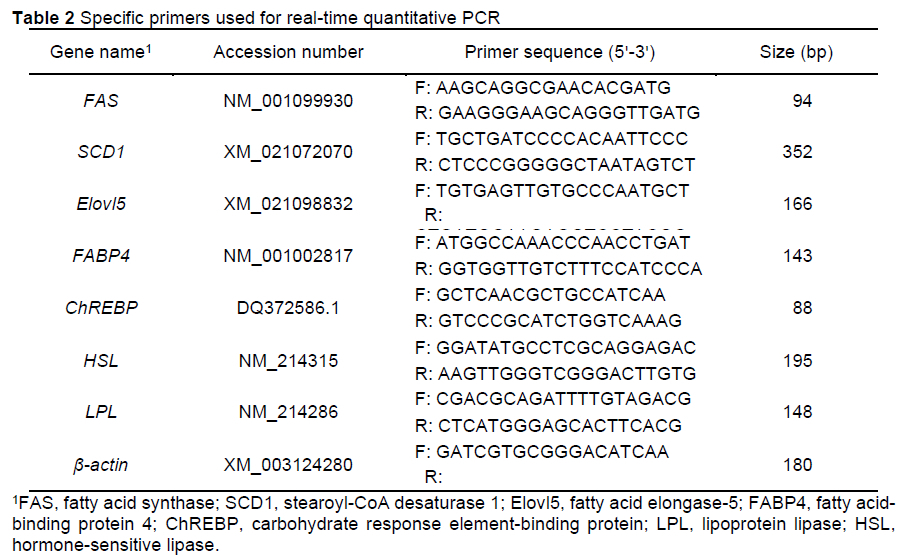
Total RNA was extracted from the LM and SAT samples using TRIzol reagent (Fermentas RNA Extraction Kit). RNA concentration and purity were evaluated by a NanoDrop 2000 (Thermo Fisher Scientific, Waltham, USA). Reverse transcription was performed using the TaqMan Reverse Transcription Reagent. Real-time, quantitative polymerase chain reaction (qPCR) was performed using a CFX96 real-time PCR detection system (Bio-Rad, Hercules, CA, USA). The PCR system was set as follows (total reaction system: 10 μΙ): 5.0 μΙ of SYBR Premix Ex Taq™ (TaKara, Kyoto, Japan), 0.4 μl of each primer, 1.0 μl of cDNA solution, and 3.2 μl of sterile double-steamed water. The PCR conditions were 40 cycles at 95 °C for 30 s (denaturation), 60 °C for 30 s (annealing), and 72 °C for 30 s (extension). Relative quantification was performed with the 2_ΔΔCt method. Primers were designed using Primer Premier 5.0 software (Table 2).
Data were analysed using one-way analysis of variance (ANOVA); the Duncan method was used to test multiple comparisons using SPSS statistical software (SPSS Inc., Chicago, IL, USA). The data in the tables are presented as the means and standard error of the mean (SEM). Results were considered significant when the P value was <0.05, and P values within 0.05 and 0.10 were viewed as tendencies.
Results and Discussion
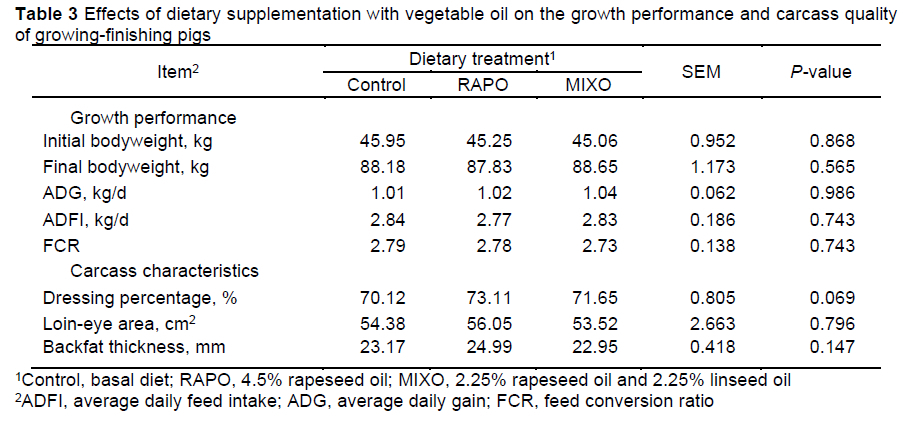
There were no differences (P >0.05) in the initial body weight, final body weight, ADG, ADFI, and FCR of pigs among the dietary treatments (Table 3). Compared with the control group, no differences (P >0.05) were observed in the loin-eye area and average backfat thickness, while the backfat thickness decreased 8% in pigs fed the MIXO diet compared with pigs fed the RAPO diet. Pigs fed the RAPO diet and MIXO diet tended to have an increased dressing percentage compared to pigs fed the control diet (P = 0.069).
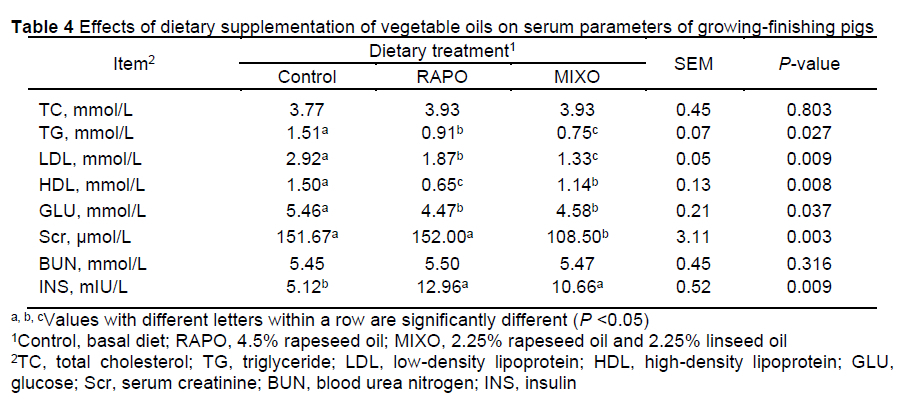
The serum levels of TC and BUN were not affected by dietary treatment (P >0.05) (Table 4). The RAPO and MIXO diets decreased serum TG, GLU, HDL, and LDL levels and increased serum INS levels (P <0.05). Compared with RAPO diet, MIXO diet decreased serum Scr and LDL levels, whereas serum HDL levels increased (P <0.05).
