










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 no.5 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i5.08
Influence of feeding on different types of pasture on the fatty acid profile and health indices of goat milk
D. MierlitaI, II, #
IUniversity of Oradea, Department of Animal Science, Oradea City, 410087, Romania
IIUniversity of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, 400372, Romania
ABSTRACT
The main objective of the present research was to study the change in fatty acid (FA) composition of milk fat in a Carpathian local breed during grazing on two plots characterized by a different cover with herbaceous species and woody (only herbaceous species: H and high woody species cover: W) allowances with or without concentrate supplementation (Cs: 500 g concentrate/head/d). Thus, four treatments resulted: H-U: only herbaceous species-unsupplemented, H-Cs: only herbaceous species-concentrate supplementation, W-U: high woody species-unsupplemented, W-Cs: high woody species-concentrate supplementation. Individual milk production (n = 10) was recorded daily, and milk samples were collected weekly for a seven-week period (June to August) and analysed for fat, protein, lactose, urea-N, and FAs. Milk production was not influenced by the type of pasture. The daily production of milk, 3.5% fat-corrected milk, and protein level increased and the content of fat and urea-N reduced with concentrate supplementation, whereas the content of lactose was similar. The botanical composition of pasture markedly influenced the FA profile of goat milk fat. The goats grazing in scrubland (W group) produced milk fat with higher concentrations of monounsaturated FA (C18:1 t11, vaccenic acid, VA) and polyunsaturated FA (omega 3 and conjugated linoleic acid, CLA) and lower concentrations of hypercholesterolaemic FA (HFA; C12:0 + C14:0 + C16:0). Regardless of the botanical composition of the pasture, milk fat of unsupplemented grazed goats had the lowest proportion of HFA, as well as the highest proportions of C18:1 c9, VA, RA (rumenic acid, C18:2 c9,t11), and total n-3 FA compared to the Cs groups (supplemented with concentrate). Concentrate supplementation increased short-chain FA and saturated FA content. In contrast, percentages of the nutritionally desirable FA (C18:3 c9, c12, c15; C18:2 c9, f 11, C18:1 f 11 ; C20:5n-3; C22:5n-3, and total n-3 FA) were substantially lower, whereas levels of total n-6 FA, n-6:n-3 ratio, atherogenicity index (AI), and thrombogenicity index (TI) were substantially higher in milk from Cs groups. No interactions occurred between pasture type and concentrate supplementation for milk FA composition. It is concluded that the milk from goats kept unsupplemented on the natural pasture were better in terms of nutraceutical parameters advantageous to human nutrition than the milk produced by concentrate supplementation.
Keywords: indigenous goats, milk fatty acids, conjugated linoleic acid, pasture type, shrublands
Introduction
Goat milk and dairy products are considered good sources of high-quality nutrients, particularly proteins and fats. The research on milk fat is oriented to the improvement of its nutritional value, with particular attention paid to increasing the content of FAs considered beneficial to human health, especially rumenic acid [RA; C18:2 c9, t11; also named c9, t11-conjugated linoleic acid (c9,t11CLA)], vaccenic acid (VA; C18:1 t11), and α-linolenic acid (ALA; C18:3 c9, c12, c15), and to enhancing the n-6:n-3 ratio, considering that a more balanced dietary intake of n-6 relative to n-3 is desired for optimal human health (Nuda et al., 2020).
Numerous studies have shown that modification of the basal diet of animals, especially by lipid supplementation, is the most appropriate procedure for changing the fatty acid profile in goats milk (Chilliard et al., 2003). However, the use of these supplements can increase feeding costs. Forages, even though containing a relatively low level of lipids, are often the major source of beneficial unsaturated fatty acids in ruminant diets. Fresh grass contains low levels of lipid extract but 45-75% of C18:3 c9, c12, c15 (Cabiddu et al., 2005). In literature, it has been recognised that favourable changes in milk FA profile can be obtained by grazing or feeding fresh forages. Results of many studies indicate that milk produced from grassland, particularly from botanically diverse pastures, has higher concentrations of fatty acids which are considered to be a benefit to human health (Chilliard et al., 2007; Moloney et al., 2008). Feeding fresh grass (rich in α-linolenic acid) compared to concentrates (rich in linoleic acid) results in higher concentrations of omega-3 FA and CLA in goat milk (Moloney et al., 2008). Linoleic and linolenic acid in feed are the precursors of CLA in milk.
The goats are capable of utilizing natural pastures that are often dominated by shrubs, selecting foliage from the upper vegetation layer (Fedele et al., 2002). Furthermore, goats can consume tannin-rich resources and digest them more efficiently than other domestic animals (Decandia et al., 2007a). Goat grazing on scrub vegetation can be used as an effective tool for the control of shrubs in protected forest areas, without losing biodiversity (Mancilla-Leytón et al., 2014).
Although previous studies have assessed the impact of pastures with different botanical composition on milk production and composition, there are still only a limited number of studies on how grazing of pastures high in shrublands affect the fat composition of the milk of dairy goats (Decandia et al., 2007b). In addition, the influence of dominant species in the botanical composition of the pastures high in shrublands in the submountain geographical areas (e.g., Rubinia pseudoacacia L., Carpenus betuíus L., Fagus syívatica L.) have been less studied.
The natural pastures don't always support adequate production in goats, and in this case, the use of supplements in the form of concentrates is necessary. The supplementation of the diet with concentrates can influence the FA composition of milk, productive performance, eating behaviour, and ruminal activity of grazing goats (Bonanno et al., 2008).
