










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 no.5 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i5.01
Effect of lupin flake supplementation on rumen fermentation and meat composition of Hanwoo steers
K.H. Um J.S. Shin; B.K. Park#
Department of Animal Science, Kangwon National University, Chunchoen 24341, Korea
ABSTRACT
Lupin is considered a potentially rich source of energy and protein for various livestock breeds. However, there are no studies on rumen fermentation and meat composition of Hanwoo (Korean native) steers following supplementation with lupin flakes. In vitro and in situ experiments of lupin flake-supplemented feed were conducted using three Hanwoo cows with rumen fistulae for four weeks. The feeding experiment was conducted using 40 early-fattening Hanwoo steers randomly divided into four groups for 14 months from 16 to 30 months of age, viz., T1, T2, T3, and T4, with feed containing 0%, 3%, 6%, and 9% lupin flakes, respectively. The pH of the ruminal fluid in vitro at 24 and 48 h was higher in T3 and T4 than T1. The rumen levels of acetate, propionate, butyrate, and total volatile fatty acids after 48 h of incubation were lower in the lupin-supplemented groups than in T1. The ruminal pH in situ after 9 and 12 h was higher in T3 than in T1. The strip loin in Hanwoo steers was lighter in T3 than in the other treatment groups. The levels of carnosine, creatinine, adenosine triphosphate, and adenosine monophosphate in the strip loin were higher in T3 and T4 than in T1. The levels of oleic acid and unsaturated fatty acids and the n-6/n-3 ratio tended to be higher in T3 than in T1. Supplementation of livestock feed with 6% lupin flakes had positive effects on ruminal pH and meat lightness, carnosine, creatinine, adenosine triphosphate, and adenosine monophosphate levels in Hanwoo beef.
Keywords: dipeptide, physicochemical characteristics, rumen, strip loin, unsaturated fatty acid
Introduction
Lupin (Lupinus polyphllus lindl) is considered a potentially rich source of energy and proteins for various livestock breeds (Nalle et al., 2011). Lupin has high protein (32%), energy (5.5% fat and 24.3% non-fibrous carbohydrate), and unsaturated fatty acid (80%) contents (Bertoglio et al., 2011). Furthermore, it contains more than 35% monounsaturated fatty acids (MUFA) (mostly oleic acid), which influence the taste of beef (Boschin et al., 2008; Calabrò et al., 2015). Therefore, it is used as a raw material for food and livestock feed worldwide (Boschin et al., 2011; Elbandy et al., 2014).
Laudadio & Tufarelli (2011) reported that the ratio of saturated to unsaturated fatty acids and the n-6/n-3 ratio decreased on administration of lupin-supplemented diets to broilers. In addition, Froidmont et al. (2005) reported that a lupin-supplemented feed not only improved the carcass grade but also reduced the saturated fatty acid/unsaturated fatty acid and n-6/n-3 ratios in pork. Moss et al. (1997) reported that the feeding of lupin had no effect on growth performance in young calves. Murphy & McNiven (1994) found that average daily gain was improved by supplementing growing beef cattle with heat-treated lupin.
Ammonia generated during the rumen fermentation of feed proteins is used for microbial protein synthesis (Maeng et al., 1976). The crude protein content of microorganisms in the rumen is 8-12% (dry matter basis); as protein digestibility in the rumen is 70-90%, 40-90% of the protein required by ruminants is constituted of microbial protein (Putri et al., 2019). The rumen by-pass protein content of lupin is more than 30% (Park, 2005). Furthermore, the low starch content of lupin does not cause acidosis in beef cattle (Park, 2005). In addition, lupin is known to have high protein utilization and absorption rates because of its highly effective protein degradability (Nowak and Wylegala, 2005).
Recently, there has been a demand for an additional supply and an improvement in the quality of protein and energy to shorten the fattening period of Hanwoo steers (Jeong et al., 2010). Lupin is used as a high-energy and high-protein raw material for livestock feed (Nalle et al., 2011); however, there are no studies on rumen fermentation and meat composition of Hanwoo steers following supplementation of feed with lupin flakes. Therefore, this study was conducted to examine the effects of lupin-flake supplementation on the rumen fermentation characteristics and meat composition of Hanwoo steers.