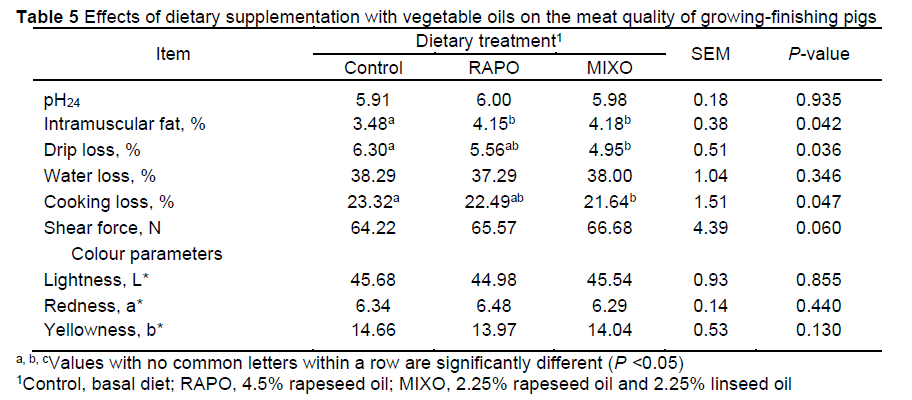
The effects of dietary vegetable oils on meat quality are shown in Table 5. The experimental diets had no effect on the pH at 24 h, meat colour (including the L*, a* and b* values), water loss ratio, and shear force (P >0.05). Compared with the control diet, the MIXO diet decreased drip loss and cooking loss (P <0.05), while there was no difference between the MIXO and RAPO diets. Compared with the control diet, the RAPO and MIXO diets increased the IMF content by 19.25% and 20.11%, respectively (P <0.05).
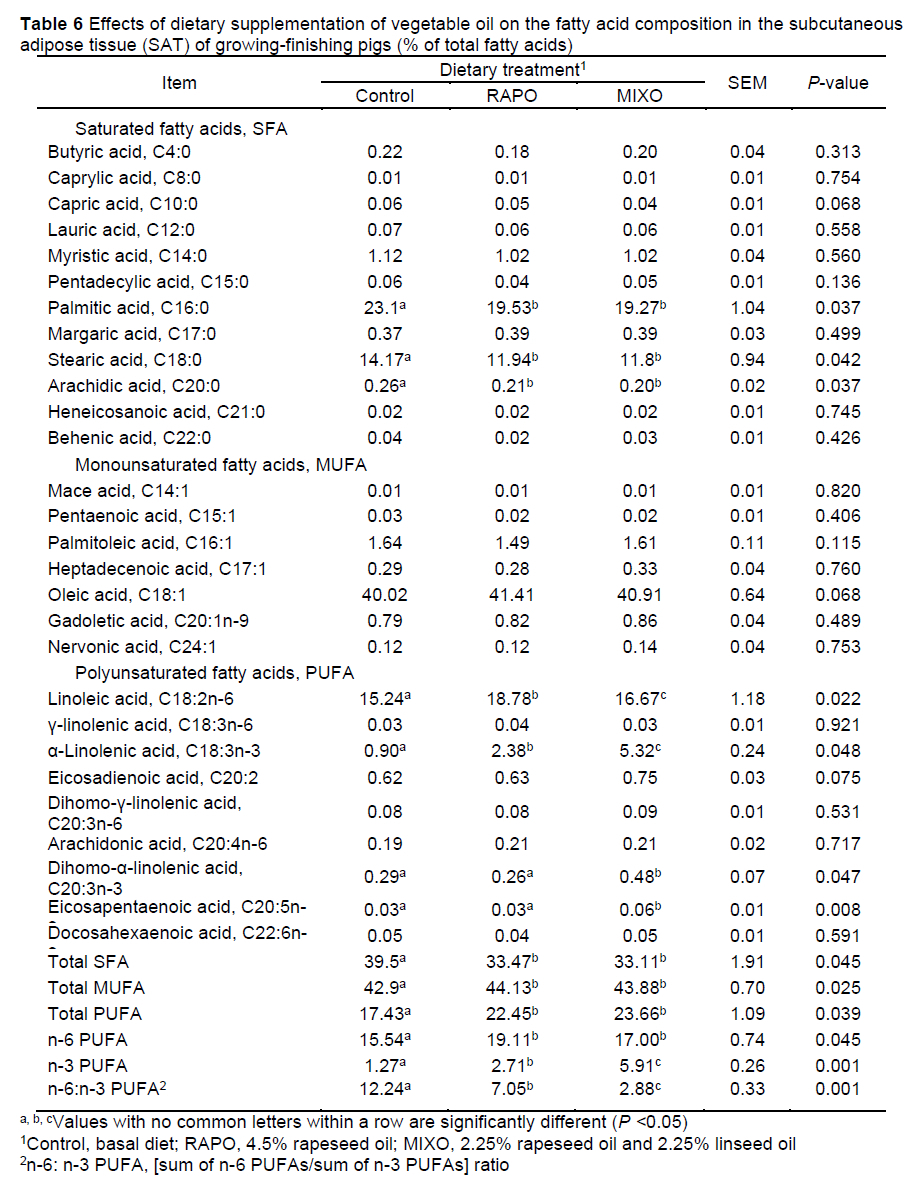
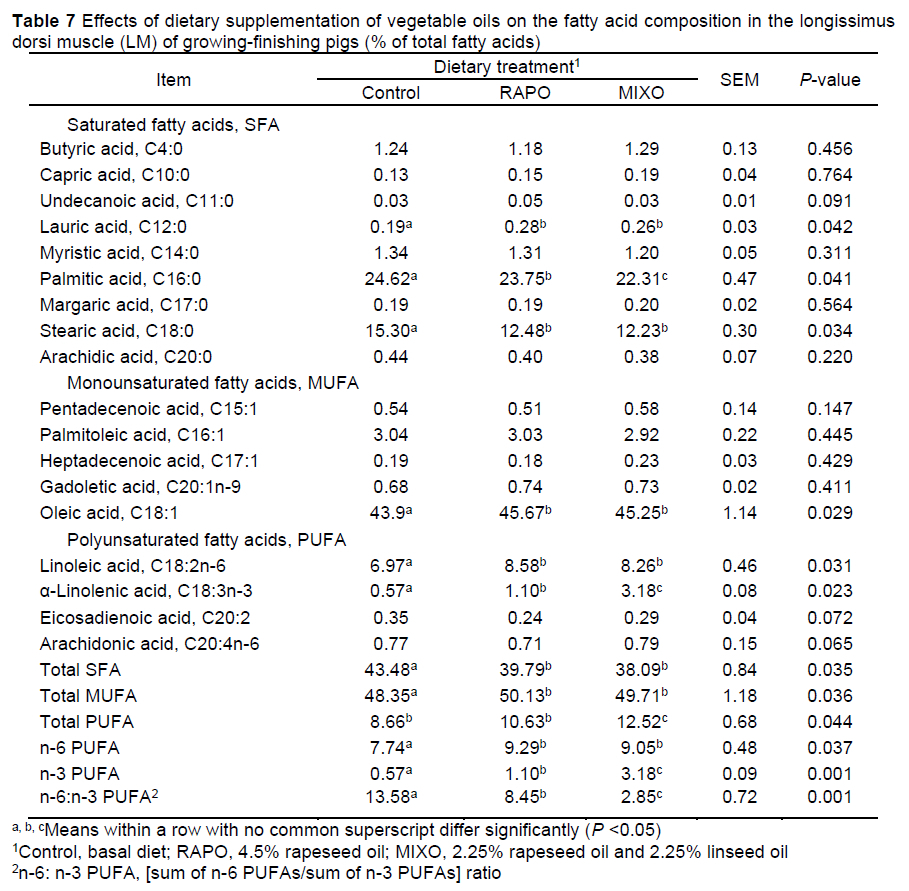
Compared with the control diet, both the MIXO and RAPO diets decreased the levels of C16:0, C18:0 and total saturated fatty acids (SFAs) (P <0.05), increased the total monounsaturated fatty acid (MUFA) content (P <0.05), and tended to increase the C18:1 content in the SAT of pigs (P = 0.068) (Table 6). In the LM, the levels of C16:0 and C18:0 and the total SFA content were decreased, and the levels of C12:0 and C18:1 and the total MUFA content were increased, by feeding with the MIXO and RAPO diets (P <0.05) (Table 7). Feeding with the MIXO and RAPO diets increased the linoleic acid (C18:2n-6), α-linolenic acid (C18:3n-3), total n-6 PUFA, and total n-3 PUFA levels (P <0.05) in both SAT and LM, resulting in a marked decrease