In this context, the main objective of the present study was to study the change in FA composition of milk fat from dairy goats during grazing on two plots characterized by a different cover with herbaceous species and woody (only herbaceous species: H and high woody species cover: W) allowances with or without concentrate supplementation (i.e., 0 versus 500 g concentrate/goat/day).
Materials and Methods
The research was carried in the submountain area of western Carpathian Mountains (northwestern Romania, 46037'42.6" N 22023'34.1" E, 960 m above sea level, total annual rainfall 718 mm; mean annual temperature 9.1 °C). All the experimental procedures were approved by the Committee of Bioethics in Scientific Research of the University of Oradea (Romania) and were in accordance with the EU Directive 2010/63/EU on the protection of animals used for scientific purposes. Goat breeding in Romania is based on native breeds, which are well adapted to submountain and mountainous regions with large areas of pastures, and prevalent breed is Carpathian (over 80% of the goats in Romania belong to this breed). Goats of this breed produce 200-250 kg of milk, with the average fat content ranging between 2.7 and 4%, with 3-4% protein.
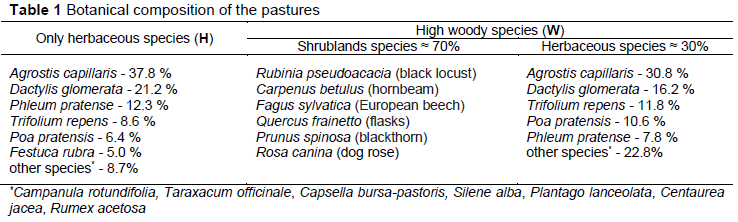
A representative flock of 360 Carpathian, local breed, adult female goats was divided into two equal herds and randomly allocated to two plots characterised by a different cover with herbaceous species and woody species (only herbaceous species - H and high woody species cover - W). The most common herbaceous species in the H pasture (without wood species) (approximately 12.0 ha) were Agrostis capiííaris, Dactyíis gíomerata, and Trifoíium repens. The rangeland (W) was approximately 14.0 ha, with the following vegetation types (% of land area): shrubland species (70%) and herbaceous species (30%). Dominating shrublands species were Rubinia pseudoacacia, Carpenus betuíus, and Fagus syívatica and most common herbaceous were A. capiííaris, D. gíomerata, and T. repens.
Using twenty adult goats from each herd, selected by age (2-3 years), productive stage (13 ± 2 w of lactation) and similar body characteristics (38.2 ± 2.9 kg at the beginning of the experiment), milk yield was determined and milk was sampled for composition analyses. Ten goats from each herd received a supplement of 500 g concentrate/head/d. The concentrate was fed twice daily during milking. Thus, four treatments resulted from the combination of two pasture types (H: only herbaceous species and W: high woody species) and two concentrate supplementation levels (i.e., 0 versus 500 g concentrate/goat/day): H-U: only herbaceous species-unsupplemented, H-Cs: only herbaceous species-concentrate supplementation, W-U: high woody species-unsupplemented, W-Cs: high woody species-concentrate supplementation.
The experiment was conducted for ten weeks, from early June to mid-August, 2019. The first three weeks were used as a covariate period (week 1) and adaptation to dietary treatments (weeks 2 and 3).
The grazing system was traditionally extensive and shepherd-led and the grazing route of the flock was planned in consideration of forage availability. The goats returned to their stalls after grazing and were housed at night.
Botanical composition was determined on forage samples taken randomly by quadratic frame (0.25 χ 0.25 m2) by manual separation of plant species. Botanical composition was calculated by dividing individual species weight by the total weight collected (wet basis) (Mierlitã et al., 2017).
Data on herbages and concentrates offered to the animals were recorded each month (n = 3). Hand plucked forage samples similar to those consumed by the goats were collected for chemical analysis (Decandia et al., 2007a). These samples were freeze-dried using Thermovac TM-20 (Froilabo, Ozoir-La-Ferrière, France) and analysed for crude protein (CP) and ether extract (EE) according to Association of Official Agricultural Chemists (AOAC, 1996), and for neutral detergent fiber (NDF) according to Van Soest et al. (1991). The results were referred on a dry matter basis (DM). The FA composition (after total lipid extraction and methylation, as described by Sukhija & Palmquist, 1988) of these samples were also measured using the same methods as for the milk.
Milk yield of individual goats was recorded daily (from two subsequent milkings). The goats were milked manually twice daily at 08:00 and 16:00 h. Samples of milk (100 ml) were collected from each goat from two subsequent milkings. Each sample of individual milk was obtained weekly by pooling 60% of the morning milk with 40% of the evening milk, based on the morning and evening milk yield. The composite sample for each goat was divided into two sets of milk sub-samples. A tablet of Bronopol (2-bromo-2-nitropropane-1,3-diol; D&F Inc., USA) was added to the first set of sub-samples (50 ml), which were analysed for fat, protein, lactose, and urea content with an infrared milk analyser (Milk Analyser System 4000, Foss Electric, Hillerod, Denmark). A second set of milk sub-samples (50 ml) was stored at -20 °C for the extraction of fat and further analysis of fatty acid composition.
To determine the composition of FAs in milk, the fat was extracted according to the international standard, ISO 14156/IDF 172:2001. Fatty acid methyl esters (FAME) were prepared according to the method proposed by Christie (1989). FAME were determined by gas chromatography using a Varian GC 3600 equipped with FID and a fused silica capillary column (SP 2560 Supelco); 100 m χ 0.25 mm id; film thickness, 0.20 μητ Helium was used as the carrier gas at a flow rate of 1 mL/min. The split ratio was 1:100. The oven temperature was programmed at 90 °C and held for 1.50 min, then increased to 210 °C at a rate of 9 °C/min, held at this temperature for 25 min, then increased to 230 °C at 15 °C/min, and held for 7 min. The temperatures of the injector and the detector were set at 270 °C (Mierlitã, 2016). Peaks were identified on the basis of the retention times of standard methyl esters of individual fatty acids (Sigma, St. Louis, MO). The proportions of the individual acids were calculated by the ratio of their peak area to the total area of all observed fatty acids.