Materials and Methods
This study was approved by the Animal Experimental Ethics Committee of the Kangwon National University, South Korea (approval number: KW-200820-2). Three Hanwoo cows (423.0 ± 44.8 kg), each implanted with a ruminal fistula, were used for four weeks. The cows were raised in pens (one cow per pen) and were offered formula feed along with roughage twice daily at 09h00 and 18h00. The cows had free access to water and mineral blocks. Other feeding management procedures were conducted according to the practices of the experimental farm.
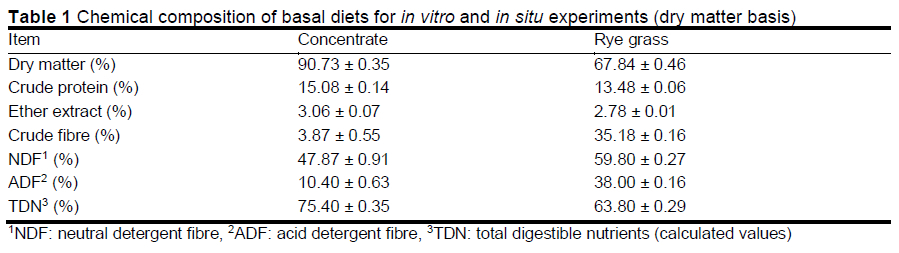
The experimental diets were divided into four feed rations, namely T1, T2, T3, and T4, containing 0%, 3%, 6%, and 9% lupin flakes, respectively. The chemical compositions of the experimental diets are listed in Table 1. The feed rations were crushed with a Wiley mill (Thomas Model 4 Wiley Mill, Thomas Scientific, New Jersey, USA) equipped with a 2-mm screen, and used in the in vitro and in situ ruminal experiments.
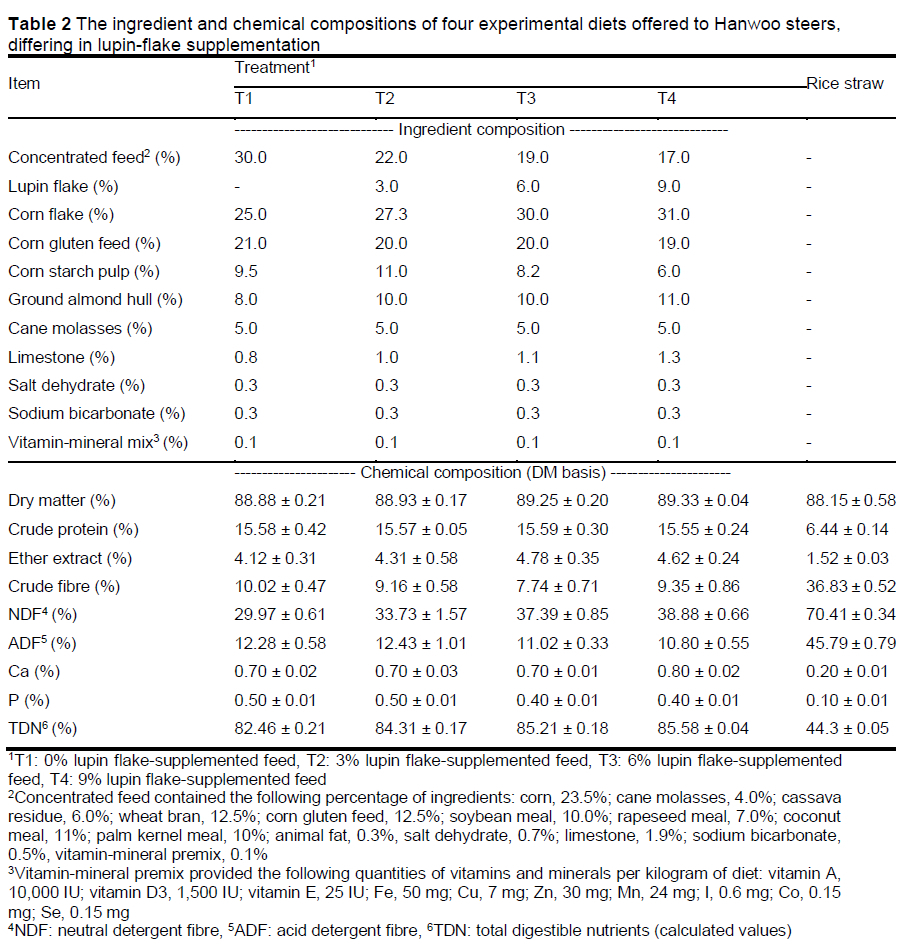
Feeding experiment: A total of 40 early-fattening Hanwoo steers (444.3 ± 32.9 kg) were randomly divided into four groups for 14 months from 16 to 30 months of age, viz. T1, T2, T3, and T4, receiving a 0%, 3%, 6%, or 9% lupin flake-supplemented feed ration, respectively. The steers were raised in eight pens (five steers per pen) and were offered the feed ration and roughage thrice daily at 08:30, 13:00, and 17:00 h. The ingredients and chemical composition of the experimental diets are listed in Table 2. The steers had free access to water and mineral blocks. Other feeding management procedures were conducted according to the practices of the experimental farm.