in the n-6:n-3 PUFA ratio (P <0.05). The α-linolenic acid content and lower n-6:n-3 PUFA ratio in the SAT and LM of pigs fed the MIXO diet were higher than those in pigs fed the RAPO diet (P <0.05), but there was no difference in linoleic acid content in the LM (P >0.05).
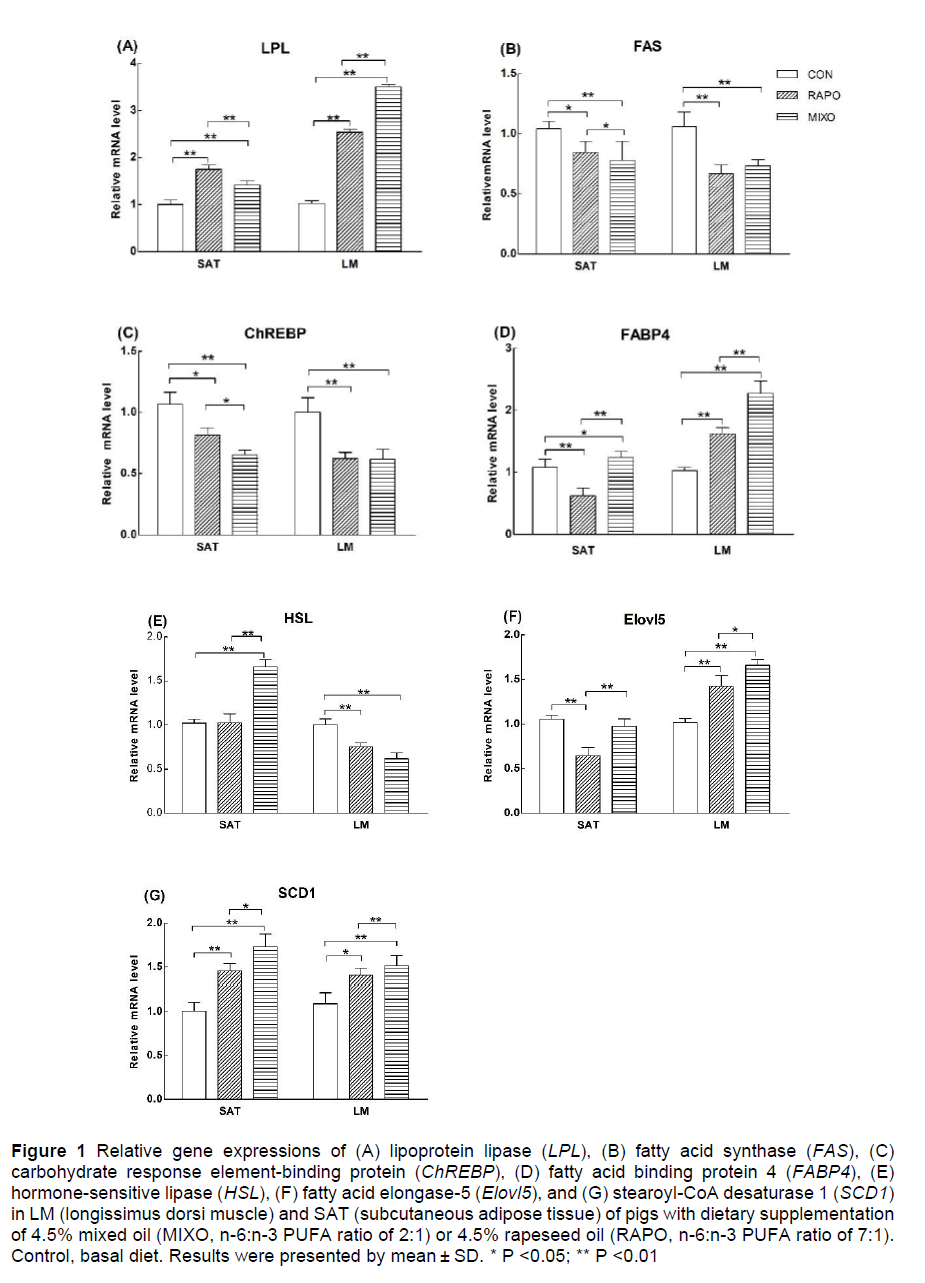
The expression of genes related to lipid metabolism was analysed (Figure 1), including lipoprotein lipase (LPL), glucose-responsive transcription factor (ChREBP), fatty acid synthase (FAS), fatty acid binding protein 4 (FABP4), hormone-sensitive lipase (HSL), stearoyl-CoA desaturase 1 (SCD1) and fatty acid elongase-5 (Elovl5). In SAT and LM, the expression levels of SCD1 and LPL were upregulated (P <0.05), and the expression levels of ChREBP and FAS were downregulated (P <0.05), by feeding with the MIXO and RAPO diets, compared with the control diet. The expression levels of FABP4 and Elovl5 were lower in the SAT of pigs fed the RAPO diet than in that of pigs fed the control or MIXO diet (P <0.01), whereas their expression levels were higher in the LM of pigs fed the RAPO or MIXO diets than in the pigs fed the control diet. The HSL expression in SAT was markedly increased by the MIXO diet compared with the control and RAPO diets, but its expression in the LM was decreased by the MIXO and RAPO diets, compared with the control diet (P <0.01).