The AI (atherogenic index), TI (thrombogenic index), h/H (hypocholesterolaemic/ hypercholesterolemic), and HPI (health promoting index) were calculated from the FA profile using the following equations:

where: C12:0, C14:0, C16:0, and C18:0 are lauric, myristic, palmitic, and stearic saturated FAs; C18:1, C18:2, C18:3 C20:4, C20:5, and C22:5 are oleic, linoleic, α-linolenic, arachidic, eicosapentaenoic (EPA), and docosapentaenoic (DPA) unsaturated FAs; n3 FAs acids are α-linolenic acid, EPA and DPA; and n6 FAs include linoleic, linolelaidic and arachidonic acids
Data obtained were analysed as a 2 χ 2 factorial randomized block design using the PROC MIXED model of SAS (2001). Fixed effects were the pasture type, concentrate supplementation, and interactions between fixed effects, the time, and an appropriate covariate. Random effects of goats were used as the error term. Variance-covariance structure was first autoregressive [AR(1)] (autoregressive model of order 1). For FA proportions in milk fat, analysis was performed without a covariate. Overall differences between treatment means and interactions for pasture type and concentrate supplementation were considered to be significant at P <0.05.
Results and Discussion
The botanical composition of the pastures is shown in Table 1. The most common species in the H pasture (approximately 12.0 ha) in terms of occurrence were (% of total weight collected, wet basis): A. capillaris (37.8), D. glomerata (21.2), T. repens (8.6), Phleum pratense (12.3), Poa pratensis (6.4), Festuca rubra (5.0), and other grasses (8.7). The rangeland (W) was approximately 14.0 ha, with the following vegetation types (% of land area): shrubland species (70) and herbaceous species (30) (Table 1). Dominating species were black locust (R. pseudoacacia L.), hornbeam (C. betulus L.), European beech (F. sylvatica L.) and common bent (A. capillaris L.). The concentrate contained the following ingredients (g/kg): maize (800), sunflower meal (150), and minerals and vitamins (50).
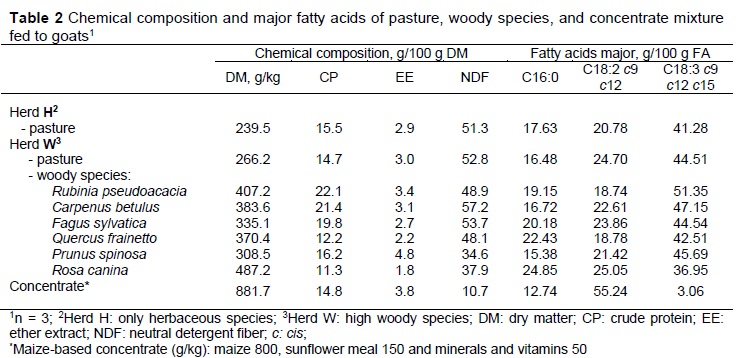
The chemical and FA compositions of the pasture, woody species and concentrates are given in Table 2. Pasture was of medium quality, averaging (DM basis) 14.7-15.5% CP and 51.3-52.8% NDF. In herd H, the pasture presented a higher CP content and lower NDF content than for herd W. Crude protein was higher when goat's diets included shrub rangeland, particularly R. pseudoacacia, C. betulus, and F. sylvatica.
The α-linolenic (C18:3, 3.0-51.3 g/100 g FA), linoleic (C18:2, 18.7-55.2 g/100 g FA), and palmitic (C16:0, 12.7-24.8 g/100 g FA) acids were the most abundant FAs in all forages used in goat feed, together representing over 70% of the fatty acids present. As expected, α-linolenic acid (ALA) was the most dominant fatty acid for pasture and woody species. The content of ALA in the pasture (41.28-44.51 g/100 g FA), was in accordance with previously reported values for similar pasture species in the study of Corazzin et al. (2019). The highest values of ALA were found in woody species, especially in R. pseudoacacia (black locust) and C. betulus (hornbeam) (51.35 and 47.15%, respectively). The lipids of forage are located in the leaf chloroplasts; a high proportion of leaves in the woody species might therefore lead to a high intake of ALA (Elgersma et al., 2005). The content of C18:3 in the woody species was higher than reported by Decandia et al. (2007b). Differences in C18:3 among studies could be due to differences in woody composition species, management, sampling, and season. The most representative FA in concentrate was linoleic acid (LA), similar to the fatty acid composition of maize grain (Rego et al., 2016).