Ruminal fluid was collected from the rumen fistulas of Hanwoo cows prior to feeding at 09.00 and filtered through four layers of cheesecloth before being stored at 39 °C in a thermos flask. This was followed by the infusion of O2-free CO2 gas for 30 s to eliminate air from the flask. The collected ruminal fluid was transferred to the laboratory and allowed to stand for 1 h in an incubator at 39 °C to eliminate feed particles, prior to use as an inoculum for incubation in vitro.
Cultures were established by adding 400 mL of rumen inoculum to 1,596 mL of previously prepared artificial saliva (composed of buffer solutions A and B) following the method of McDougall (1948). Briefly, 1,330 mL of buffer solution A (containing 10.0 g KH2PO4, 0.5 g MgSO4-7H2O, 0.5 g NaCl, 0.1 g CaCb2H2O, and 0.5 g urea per litre of distilled water) and 266 mL of buffer solution B (containing 15.0 g NaCO3 and 1.0 g Na2S9H2O per litre of distilled water) were mixed simultaneously, and the pH was adjusted to 6.8.
A total of 70 mL of the prepared culture solution was transferred to 100 mL bottles (120 ml glass bottle), each containing 2 g of the experimental diets, and O2-free CO2 gas was infused for 5 s to eliminate air from the culture bottles. The bottles were then incubated at 39 °C for 0, 3, 6, 9, 12, 24, and 48 h in a shaking incubator (HB- 201SLI, Hanbaek Scientific Co, Bucheon, Korea).
The ruminal pH of the culture solution for each incubation period was measured using a pH meter (Starter 2100, OHAUS Corporation, New Jersey, USA) in a 100 mL bottle. To analyse ammonia concentration, 10 mL of the culture solution for each incubation period was collected in a 100 mL bottle and centrifuged (3,000 χ g, 15 min, 4 °C). This was followed by mixing of 5 mL of the supernatant with 0.05 mL HgCl2 prior to another step of centrifugation at 3,000 χ g and 15 min for 4 °C. Then, 1 mL of the supernatant was collected, and the ammonia concentration was analysed according to the method of Chaney and Marbach (1962).
To analyse volatile fatty acid (VFA) concentrations, 10 mL of the culture solution for each incubation period was collected in a 100 mL bottle. This was followed by addition of 1 mL of 20% HPO3 and 0.5 mL of saturated HgCb and centrifugation at 1,250 χ g and 4 °C for 15 min. The supernatant was then collected, and VFA concentration was measured using gas chromatography (6890N, Agilent Technologies, Palo Alto, CA, USA).