This study showed no marked effect of vegetable oil supplementation on the ADG, ADFI, FCR, loin-eye area, or backfat thickness but a tendency to increase the dressing percentage in the 42-day trial. Similar to our results, diets supplemented with 4% rapeseed oil or soybean oil (Vehovsky et al., 2019) or with 10% linseed oil or sunflower oil (Realini et al., 2010) did not affect ADG, ADFI, or carcass traits in pigs. Conversely, Nong et al. (2020) reported that a diet with a lower n-6:n-3 PUFA ratio (3:1 vs. 5:1 and 8:1) increased the F:G ratio in Heigai pigs (a local fatty breed). Duan et al. (2014) reported that the final body weight and ADG were substantially increased when pigs were fed a mixed oil diet with an n-6:n-3 PUFA ratio of 5:1 (0.75% linseed oil and 2.25% soybean oil). These discrepant results could be due to differences in genetic background, feeding periods, dietary composition, dietary levels of n-3 PUFAs, or environmental conditions in the previous studies. In the current study, a slight increase in the dressing percentage of the pigs was observed when the diets were supplemented with 4.5% rapeseed oil or mixed oil, which could be related to the n-3 PUFA-enriched diets improving energy utilization by increasing insulin activity (Palmquist, 2009).
The LDL is a primary cholesterol carrier in blood for delivering cholesterol to tissues and a key indicator for disease diagnosis, therapy, and monitoring of disease progression. In this study, we found that supplementation with rapeseed oil and mixed oil substantially decreased serum levels of LDL, HDL, GLU, and TG without affecting TC (especially serum LDL and TG levels, which were most significantly decreased in the mixed oil diet). This suggests that reducing the dietary n-6:n-3 PUFA ratio from 7:1 to 2:1 could further improve glucose and lipid metabolism in pigs. Consistent with a previous study (Song et al., 2020), a low dietary n-6:n-3 PUFA ratio effectively decreased serum concentrations of LDL and TG. Recent studies have confirmed that a low dietary ratio of n-6:n-3 PUFA and total consumption of n-3 PUFA are beneficial for improving obesity-induced inflammation and insulin resistance (Fan et al., 2020). In the current study, we observed that a dietary n-6:n-3 ratio of 2:1 resulted in increased expression of LPL, the rate-limiting enzyme for the clearance of circulating TGs in the LM (Wang and Eckel, 2009). However, Nguyen et al. (2019) reported that a decrease in the dietary n-6:n-3 PUFA ratio from 17:1 to 5:1 did not affect serum HDL, TC, or TG levels. Song et al. (2020) reported that dietary n-6:n-3 PUFA ratios of 2:1 and 4:1 decreased serum TC levels, while the addition of 10% sunflower oil or coconut oil tended to increase serum concentrations of HDL, LDL, and TC (Iyer et al., 2012). These divergent results might be partly due to differences in the total fat content and fatty acid composition of the diet.
Although the mixed oil diet reduced the serum creatinine level, the value fell in the normal physiological range (70-203 μmοΙ/Ι_) (Friendship et al., 1984), suggesting that this diet had no adverse effect. Hăbeanu et al. (2019) reported that diets rich in n-3 PUFAs substantially decreased the serum creatinine concentration in barrows and were related to increased retention and utilization of nitrogen. The results indicated that a dietary n-6:n-3 PUFA ratio of 2:1 could further improve the metabolism of glucose, fat, and nitrogen in finishing pigs, which would have additional potential benefits for the health and productivity of pigs.
The effect of dietary lipids on IMF content and backfat thickness in pigs is still controversial. Diets supplemented with lipids consisting of 1% linseed oil and 1-5% poultry fat (Huang et al., 2020), 4% rapeseed oil or soybean oil (Vehovsky et al., 2019), or a low dietary n-6:n-3 PUFA ratio (Nong et al., 2020) had no effect on the IMF content or backfat thickness of pigs. However, Li et al. (2015) reported that the IMF content was higher in pigs fed diets with n-6:n-3 PUFA ratios of 2.5:1, 5:1, and 10:1 than pigs fed diets with an n-6:n-3 PUFA ratio of 1:1 but had no effect on backfat thickness. In our study, the IMF content increased by 19.25% and 20.11% in the LM of pigs fed the RAPO and MIXO diets, respectively. These findings suggest that the dietary n-6:n-3 PUFA ratio could be a key factor of vegetable oils in affecting the IMF content of pork; ratios of 2:1 and 7:1 were suitable for improving the IMF content.