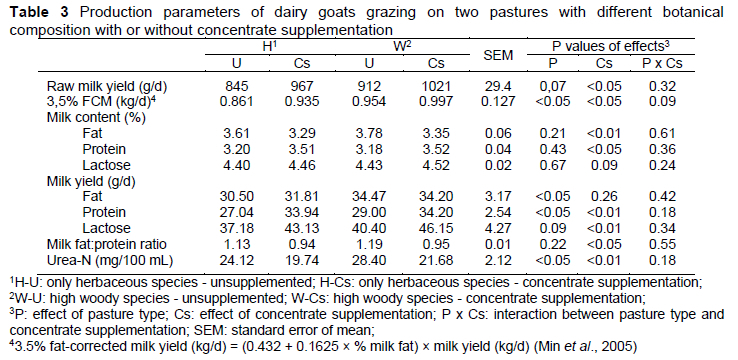
Raw milk production and milk composition was not influenced by the type of pasture. Similar results were reported by Decandia et al. (2007a) when goats were fed on natural pastures with different botanical composition: woody and herbaceous species vs. only herbaceous species. Even though woody species have a lower nutritional value than herbaceous species due to the higher content of NDF and ADF, milk production is not affected because the presence of woody vegetation causes an increase in goat intake from 420-910 g/day (only herbaceous vegetation) to 550-1250 g/day of herbaceous and woody vegetation (Fedele et al., 2002). In particular, the milk urea level was high in the W-group goats, as a consequence of a higher CP level of the woody species than herbaceous vegetation (Table 2), an aspect supported by studies previously conducted by Decandia et al. (2007a).
The daily production of milk, 3.5% fat-corrected milk, and protein level increased (p <0.05) and the levels of fat and urea reduced with concentrate supplementation (p <0.01), whereas the content of lactose was similar (Table 3). In experiments on natural pastures (Rubino et al., 1995) the effects of a high supply of concentrates on milk production were absent or almost negligible, because herbage intake decreased as concentrate supplementation increased, whereas in the experiments of Claps et al. (1994), the highest milk production was reached with the highest concentrate intake (1050 g/day). Nevertheless, a lower intake of structural carbohydrates from the pasture induced by the supply of concentrates decreased milk fat concentration (Bonanno et al., 2004), probably due to a lower rumen production of acetic acid, the precursor of short- and long-chain fatty acids in milk.
An inverse relationship was observed between concentrate supplementation and fat content in the milk (Table 3), which can be related to the reduction of precursors for the synthesis of fat in the mammary gland (Bauman & Griinari, 2003). According to these authors, the increase in the production of propionic acid in diets with a high content of grains promotes a glycaemic increase through gluconeogenesis, which would result in a greater secretion of insulin. As a consequence, there would be a decrease in lipolysis, reducing the contribution of preformed fatty acids for fat synthesis in the mammary gland (Rufino et al., 2012). Our results are in contrast to those in the literature (Min et al., 2005), which report increased milk fat percentages when cereal-based concentrates are provided to grazing, dairy Alpine goats. One possible explanation for the high milk fat percentage is the highly digestible fibre in good-quality pasture (Min et al., 2005).
Milk protein content was increased when goats received concentrate supplementation. These results suggest that the CP ingested with pasture in unsupplemented diets (H-U and W-U groups) was inefficiently used due to the higher content of rumen degradable N in pasture in relation to its fermentable energy content and that some amino acids might have been used as glucose precursors (due to the low dietary energy), rather than in milk protein synthesis (Bargo et al., 2003). This hypothesis is supported by the increased content of milk urea in goats without concentrate (especially in goats fed on pasture with high woody species cover).
In the present experiment, milk urea-N was higher in treatment W-U compared with treatment H-U, as a consequence of a higher CP level of the shrubland species. Average urea-N milk concentrations decreased for treatment Cs, suggesting that concentrate supplementation altered rumen metabolites (Min et al., 2005). Milk urea-N can be used as a tool to monitor protein feeding efficiency, dietary protein:energy ratio, and animal health status.
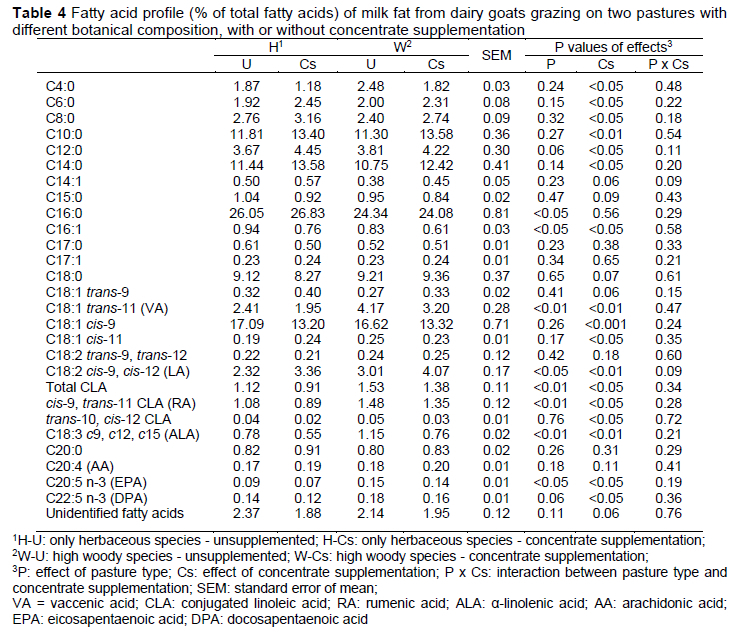
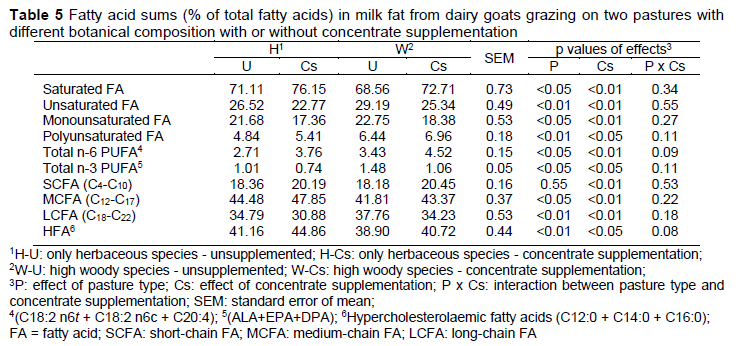
Feeding strategies have been shown to affect milk FA composition of dairy goats fed on pasture. The percentages of the nutritionally desirable FA (α-linolenic acid, total n-3 PUFA, total conjugated linoleic acid and its isomers, and vaccenic acid) were substantially higher in milk from the W group, whereas the C16:0, total saturated FA, and n6:n3 ratio were substantially lower than in the H group (Tables 4 and 5). The results obtained in the current study are inconsistent with Decandia et al. (2007a), who found a decrease in the content of medium-chain FA (primarily due to a decrease in C16:0) when goats grazed on woody and herbaceous species compared to those that grazed only herbaceous species. Chilliard et al. (2006) reported that oleic acid was the most prominent mono-unsaturated fatty acid, with concentrations of 15-21% of total FA, which is similar to that found in the current study. In a study conducted in Australia, the most abundant saturated fatty acid was C18:0 (Thomas & Rowney, 1996), not C16:0, as in this study. This difference may be due to goats eating pastures with different botanical composition in the rangeland. In Australia, the animals grazed on sea grass, thus the presence of C18:0 could be due to biohydrogenation of α-linolenic acid (Chilliard et al., 2003).