Evaluation of rumen fermentation in situ in Hanwoo cows: Rumen fluid was collected from Hanwoo cows fitted with rumen fistulas at 0, 3, 6, 9, 12, 24, and 48 h after a 14-day feed adaptation period. The pH of 5 mL of the ruminal fluid at each incubation time point was measured using a pH meter (Starter 2100, OHAUS Corporation, New Jersey, USA). To analyse ammonia concentration, 10 mL of rumen fluid collected at each incubation time point was centrifuged (3,000 χ g, 15 min, 4 °C) and then 5 mL of the supernatant and 0.05 mL of HgCl2 were mixed and centrifuged again (3,000 χ g, 15 min, 4 °C). One millilitre of the supernatant was then collected, and ammonia concentration was analysed according to the method of Chaney and Marbach (1962).
Five grams of each experimental diet was placed in a nylon bag (ANKOM 5 χ 10 concentrate bags) and inserted into the rumen of the Hanwoo cows via a fistula prior to feeding at 09.00. Experiments were performed by collecting nylon bags at 0, 3, 6, 9, 12, 24, and 48 h. The nylon bag was washed until the water ran clear and was dried in a 70 °C forced air drying oven for 72 h. The dried samples were weighed, and the weight was used to calculate the disappearance of the dry matter (DM). The samples were then ground to a particle size of 1.0 mm, and their chemical composition was analysed to determine the disappearance of crude protein.
Evaluation of the physicochemical composition of beef from Hanwoo steers: Strip loin samples from cold carcasses of Hanwoo steers were used for meat composition analysis. The carcass was transported to the laboratory, and its quality was analysed in a low-temperature room (5 °C) after the removal of fat, connective tissue, and blood.
The meat surface colour of the strip loin was measured using a Chroma Meter (Colorimeter CR-300, Minolta Co., Osaka, Japan). The following colour parameters were determined: lightness (L*), redness (a*), and yellowness (b*). For standardization, a white plate reference was used with a Y value of 93.60, an x value of 0.3134, and a y value of 0.3194. Myoglobin content was analysed according to the method described by Trout (1989).
To analyse myoglobin content, a strip loin sample (2 g) was homogenized with phosphate buffer (20 mL, 40 mM, pH 6.8) at 11,200 χ g for 30 s. The homogenate was centrifuged at 3,000 χ g for 10 min and filtered through a Whatman No. 1 filter paper (Cytiva, UK). The absorbance of the filtrate was measured at 700 and 525 nm using a microplate reader (VersaMax ELISA microplate reader, Molecular Devices, CA, USA). The chemical composition of the strip loin was determined on the basis of moisture, crude protein, ether extract, and crude ash contents, according to the standard AOAC (1995) methods.
To measure the pH, a strip loin sample (10 g) was homogenized with distilled water (90 mL) for 1 min at 20,000 χ g in a homogenizer (IKA ULTRA-TURRAX, Ika Werkes, Staufen, Germany). The pH of the homogenate was measured using a pH meter (Starter 2100, OHAUS Corporation, New Jersey, USA). To measure the water-holding capacity (WHC), a strip loin sample (0.5 g) was heated in a constant-temperature water bath at 80 °C for 20 min. It was then allowed to cool for 10 min, centrifuged at 2000 χ g for 20 min, and weighed.
Thiobarbituric acid-reactive substance (TBARS) content of the strip loin was measured according to the method of Buege & Aust (1978). Briefly, a strip loin sample (5 g) was homogenized in distilled water (15 mL). To inhibit the oxidation reaction, the homogenate (1 mL) was incubated with 7.2% butylated hydroxyanisole (50 μL). The mixture was then added with 2 mL of trichloroacetic acid (TCA)/ thiobarbituric acid (TBA) reagent, heated at 90 °C for 15 min, cooled in cold water, and centrifuged at 2000 χ g for 10 min. The absorbance of the supernatant, and a blank sample (prepared in a similar manner using only distilled water), at 531 nm was measured using a microplate reader (VersaMax ELISA microplate reader, Molecular Devices, CA, USA). The TBARS value was calculated by multiplying the absorbance value by 5.88.