In the current study, ChREBP, a glucose-responsive transcription factor, was significantly downregulated, indicating that the de novo fatty acid synthesis pathway promoted by glucose could be downregulated (Gregoire et al., 1998). The downregulation of FAS expression was accompanied by the upregulation of FABP4 in the RAPO and MIXO diets, implying that vegetable oil supplementation would increase fatty acid transport and decrease fatty acid synthesis in the LM (Boss et al., 2015; Wang et al., 2015). In addition, although the dietary vegetable oils had no significant effect on the backfat thickness of pigs, feeding with the MIXO diet decreased the backfat thickness by 8%, compared with the RAPO diet. Pigs fed the mixed oil diet showed increased expression of HSL in SAT, indicating an increase in the lipolysis of TGs stored in adipose tissues (Watt et al., 2006). This is consistent with the findings of Song et al. (2020). The diets with different n-6:n-3 PUFA ratios had different effects on fat deposition in SAT and LM, suggesting that fat deposition in these tissues might be differentially regulated by diets.
Although the underlying mechanism can be complex, several studies have reported that high n-3 PUFA levels can affect fat metabolism by regulating the expression of lipogenic genes (Ogłuszka et al., 2017). A diet rich in the n-3 PUFAs stimulated the expression of genes involved in the regulation of muscle metabolism and affected the interaction between lipogenesis and oxidative processes in the longissimus thoracis muscle of pigs (Vitali et al., 2018). N-3 PUFAs selectively drove the expansion of adipocyte numbers to control adipogenesis by activating the n-3 fatty acid receptor, FFAR4 (Hilgendorf et al., 2019). The heritability estimates for linoleic and arachidonic acids in IMF and SAT reveal that the linoleic acid metabolic pathway facilitates an efficient increase in the IMF content in pigs (Gol et al., 2019).
In our study, the pH value, meat colour, water loss ratio, and shear force in the LM were not affected by vegetable oil supplementation in the diets. One unanticipated finding was that the drip loss and cooking loss were substantially decreased in the LM of pigs fed the mixed oil diet. Similarly, Song et al. (2020) reported that cooking loss in the LM was decreased by linseed-supplemented diets with 2:1 and 4:1 ratios of n-6:n-3 PUFA, while the pH, colour parameters, drip loss, and shear force were not influenced. However, studies have reported that incorporating 1% linseed oil (Huang et al., 2020) or 4% rapeseed oil or soybean oil (Vehovsky et al., 2019) to the diets had no influence on the IMF content, meat colour, shear force, or drip loss in pigs; even dietary inclusion of 20% rapeseed meal had no effect on drip loss in the loin and belly (Skugor et al., 2019). In the current study, the decreased drip loss and cooking loss of meat might be related to the increased IMF content, which will eventually improve the edible value of pork. Overall, it can be concluded that a 4.5% mixed oil in the diet had a positive effect on meat quality.
Previous studies have shown that linseed oil is richer in α-linolenic acids compared to other plant-derived oils, which increases the n-3 PUFA content in tissues and modifies the lipid profile (De Tonnac et al., 2017). We observed that an increase in the linoleic acid and α-linolenic content in the LM and SAT substantially decreased the n-6:n-3 PUFA ratio. In SAT, the rapeseed oil diet with an n-6:n-3 PUFA ratio of 7:1 increased the α-linolenic and linoleic acid levels by 164% and 23%, respectively, resulting in a low n-6:n-3 PUFA ratio (7.05 vs. 12.24), and the mixed oil diet with an n-6:n-3 PUFA ratio of 2:1 increased the α-linolenic and linoleic acid levels by 490% and 9%, respectively, resulting in a lower n-6:n-3 PUFA ratio (2.88 vs. 12.24), compared with a basal diet rich in carbohydrates. Similarly, dietary n-6:n-3 PUFA ratios of 4:1 and 2:1 led to a nearly 4fold increase in the α-linolenic acid content in adipose tissue (Song et al., 2020). In LM, the rapeseed oil diet increased the α-linolenic and linoleic acid levels by 92% and 23%, respectively, and the n-6:n-3 PUFA ratio decreased from 13.58 to 8.45; the mixed oil diet increased the α-linolenic and linoleic acid levels by 457% and 18%, respectively, and the n-6:n-3 PUFA ratio decreased to 2.85. Consistent with our results, diets supplemented with rapeseed oil substantially increased the linoleic acid and α-linolenic acid proportions in IMF (Vehovsky et al., 2019), and more n-6 and n-3 PUFAs accumulated in the LM when pigs were fed a 10% linseed diet (Huang et al., 2008). These results suggest that both rapeseed and mixed oil diets can substantially increase the α-linolenic acid and linoleic acid levels and reduce the n-6:n-3 PUFA ratio in tissues. However, a mixed oil diet with an n-6:n-3 PUFA ratio of 2:1 appeared to be more effective in improving fatty acid composition, which is supported by the opinion that dietary fatty acids can be incorporated into these tissues with few modifications (Leikus et al., 2018).
In addition, the rapeseed and mixed oil supplementation decreased the content of C16:0, C18:0 and total SFAs and increased the total MUFA and C18:1 content in SAT and LM, and the C12:0 level in the LM. Similarly, the contents of C14:0, C16:0, C16:1, and C18:1 in the LM and SAT of pigs were affected by the vegetable oil type or n-6:n-3 PUFA ratios (Nuernberg et al., 2005; Realini et al., 2010). In the current study, we found that dietary vegetable oils increased the expression of SCD1 in SAT and in LM, and the expression of Eíoví5 in the SAT of pigs fed the RAPO diet was lower than that in the SAT of pigs fed the MIXO diet. These two enzymes can catalyse the synthesis of MUFAs and long-chain fatty acids in tissues (Gregory et al., 2011). Fan et al. (2020) reported that reducing the n-6:n-3 PUFA ratio in the diet is a key determinant in promoting n-3 PUFA biosynthesis and subsequent lipidome modifications. Considering the effects on IMF, blood glucose and insulin, it can be reasonably postulated that the synthesis and conversion of fatty acids was also affected by the diets. This finding is supported by other studies. Doran et al. (2006) found that vegetable oil types differentially affected SCD activity and SCD protein expression in the muscle and adipose tissue, and SCD catalysed the formation of MUFAs (mainly 16:1 and 18:1) from the corresponding SFAs (16:0 and 18:0). The dietary n-6:n-3 PUFA ratios of 1:1-5:1 produced different effects on the expression level of genes involved in fatty acid metabolism in the LM and SAT of pigs, such as fatty acid transport protein (FATP)-1, FATP-4 and ACC (Li et al., 2015), which encode key enzymes for the de novo biosynthesis of fatty acids. The n-3 PUFAs resulted in increased expression of genes encoding fatty acid synthesis, desaturation, and elongation in the longissimus thoracis muscle of pigs, including ACC, ChREBP, ELOVL6, FASN, and SCD (Vitali et al., 2018). Thus, the effects of dietary PUFAs on the fatty acid profile of muscle and adipose tissue can include both direct incorporation into and effects on fatty acid metabolism, and the metabolism is different in both tissues (Hernández-Sánchez et al., 2013).