Higher proportion of C18:1 t11 and c9, t11 CLA in milk produced on W than on H could be explained by higher intake of the precursors, C18:2 c9, c12 and C18:3 c9, c12, c15, but also due to plant secondary metabolites, like tannins, present in grazed woodland species, which decrease the ruminal biohydrogenation of linolenic acid by ~30%, resulting in an increase in linolenic acid content in milk (Cabiddu et al., 2009). Grazing condensed tannin-containing herbage has been shown to increase milk proportions of C18:1 t11, c9, t11 CLA, C18:2 c9, c12, and C18:3 c9, c12, c15 in ewes (Cabiddu et al., 2009). Similarly, the milk proportions of C18:2 c9, c12 and C18:2 c9, c12, c15 were higher on W than on H in the present study (Table 4). However, if this effect was due to differences in intake of condensed tannins, further studies should be done.
Concentrate supplementation decreased the nutritionally desirable FAs such as CLA, VA, total n-3 FA, ALA, eicosapentaenoic acid (EPA; C20:5n-3), docosapentaenoic acid (DPA; C22:5n-3) and increased the n-6/n-3 ratio; these feeds increased SFA, PUFA n-6, and LA of goat milk (Table 4 and 5). Consistent with our results, concentrate supplementation reduced the milk fat content of VA and ALA in grazing dairy goats (LeDoux et al., 2002). The concentrate supplementation probably reduced the intake of grass and it subsequently resulted in a reduction in the intake of ALA, explaining not only the lower concentration of this FA in milk but also the reduction in VA, which is the common intermediate in ruminal biohydrogenation of LA and ALA (Nuda et al., 2020). The increased content of short- and medium-chain saturated FA in our study supports previous grazing studies that have evaluated the effects of concentrate supplementation on milk FA. Bargo et al. (2006) reported an increase in C6:0-C10:0 and C12:0-C17:0 content when grazing cows were supplemented with 1 kg concentrate/4 kg milk compared with unsupplemented cows. This increase was attributed to increased availability of ruminal volatile fatty acids for endogenous synthesis of these FA, due to higher energy intake with concentrate supplementation.
The proportions of oleic acid in goat milk decrease with an increasing percentage of concentrate in the diet (Mele et al., 2008). According to this, we detected the lowest values for oleic acid in samples of the Cs group, in which goats were fed supplemented with concentrates (500 g/d) (Table 4).
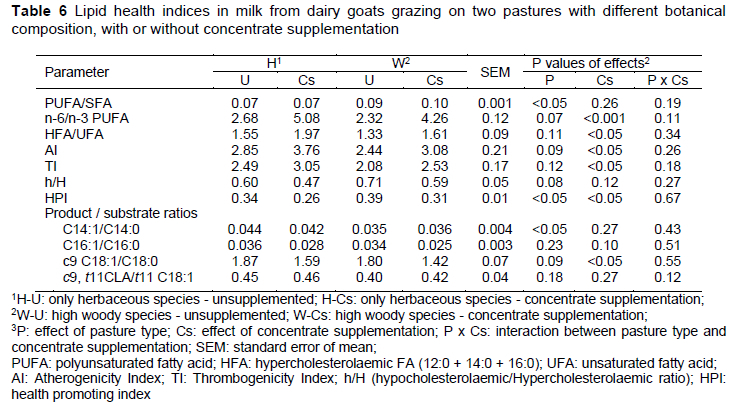
The lower content of c9, t11 CLA (by 10.0-18.7%) in milk fat in concentrate-supplemented goats supports the results from previous grazing studies (Elgersma, 2015). Bargo et al. (2006) suggested that the reduction in c9, t11 CLA (rumenic acid, RA) content in milk fat due to concentrate supplementation was primarily because of a reduction in the contribution of pasture to the diets. The higher RA content in milk of unsupplemented goats could be related to higher pasture DM intake and subsequently higher intake of C18:3 due to the high C18:3 content of pasture (Table 2). Biohydrogenation of dietary C18:3 yields, among others, C18:1 t11. The content of RA in milk fat depends on ruminal production of both c9 t11 CLA and C18:1 t11, and tissue activity of Δ9-desaturase (Bauman et al., 2001). Product to substrate ratios of some FAs represent a desaturase index and can serve as a proxy for Δ9-desaturase activity (Bargo et al., 2006). In our study, the ratio c9, t11 CLA/t11 C18:1 was not affected by treatment (Table 6), suggesting a similar ΔA9-desaturase activity in the mammary gland. The higher RA content in the unsupplemented goats would therefore not have been a result of differential enzyme activity, but of a higher amount of RA precursors, as indicated by the higher amount of t11 C18:1.