The contents of carnosine, anserine, creatine, and creatinine in the strip loin were analysed using the methods described by Mora et al. (2007). Briefly, a strip loin sample (2.5 g) was homogenized with 7.5 mL of 0.01 N HCl at 11,200 χ g for 30 sec. The homogenate was then centrifuged at 3000 χ g for 30 min at 4 °C, and the supernatant was filtered using a Whatman GF/C glass microfibre filter (Cytiva, UK). The filtrate (250 μL) was then incubated with 750 μL acetonitrile at 4 °C for 20 min, followed by centrifugation at 10,000 χ g for 10 min. The supernatant was then filtered through a 0.22 μηι membrane filter and subjected to high-performance liquid chromatography (HPLC) analysis using an HPLC system (Agilent Infinity 1260 Series, Agilent Technologies, CA, USA).
The contents of hypoxanthine (HX), inosine monophosphate (IMP), adenosine monophosphate (AMP), and adenosine triphosphate (ATP) in the strip loin were analysed using the methods described by Lee et al. (1987). Briefly, a strip loin sample (5 g) was homogenized with 0.7 M perchloric acid (PCA) using a homogenizer. The homogenate was neutralized with 5 N KOH and then centrifuged at 2000 χ g for 15 min. The supernatant was filtered through a Whatman No. 1 filter paper (Cytiva, UK) and then neutralized to pH 6.5 with 5 N KOH. The sample volume was adjusted to 50 mL by adding neutralized PCA; the solution was then filtered through a 0.22 μm membrane filter and subjected to HPLC analysis using an HPLC system (Agilent Infinity 1260 Series, Agilent Technologies, CA, USA).
To measure fatty acid composition, a strip loin sample (10 g) was incubated with 200 mL of an organic solvent mixture (chloroform:methanol = 2:1) and 6 mL of 0.88% KCl and stirred for 3 min. The sample was then centrifuged at 3000 χ g for 10 min, followed by separation of the lipid layer. This process was repeated thrice, followed by the concentration of extracted lipids using nitrogen gas. The lipid methylation process was performed according to the method described by Morrison & Smith (1964). Briefly, the concentrated lipid fraction (10 mg) was incubated with 1 mL of freshly prepared 0.5 N methanolic NaOH (2 g NaOH/100 mL methanol) in a saponification reaction vessel, heated for 15 min, and then cooled. The mixture was then incubated with 2 mL of BF3-methanol (a methylation reagent), heated for 15 min, and cooled to room temperature (approximately 20 °C). The sample was then incubated with heptane (1 mL) and a saturated NaCl solution (2 mL) for 1 min and maintained at room temperature for 30 min. Fatty acid content was analysed by injecting 1 μL of the supernatant into a gas chromatography (GC) system (ACEM 6000 Model, Youngin Scientific, Seoul, Korea). The conditions for GC were as follows: column, Omegawax 320 capillary column (100 m χ 0.32 mm I. D., 0.25 μm film); carrier gas, nitrogen; nitrogen flow rate, 1 mL/min; injection port temperature, 240 °C; detector temperature, 250 °C; oven temperature, 160 °C; and split ratio, 10:1.
For amino acid analysis of the strip loin, the sample was weighed to contain about 30 mg of crude protein. The sample and 40 mL of 6 N HCl were placed in a decomposition bottle, mixed, and then hydrolysed at 110 °C for 24 h. The resultant solution was then concentrated via rotary evaporation, diluted with distilled water, and filtered through a filter paper. The filtered samples were analysed using an amino acid analyser (L-8900, Hitachi, Tokyo, Japan).
Statistical analyses to obtain the average values and standard differences were performed using IBM SPSS (Statistical Package for the Social Sciences, SPSS Inc., Chicago, IL, USA). A one-way analysis of variance was used to calculate the average value for each treatment group. Statistically significant differences between the treatments were determined using Duncan's multiple range test (Duncan, 1955) at a level of significance of P <0.05.