Moreover, we observed that although supplementation with vegetable oil substantially increased the α-linolenic acid content, it had little effect on the DHA and EPA levels. Alpha-linolenic acid is capable of undergoing a series of desaturation and elongation reactions leading to its conversion to EPA (C20:5n-3) and DHA (C22:6n-3). However, due to the limited activity of Δ6 desaturase generating stearidonic acid (SDA; C18:4n-3), a rate-limiting enzyme for α-linolenic acid substrates, the conversion to EPA and DHA is poor (Li et al. 2017). Dietary SDA increased the red blood cell EPA content by approximately 17%, whereas the efficiency of α-linolenic acid was approximately 0.1% (Harris et al., 2008).
Conclusions
Diets supplemented with 4.5% rapeseed or mixed oil did not affect growth performance and carcass characteristics, but increased the IMF content in the LM, improved the fatty acid composition of the LM and SAT, and promoted the metabolic health of finishing pigs. However, the mixed oil diet with an n-6:n-3 PUFA ratio of 2:1 had multiple benefits for meat quality, including reduced drip loss and cooking loss, increased α-linolenic and linoleic acid content, and a decreased n-6:n-3 PUFA ratio. A mixed oil diet with an n-6:n-3 PUFA ratio of 2:1 is an effective strategy for improving meat quality and promoting the metabolic health of pigs.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31860633) and by the Special Project for Central Universities to Build World-Class Universities (disciplines) and Characteristic Development Guidance and by the Natural Science Foundation of Gansu Province (20JR5RA504).
Authors' contributions
SSJ and JXL wrote this manuscript. FW, CCT, and DLYEA conducted the major experiments. GHZ, SSJ, and LF performed parts of the experiments and analyses. JXL and GHZ reviewed and edited the manuscript.
Conflict of Interest Declaration
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Alfaia, C.M., Lopes, P.A., Madeira, M.S., Pestana, J.M., Coelho, D., Toldrá, F., & Prates, J.A.M., 2019. Current feeding strategies to improve pork intramuscular fat content and its nutritional quality. Adv. Food. Nutr. Res. 89, 53-94. doi: 10.1016/bs.afnr.2019.03.006. [ Links ]
Ba, H.V., Seo, H.W., Seong, P.N., Cho, S.H., Kang, S.M., Kim, Y.S., Moon, S.S., Choi, Y.M., & Kim, J.H., 2019. Live weights at slaughter significantly affect the meat quality and flavor components of pork meat. Anim. Sci. J. 90(5), 1-13. doi: 10.1111/asj.13187. [ Links ]
Boss, M., Kemmerer, M., Brüne, B., & Namgaladze, D., 2015. FABP4 inhibition suppresses PPARy activity and VLDL- induced foam cell formation in IL-4-polarized human macrophages. Atherosclerosis. 240(2), 424-430. doi: 10.1016/j.atherosclerosis.2015.03.042. [ Links ]
China, M.o.A.o.t.P.s.R.o., 2014. "Technical regulation for testing of carcass traits in lean-type pig". NY/T 825. [ Links ]
Corino, C., Rossi, R., Cannata, S., & Ratti, S., 2014. Effect of dietary linseed on the nutritional value and quality of pork and pork products: Systematic review and meta-analysis. Meat. Sci. 98(4), 679-688. doi: 10.1016/j.meatsci.2014.06.041. [ Links ]
Dannenberger, D., Nuernberg, K., Nuernberg, G., & Priepke, A., 2012. Different dietary protein and PUFA interventions alter the fatty acid concentrations, but not the meat quality, of porcine muscle. Nutrients. 4(9), 1237-1246. doi: 10.3390/nu4091237. [ Links ]
De Tonnac, A., Karim-Luisset, S., & Mourot, J., 2017. Effect of different dietary linseed sources on fatty acid composition in pig tissues. Livest. Sci. 203, 124-131. doi: 10.1016/j.livsci.2017.03.022. [ Links ]
Doran, O., Moule, S.K., Teye, G.A., Whittington, F.M., Hallett, K.G., & Wood, J.D., 2006. A reduced protein diet induces stearoyl-CoA desaturase protein expression in pig muscle but not in subcutaneous adipose tissue: Relationship with intramuscular lipid formation. Br. J. Nutr. 95(3), 609-617. doi: 10.1079/bjn20051526. [ Links ]
Duan, Y.H., Li, F., Li, L.N., Fan, J., Sun, X.M., & Yin, Y.N., 2014. n-6:n-3 PUFA ratio is involved in regulating lipid metabolism and inflammation in pigs. Br. J. Nutr. 111(3), 445-451. doi: 10.1017/S0007114513002584. [ Links ]
Fan, R., Kim, J., You, M., Giraud, D., Toney, A.M., Shin, S.H., Kim, S.Y., Borkowski, K., Newman, J.W., & Chung, S., 2020. α-Linolenic acid-enriched butter attenuated high fat diet-induced insulin resistance and inflammation by promoting bioconversion of n-3 PUFA and subsequent oxylipin formation. J. Nutr. Biochem. 76, 108285. doi:10.1016/j.jnutbio.2019.108285. [ Links ]
Folch J., Lees M., & Sloane Stanley G.H., 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Bio. Chem. 226(1), 497-509. [ Links ]
Friendship, R.M., Lumsden, J.H., McMillan, I., & Wilson, M.R., 1984. Hematology and biochemistry reference values for Ontario swine. Can. J. Comp. Med. 48(4), 390-393. [ Links ]