In our research, the higher amount of PUFA in Cs milk is mainly due to the higher amount of n-6 FA in these samples. The concentration of linoleic acid, the most representative FA of the PUFA, was higher in Cs milk (W-Cs and H-Cs groups) compared to U milk (W-U and H-U groups). These results agree with values reported in literature, specifically in the absence of lipids added to diets, as the proportion of LA in goat milk FAs is between 2% and 4% (Chilliard et al., 2007). Generally, all or most of LA in milk fat comes from dietary LA that escapes rumen biohydrogenation activity, and its transfer to milk is related to the amount of this fatty acid that is ingested. The composition of concentrate mixture offered to Cs groups appears to explain the highest content of LA found in milk fat.
It is interesting to note that the C18:0 level, a final product of the ruminal biohydrogenation of dietary FA, was not affected by dietary treatments. Dewanckele et al. (2018) explained that diets rich in PUFA can inhibit the last step of rumen biohydrogenation by modifying the rumen bacterial population, resulting in increased intermediate products, as observed in the present study. Since C18:1 c9 can be derived from the desaturation of C18:0, the difference observed between experimental groups in this FA could be due to the desaturation difference of C18:0. In fact, the desaturation ratio 18:1 c9/18:0 in the mammary gland was the lowest for samples from the Cs groups (p <0.05), in which goats' diet was supplemented with concentrates, since the increase in the availability of PUFA inhibits the Δ9-desaturase (Mele et al., 2008) (Table 6).
Grazing of woody species increased the proportion of long-chain n-3 PUFA, such as EPA, probably because of the high content of its precursor (ALA) in the pasture. Concentrate supplementation decreased the content of EPA and DPA in milk fat. The PUFA/SFA and n-6/n-3 PUFA ratios are commonly used to measure the nutritional value and consumer health of animal fat (Pilarczyk et al., 2015). Generally, a ratio of PUFA to SFA above 0.45 and a ratio of n-6/n-3 below 4.0 are required in the diet to combat some diseases such as coronary heart disease and cancer (Simopoulos, 2002). In the present study, the PUFA/SFA ratios (0.07-0.10) were considerably lower than the recommended values, whereas the n-6/n-3 ratios for unsupplemented goats (2.32 and 2.68, respectively) were within the recommended values (Table 6). The concentrate supplement had a negative effect on this ratio, due to the increase in LA content and the decrease in ALA content in milk fat. Values obtained in this study for the Cs groups are comparable to the values reported in literature for goat milk (5.0) (Lopez et al., 2019).
There was no significant difference in atherogenicity index (AI) and trombogenicity index (TI) of H and W goat groups. The usage of concentrates raised the AI and TI values, diminishing the beneficial effects of FA regarding consumer health. According to the findings by Tsiplakou et al. (2006) and Renna et al. (2012), milk from grazing goats had better quality parameters for human nutrition when compared with those fed with diets based on preserved forages and concentrates. Our results are similar to those reported by Chilliard et al. (2003) for goat milk, which produced similar values according to feeding.
The HPI (health promoting index) was proposed by Chen et al. (2004) as an indicator of the health value of dietary fat and is largely focused on the effect of some FA on cardiovascular diseases. Milk with a high HPI value is assumed to be more beneficial to human health. In the current study, the highest HPI value of goat milk was detected in the W-U group (0.39), followed by the H-U group (0.34) (p <0.05).
Supplementing pastures with concentrates had a negative effect on the HPI value, regardless of the type of pasture (p <0.05).
Conclusions
The feeding system based on pasture (only herbaceous species and high woody species) confirmed its positive effect on beneficial fatty acid content in milk, in particular when the pasture is characterised by a high proportion of shrubland species. In fact, when goats are fed pastures rich in shrubs, milk fat presented consistently higher concentrations of unsaturated FA including 18:1 t11; c9, t11 CLA; 18:3n-3; and 20:5n-3 and decreased the concentrations of hypercholesterolaemic FA (sum of 12:0, 14:0, and 16:0). Concentrate supplementation decreased the level of FAs beneficial for human health, such as C18:3n-3, and C20:5n-3, as well as some FA intermediates of rumen biohydrogenation (c9, t11 CLA and t11 C18:1). The use of pastures with different botanical composition used in the geographical study area in goats diets could serve as a strategy to improve milk FA profiles with beneficial effects on health in humans, having a positive effect on consumer perception and the acceptance of milk and dairy products from the local region.
The high heterogeneity of goat diets while feeding on shrublands makes it very difficult to understand their effect on milk FA composition. Further research is needed to investigate the relationship between dietary composition and milk content of beneficial FA.
Acknowledgements
The author gratefully acknowledges Rosbro Avicom SRL for providing the experimental facilities. The author thanks Prof. Dr Daraban Stelian for his help in collecting samples and the statistical analyses of results.
Authors' contributions
DM conceived the research, interpreted the data, and drafted the manuscript.
Conflict of Interest Declaration
The author declares having no conflict of interest relative to this work.