Results and Discussion
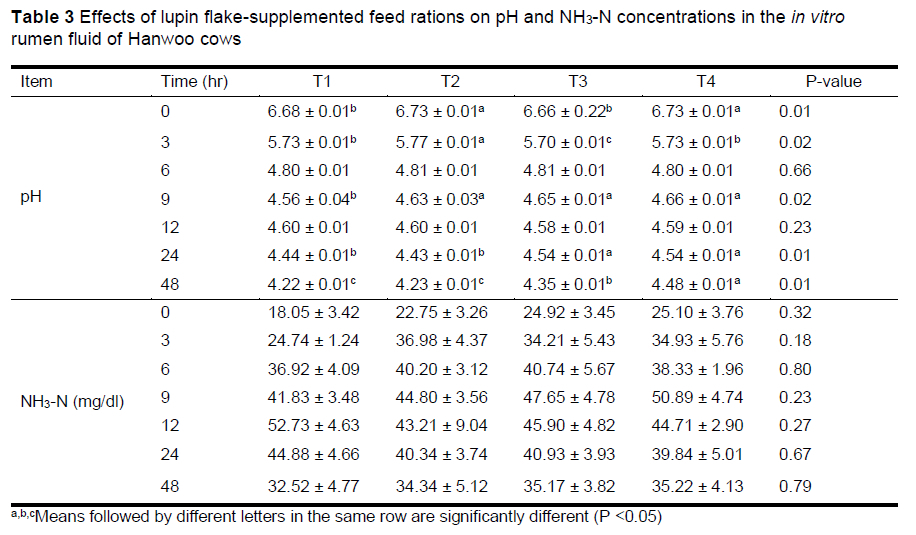
Table 3 depicts the effects of lupin flake supplementation on the pH and ammonia level of the rumen fluid culture solution in vitro. The ruminal pH at 9 h was higher in the lupin flake-supplemented groups than in the T1 group. It was also higher in the T3 and T4 groups than in the T1 group at 24 and 48 h (P <0.05). Although there was no statistical difference, ammonia levels tended to be higher in the lupin flake-supplemented groups than in the T1 group until 9 h. Shim (1992) reported that lupin supplementation led to a slight decrease in rumen pH compared to supplementation with other cereals because it had a buffering effect. Furthermore, Miyaji et al. (2012) reported that the rumen levels of ammonia changed according to the grain variety, processing method, and feeding conditions. In the current study, the rumen pH remained relatively high following an increase in ammonia level because of the supplementation of lupin flakes. Therefore, the authors postulate that the supplementation of lupin flakes has a positive effect on rumen pH and ammonia levels.
Table 4 depicts the effects of lupin flake supplementation on VFA content in the ruminai fluid. The contents of acetate, propionate, and total-VFA at 3 h were lower in the lupin flake-supplemented groups than in the T1 group (P <0.05), and acetate content at 9 h was lower in the T2 group than in the other groups (P <0.05). Propionate contents at 12 h and 24 h were lower in the T3 group than in the other supplemented groups (P <0.05). The contents of acetate, propionate, butyrate, and total VFAs at 48 h were lower in the T2 and T3 groups than in the T1 group (P <0.05).
Contents of VFAs vary depending on the type of grain, processing method, and management conditions (Miyaji et al., 2012). In addition, non-structural carbohydrates such as starch are converted to propionate and lactic acid by rumen microbes, which affects the rumen pH (Nishino et al., 2004; Keles & Demirci, 2011). Thus, when the starch content is low, the digestible protein synthesis of microorganisms is lowered and the concentration of VFAs is reduced (Keles & Demirci, 2011). In this study, it was considered that the low propionate and acetate levels in the lupin-supplemented groups after 3 h were related to the low starch (Park, 2005) and fibre (Table 2) contents of lupin.
Table 5 depicts the effects of lupin flake supplementation on the pH, NH3-N concentrations, DM, and crude protein disappearance rates in the rumen. Rumen pH was higher in proportion to the lupin flake-supplemented levels at 3 h (P <0.05), and the pH at 9 h and 12 h was higher for the T3 group than for the T1 group (P <0.05). The rumen ammonia levels tended to be higher in the lupin flake-supplemented groups than in the T1 group during most of the incubation period, but the difference was not significant. The disappearance rates of DM at 3, 9, and 48 h tended to be lower in the T3 and T4 groups than in the T1 group. The disappearance rate of crude protein at 6 h was lower in the lupin flake-supplemented groups than in the T1 group (P < 0.05). In particular, the disappearance rates of crude protein at 9, 12, and 24 h were lower in the T4 group than in the T1 group (P <0.05).