Gol, S., Gonzalez-Prendes, R., Bosch, L., Tor, M., Reixach, J., Pena, R.N., & Estany, J., 2019. Linoleic acid metabolic pathway allows for an efficient increase of intramuscular fat content in pigs. J. Anim. Sci. Biotechnol. 10, 33. doi:10.1186/s40104-019-0343-8. [ Links ]
Gregoire, F.M., Smas, C.M., and Sul, H.S., 1998. Understanding adipocyte differentiation. Physiol. Rev. 78(3), 783-809. doi: 10.1152/physrev.1998.78.3.783. [ Links ]
Gregory, M.K., Gibson, R.A., Cook-Johnson, R.J., Cleland, L.G., & James, M.J., 2011. Elongase reactions as control points in long-chain polyunsaturated fatty acid synthesis. PLoS One. 6(12), e29662. doi: 10.1371/journal.pone.0029662. [ Links ]
Hăbeanu, M., Lefter, N.A., Gheorghe, A., Nagy, A., Marin, D., & Ropotă, M., 2014. Effects of dietary flaxseed oil on the muscle fatty acid composition in Mangalitsa pigs in an extensive rearing system. S. Afr. J. Anim. Sci. 44(3), 240. doi: 10.4314/sajas.v44i3.5. [ Links ]
Hăbeanu, M., Lefter, N.A., Gheorghe, A., Untea, A., Ropotă, M., Grigore, D.M., Varzaru, I., & Toma, S.M., 2019. Evaluation of Performance, nitrogen metabolism and tissue composition in barrows fed an n-3 PUFA-rich diet. Animals (Basel). 9(5), 234. doi: 10.3390/ani9050234. [ Links ]
Harris, W.S., Lemke, S.L., Hansen, S.N., Goldstein, D.A., DiRienzo, M.A., Su, H., Nemeth, M.A., Taylor, M.L., Ahmed, G., & George, C., 2008. Stearidonic acid-enriched soybean oil increased the omega-3 index, an emerging cardiovascular risk marker. Lipids. 43(9), 805-811. doi: 10.1007/s11745-008-3215-0. [ Links ]
Hernández-Sánchez, J., Amills, M., Pena, R.N., Mercadé, A., Manunza, A., & Quintanilla, R., 2013. Genomic architecture of heritability and genetic correlations for intramuscular and back fat contents in Duroc pigs. J. Anim. Sci. 91(2), 623-632. doi: 10.2527/jas.2012-5270. [ Links ]
Hilgendorf, K.I., Johnson, C.T., Mezger, A., Rice, S.L., Norris, A.M., Demeter, J., Greenleaf, W.J., Reiter, J.F., Kopinke, D., & Jackson, P.K., 2019. Omega-3 fatty acids activate ciliary FFAR4 to control adipogenesis. Cell. 179(6), 1289-1305.e1221. doi: 10.1016/j.cell.2019.11.005. [ Links ]
Huang, C., Chiba, L.I., Magee, W.M., Wang, Y., Roding, S.P., Bratcher, C.L., Bergen, W.G., & Spangler,., 2020. Effect of flaxseed oil, poultry fat, and vitamin E supplementation on physical and organoleptic characteristics and fatty acid profile of pork, and expression of genes associated with lipid metabolism. Livest. Sci. 231 , 1-9. doi: 10.1016/j.livsci.2019.103849. [ Links ]
Huang, F.R., Zhan, Z.P., Luo, J., Liu, Z.X., & Peng, J., 2008. Duration of dietary linseed feeding affects the intramuscular fat, muscle mass, and fatty acid composition in pig muscle. Livest. Sci. 118(1-2), 132-139. doi: 10.1016/j.livsci.2008.01.002. [ Links ]
Iyer, M.N., Sarmah, B.C., Tamuli, M.K., Das, A., & Kalita, D., 2012. Effect of dietary sunflower oil and coconut oil on adipose tissue gene expression, fatty acid composition, and serum lipid profile of grower pigs. Arch. Anim. Nutr. 66(4), 271-282. doi: 10.1080/1745039x.2012.683324. [ Links ]
Leikus, R., Juskiene, V., Stankeviciene, Juska, R., Juodka, R., Stankeviciene, D., Nainiene, R., Juodka, R., & Siukscius, A., 2018. Effect of linseed oil sediment in the diet of pigs on the growth performance and fatty acid profile of meat. Rev. Bras. Zootecn. 47, e20170104. doi: 10.1590/rbz4720170104. [ Links ]
Li, F., Duan, Y., Li, Y., Tang, Y., Geng, M., Oladele, O.A., Kim, S.W., & Yin, Y., 2015. Effects of dietary n-6:n-3 PUFA ratio on fatty acid composition, free amino acid profile, and gene expression of transporters in finishing pigs. Br. J. Nutr. 113(5), 739-748. doi: 10.1017/S0007114514004346. [ Links ]
Li, Y., Rong, Y., Bao, L., Nie, B., Ren, G., Zheng, C., Amin, R., Arnold, R.D., Jeganathan, R.B., & Huggins, K.W., 2017. Suppression of adipocyte differentiation and lipid accumulation by stearidonic acid (SDA) in 3T3-L1 cells. Lipids. Health. Dis. 16(1), 181. doi: 10.1186/s12944-017-0574-7. [ Links ]
Martínez-Fernández, L., Laiglesia, L.M., Huerta, A.E., Martínez, J.A., & Moreno-Aliaga, M.J., 2015. Omega-3 fatty acids and adipose tissue function in obesity and metabolic syndrome. Prostag. Oth. Lipid. M. 121(Pt A), 24-41. doi: 10.1016/j.prostaglandins.2015.07.003. [ Links ]
Nguyen, D.H., Upadhaya, S.D., Lei, X.J., Yin, J., & Kim, I.H., 2019. Influence of dietary protease supplementation to corn-soybean meal-based high- and low-energy diets on growth performance, nutrient digestibility, blood profiles, and gas emission in growing pigs. Can. J. Anim. Sci. 99(3), 482-488. doi: 10.1139/cjas-2017-0104. [ Links ]
Nong, Q.Y., Wang, L.Y., Zhou, Y., Sun, Y.B., Zhou, Y.B., Sun, Y., Chen, W.T., Xie, J.T., Zhu, X.D., & Shan, T.Z., 2020. Low dietary n-6/n-3 PUFA ratio regulates meat quality, reduces triglyceride content, and improves fatty acid composition of meat in Heigai pigs. Animals (Basel). 10(9),1543. doi: 10.3390/ani10091543. [ Links ]