References
AOAC, 1996. Methods of Analysis. Association of Official Analytical Chemists, 16th Edition. [ Links ]
Bargo, F., Delahoy, J.E., Schroeder, G.F., Baumgard, L.H. & Muller, L.D., 2006. Supplementing total mixed rations with pasture increase the content of conjugated linoleic acid in milk. Anim. Feed Sci. Technol. 131, 226-240. [ Links ]
Bargo, F., Muller, L.D., Kolver, E.S. & Delahoy, J.E., 2003. Invited review: Production and digestion of supplemented dairy cows on pasture. J. Dairy Sci. 86, 1-42. [ Links ],
Bauman, D.E. & Griinari, J.M., 2003. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 23, 203-227. [ Links ]
Bauman, D.E., Corl, B.A., Baumgard, L.H. & Griinari, J.M., 2001. Conjugated linoleic acid (CLA) and the dairy cow. In: Garnsworthy, P.C.,Wiseman, J. (Eds.), Recent Advances in Animal Nutrition. Nottingham University Press, Nottingham, UK. [ Links ]
Bonanno, A., Di Grigoli, A., Alicata, M.L., Avondo, M., Pagano, R., De Vita, G., Stringi, L. & Giambalvo, D., 2004. Ingestione al pascolo e produzione di latte di capre Girgentane in funzione della combinazione spaziale delle specie foraggere e dell'integrazione conconcentrato. In: Proceeding of the 16th SIPAOC Congress, Siena, Italy, 241 (Abstract). [ Links ]
Bonanno, A., Fedele, V. & Grigoli, D., 2008. Grazing management of dairy goats on Mediterranean herbaceous pastures. Dairy Goats Feeding and Nutrition, eds A. Cannas and G. Pulina, CAB International, 189-220. [ Links ]
Cabiddu, A., Molle, G., Decandia, M., Spada, S., Fiori, M., Piredda, G. & Addis, M., 2009. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep: Part 2: Effects on milk fatty acid profile. Livest. Scie. 123(2), 230-240. DOI: 10.1016/j.livsci.2008.11.019. [ Links ]
Cabiddu, A., Decandia, M., Addis, M., Piredda, G., Pirisi, A. & Molle, G., 2005. Managing Mediterranean pastures in order to enhance the level of beneficial fatty acids in sheep milk. Small Rumin. Res. 59, 169-180. [ Links ]
Chen, S., Bobe, G., Zimmerman, S., Hammond, E.G., Luhman, C.M., Boylston, T.D., Freeman, A.E. & Beitz, D.C. 2004. Physical and sensory properties of dairy products from cows with various milk fatty acid compositions. J. Agric. Food Chem. 52, 3422-3428. [ Links ]
Chilliard, Y., Glasser, F., Ferlay, A., Bernard, L., Rouel, J. & Doreau, M., 2007. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur J Lipid Sci Technol. 109(8), 828-855. DOI: 10.1002/ejlt.200700080. [ Links ]
Chilliard, Y., Rouel, J. & Leroux, C., 2006. Goat's a-s1 casein genotype influences its milk fatty acid composition and D-9 desaturation ratios. Anim Feed Sci Tech 131, 474-487. [ Links ]
Chilliard, Y., Ferlay, A., Rouel, J. & Lambere, G., 2003. A review of nutritional and physiological factors affecting goat milk synthesis and lipolysis. J. Dairy Sci. 86, 1751-1770. [ Links ]
Christie, W.W., 1989. Gas chromatography and lipids - a practical guide; The Oily Press: Ayr, Scotland, UK, 307. [ Links ]
Claps, S., Rubino, R., Cogliandro, E. & Fedele, V., 1994. Fabbisogni alimentari e nutritivi, comportamento alimentare e produzione di latte di capre allevate al pascolo con differenti integrazioni alimentari. In: Rubino, R. (ed.) Atti Giornata Scientifica Miglioramento dell'efficienza produttiva degli ovini e dei caprini. Ars Grafica, Villa D'Agri, Italy, 1-16. [ Links ]
Corazzin, M., Romanzin, A., Sepulcri, A., Pinosa, M., Piasentier, E. & Bovolenta, S., 2019. Fatty acid profiles of cow's milk and cheese as affected by mountain pasture type and concentrate supplementation. Animals, 9, 68-81; doi:10.3390/ani9020068. [ Links ]
Decandia, M., Cabiddu, A., Molle, G., Branca, A., Epifani, G., Pintus, S., Tavera, F., Piredda, G., Pinna, G. & Addis, M., 2007a. Effect of different feeding systems on fatty acid composition and volatile compound content in goat milk. Options Méditerranéennes, 74, 129-134. [ Links ]
Decandia, M., Yiakoulaki, M., Pinna, G., Cabiddu, A. & Molle, G., 2007b. Foraging behaviour and intake of goats browsing on Mediterranean shrublands. CAB International, Dairy Goats Feeding and Nutrition (eds A. Cannas and G. Pulina), 161-188. [ Links ]
Dewanckele, L., Vlaeminck, B., Hernandez-Sanabria, E., Ruiz-González, A., Debruyne, S., Jeyanathan, J. & Fievez, V., 2018. Rumen biohydrogenation and microbial community changes upon early life supplementation of 22:6n-3 enriched microalgae to goats. Front. Microbiol. 9, 573. [ Links ]
Elgersma, A., Maudet, P., Witkowska, I.M. & Wever, A.C., 2005. Effects of nitrogen fertilisation and regrowth period on fatty acid concentrations in perennial ryegrass. Ann App Biol 147, 145-152. [ Links ]
Elgersma, A., 2015. Grazing increases the unsaturated fatty acid concentration of milk from grass-fed cows: A review of the contributing factors, challenges and future perspectives. Eur J Lipid Sci Technol. 117(9), 1345-1369. DOI: 10.1002/ejlt.201400469. [ Links ]
Fedele, V., Claps, S., Rubino, R., Calandrelli, M. & Pilla, A.M., 2002. Effect of free-choice and traditional feeding systems on goat feeding behaviour and intake. Livest. Prod. Sci. 74, 19-31. [ Links ]
Fernández, M., Ordonez, J.A., Cambero, I., Santos, C., Pin, C. & De la Hoz, L., 2007. Fatty acid compositions of selected varieties of Spanish dry ham related to their nutritional implications. Food Chem. 9, 107-112. [ Links ]
LeDoux, M., Rouzeau, A., Bas P. & Sauvant, D., 2002. Occurrence of trans-C18:1 fatty acid isomers in goat milk: Effect of two dietary regimens. J. Dairy Sci. 85, 190-197. [ Links ]
Lopez, A., Vasconi, M., Moretti, V.M. & Bellagamba, F., 2019. Fatty acid profile in goat milk from high- and low-input conventional and organic systems. Animals, 9, 452-466; doi:10.3390/ani9070452. [ Links ]
Mancilla-Leytón, J.M., Martin, V.A. & Parejo-Farnés, C., 2014. A vegetation management experiment: Goats grazing shrublands in Donana Natural Park. Russ J Ecol 45, 384-390. [ Links ]
Mele, M., Serra, A., Buccioni, A., Conte, G., Pollicardo, A. & Secchiari, P., 2008. Effect of soybean oil supplementation on milk fatty acid composition from Saanen goats fed diets with different forage: concentrate ratios. Ital. J. Anim. Sci., 7, 297-311. [ Links ]
Mierlita, D., 2016. Fatty acid profile and health lipid indices in the raw milk of ewes grazing part-time and hemp seed supplementation of lactating ewes. S. Afr. J. Anim. Sci. 46(3), 237-246. [ Links ]
Mierlita, D., Pop, I.M., Teusdea, A., Lup, F., Dãrãban, S., Georgescu, B., Boaru, A. & Rahmann G., 2017. Effect of forage preservation method on fatty acid composition and oxidative stability of organic sheep milk. Landbauforsch Appl. Agric. Forestry Res. 67(1), 43-52. [ Links ]
Min, B.R., Hart, S.P., Sahlu, T. & Satter, L.D., 2005. The effect of diets on milk production and composition, and on lactation curves in pastured dairy goats. J. Dairy Sci. 88, 2604-2615. [ Links ]
Moloney, A.P., Fievez, V., Martin, B., Nute, G.R. & Richardson, R.I., 2008. Botanically diverse forage-based rations for cattle: Implications for product composition and quality and consumer health. Grassl. Sci. in Europe 13, 361-374. [ Links ]
Nudda, A., Cannas, A., Correddu, F., Atzori, A.S., Lunesu, M.F., Battacone, G. & Pulina, G., 2020. Sheep and goats respond differently to feeding strategies directed to improve the fatty acid profile of milk fat. Animals, 10, 1290-1312; doi:10.3390/ani10081290. [ Links ]
Pilarczyk, R., Wójcik, J., Sablik, P. & Czerniak, P., 2015. Fatty acid profile and health lipid indices in the raw milk of Simmental and Holstein-Friesian cows from an organic farm. S. Afr. J. Anim. Sci. 45, 30-38. [ Links ]
Rego, O.A., Cabrita, A., Rosa, H., Alves, S., Duarte, V., Fonseca, A., Vouzela, C., Pires, F. & Bessa R., 2016. Changes in milk production and milk fatty acid composition of cows switched from pasture to a total mixed ration diet and back to pasture. Ital. J. Anim. Sci., 15(1), 76-86. [ Links ]
Renna, M., Lussiana, C., Cornale, P., Fortina, R. & Mimosi, A., 2012. Changes in goat milk fatty acids during abrupt transition from indoor to pasture diet. Small Rumin. Res. 108, 12-21. [ Links ]
Rubino, R., Moioli, B., Fedele, V., Pizzillo, M. & Morand, P., 1995. Milk production of goats grazing native pasture under different supplementation regimes in Southern Italy. Small Rum. Res. 17, 213-221. [ Links ]
Rufino, M., Alves, A.A., Marcônio, M.R., Moura, R.L., Cavalcante A.C. & Rogério, M.C., 2012. Goat milk production and quality on Tanzania-grass pastures, with supplementation. Acta Scientiarum, Anim. Sci. 34(4), 417-423. doi: 10.4025/actascianimsci.v34i4.14339. [ Links ]
SAS, 2001. SAS System for Microsoft Windows. Release 8.2. SAS Institute Inc., Cary, NC., USA. [ Links ]
Simopoulos, A.P., 2002. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 56, 365-379. [ Links ]
Sukhija, P.S. & Palmquist, D.L., 1988. Rapid method for determination of total fatty acid content and composition of feedstuffs and faeces. J. Agric. Food Chem. 36, 1202-1206. [ Links ]
Thomas, L. & Rowney, M., 1996. Australian milk fat survey fatty acid composition. Aust. J. Dairy Tech. 51, 112-114. [ Links ]
Tsiplakou, E., Mountzouris, K.C. & Zervas, G., 2006. Concentration of conjugated linoleic acid in grazing sheep and goat milk fat. Livest. Sci. 103, 74-84. doi: 10.1016/j.livsci.2006.01.010. [ Links ]
Ulbricht, T. & Southgate, D., 1991. Coronary heart disease: Seven dietary factors. Lancet, 338, 985-992. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Submitted 15 December 2021
Accepted 8 May 2022
Published 28 January 2023
# Corresponding author: dadi.mierlita@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}