NRC (National Research Council), 2012. Nutrient Requirements of Swine. 11th ed. National Academy Press, Washington DC, USA. [ Links ]
Nuernberg, K., Fischer, K., Nuernberg, G., Kuechenmeister, U., Klosowska, D., Eliminowska-Wenda, G., Fiedler, I., & Ender, K., 2005. Effects of dietary olive and linseed oil on lipid composition, meat quality, sensory characteristics and muscle structure in pigs. Meat. Sci. 70(1), 63-74. doi: 10.1016/j.meatsci.2004.12.001. [ Links ]
Ogłuszka, M., Szostak, A., Te Pas, M.F.W., Poławska, E., Urbański, P., Blicharski, T., Pareek, C.S., Juszczuk-Kubiak, E., Dunkelberger, J.R., Horbańczuk, J.O., & Pierzchala, M., 2017. A porcine gluteus medius muscle genome-wide transcriptome analysis: Dietary effects of omega-6 and omega-3 fatty acids on biological mechanisms. Genes. Nutr. 12, 4. doi: 10.1186/s12263-017-0552-8. [ Links ]
Okrouhla, M., Stupka, R., Citek, J., Lebedova, N., & Zadinova, K., 2018. Effect of duration of dietary rapeseed and soybean oil feeding on physical characteristics, fatty acid profile, and oxidative stability of pig backfat. Animals. 8(11),193. doi: 10.3390/ani8110193. [ Links ]
Palmquist, D.L., 2009. Omega-3 fatty acids in metabolism, health, and nutrition and for modified animal product foods. Prof. Anim. Sci. 25(3), 207-249. doi: 10.15232/S1080-7446(15)30713-0. [ Links ]
Puig-Oliveras, A., Ramayo-Caldas, Y., Corominas, J., Estellé, J., Pérez-Montarelo, D., Hudson, N.J., Casellas, J., Folch, J.M., & Ballester, M., 2014. Differences in muscle transcriptome among pigs phenotypically extreme for fatty acid composition. PLoS One. 9(6), e99720. doi: 10.1371/journal.pone.0099720. [ Links ]
Realini, C.E., Duran-Montge, P., Lizardo, R., Gispert, M., Oliver, M.A., & Esteve-Garcia, E., 2010. Effect of source of dietary fat on pig performance, carcass characteristics, and carcass fat content, distribution and fatty acid composition. Meat. Sci. 85(4), 606-612. doi: 10.1016/j.meatsci.2010.03.011. [ Links ]
Skugor, A., Kjos, N.P., Sundaram, A.Y.M., Mydland, L.T., Ånestad, R., Tauson, A.-H., & 0verland, M., 2019. Effects of long-term feeding of rapeseed meal on skeletal muscle transcriptome, production efficiency and meat quality traits in Norwegian Landrace growing-finishing pigs. PLoS One. 14(8), 1-27. doi: 10.1371/journal.pone.0220441. [ Links ]
Song, C.H., Oh, S.M., Lee, S., Choi, Y., Kim, J.D., Jang, A., & Kim, J., 2020. The ratio of dietary n-3 polyunsaturated fatty acids influences the fat composition and lipogenic enzyme activity in adipose tissue of growing pigs. Food. Sci. Anim. Resour. 40(2), 242-253. doi: 10.5851/kosfa.2020.e8. [ Links ]
Tarricone, S., Colonna, M.A., Giannico, F., Ragni, M., Lestingi, A., & Facciolongo, A.M., 2020. Effect of an extruded linseed diet on meat quality traits in Nero Lucano pigs. S. Afr. J. Anim. Sci. 49(6), 1093-1103. doi: 10.4314/sajas.v49i6.13. [ Links ]
Vehovsky, K., Stupka, R., Zadinova, K., Sprysl, M., Okrouhla, M., Lebedova, N., Mlynekova, E., & Citek, J., 2019. Effect of dietary rapeseed and soybean oil on growth performance, carcass traits, and fatty acid composition of pigs. Rev. Bras. Zootecn. 48. doi: 10.1590/rbz4820180131. [ Links ]
Vitali, M., Dimauro, C., Sirri, R., Zappaterra, M., Zambonelli, P., Manca, E., Sami, D., Fiego, D.P.L., & DavoliId, R., 2018. Effect of dietary polyunsaturated fatty acid and antioxidant supplementation on the transcriptional level of genes involved in lipid and energy metabolism in swine. PLoS One. 13(10). doi: 10.1371/journal.pone.0204869. [ Links ]
Wang, H., & Eckel, R.H., 2009. Lipoprotein lipase: From gene to obesity. Am. J. Physiol. Endocrinol. Metab. 297(2), E271-288. doi: 10.1152/ajpendo.90920.2008. [ Links ]
Wang, Y., Viscarra, J., Kim, S.J., & Sul, H.S., 2015. Transcriptional regulation of hepatic lipogenesis. Nat. Rev. Mol. Cell Biol. 16(11), 678-689. doi: 10.1038/nrm4074. [ Links ]
Watt, M.J., Holmes, A.G., Pinnamaneni, S.K., Garnham, A.P., Steinberg, G.R., Kemp, B.E., & Febbraio, M.A., 2006. Regulation of HSL serine phosphorylation in skeletal muscle and adipose tissue. Am. J. Physiol. Endocrinol. Metab. 290(3), E500-508. doi: 10.1152/ajpendo.00361.2005. [ Links ]
Xu X., Chen X., Chen D., Yu B., Yin J., & Huang Z., 2019. Effects of dietary apple polyphenol supplementation on carcass traits, meat quality, muscle amino acid, and fatty acid composition in finishing pigs. Food. Funct. 10(11 )7426- 7434. doi: 10.1039/c9fo01304k. [ Links ]
Yu, C.H., Xi, L.L., Chen, J., Jiang, Q., Yi, H.B., Wang, Y.Z., & Wang, X.X., 2017. PAM, OLA, and LNA are differentially taken up and trafficked via different metabolic pathways in porcine adipocytes. Lipids. 52(11), 929-938. doi: 10.1007/s11745-017-4302-x. [ Links ]
Submitted 4 April 2022
Accepted 28 September 2022
Published 27 February 2023
# Corresponding author: JXL, lu2003jx@163.com; GHZ, 381284797@qq.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}