In this study, rumen pH and ammonia levels showed a tendency to increase during most of the rumen fermentation period in the lupin-supplemented groups compared with those in the T1 group, which can be attributed to the low starch and high rumen degradable protein contents of lupin flakes (Park, 2005).
The disappearance rates of rumen dry matter and crude protein showed a tendency towards lower concentrations in the lupin-supplemented groups than in the T1 group, which can be attributed to the difference in composition of ingredients between the feed rations (Table 2). The T1 ration had a relatively high ratio of ingredients (concentrated feed and corn gluten feed), which induced a higher rumen degradation rate compared with that of the lupin-supplemented groups.
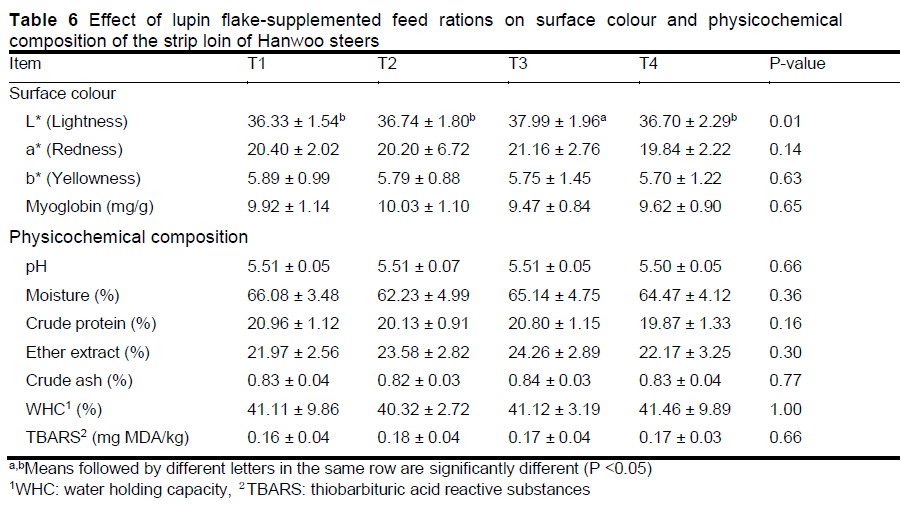
Table 6 depicts the effects of lupin flake supplementation on surface colour and physicochemical composition of the strip loin of Hanwoo steers. The lightness (L*) of the strip loin was higher in the T3 group than in the other treatment groups (P <0.05). Redness (a*), yellowness (b*), and the levels of myoglobin were similar among the treatment groups. Supplementation with lupin flakes did not affect the pH, moisture, crude protein content, ether extract, crude ash content, WHC, or TBARS in the strip loin.
Surface colour is an important factor in determining the quality of meat. Myoglobin in muscle cells is responsible for storing and distributing oxygen (Livingston et al., 1983). Previous studies (Vicenti et al., 2009; Lestingi et al., 2016) reported that supplementation with lupin did not affect meat surface colour. In addition, Lestingi et al. (2016) reported that supplementation with lupin reduces the content of myoglobin. However, in the current study, 6% lupin flake supplementation had a positive effect on surface lightness in the strip loin of Hanwoo steers. Thus, there are some differences between the present study and previous studies regarding the effect of lupin flake supplementation on the surface colour of the strip loin of Hanwoo steers.
The quality of beef is affected by its chemical properties (Muchenje et al., 2009). Vienti et al. (2009) and Lestingi et al. (2016) reported that lupin supplementation had no effect on the chemical composition of beef, which is consistent with the results of the present study. Further, Lestingi et al. (2016) reported that lupin supplementation did not affect the pH of strip loin, which was again in accordance with the results of the current study. TBARS values indicate the content of malondialdehyde (which is the final product of lipid peroxidation) and the rancidity of meat (Cho et al., 2019). TBARS values of <0.2 mg and 4.0 mg malondialdehyde/kg meat indicate the freshness and rancidity of meat, respectively (Brewer et al., 1992; Sung et al., 2013). In the current study, although the value of TBARS was not investigated during the storage period, it was found to be less than 0.2 mg malonaldehyde (MA)/kg for all treatment groups. Therefore, it was presumed that lupin flake supplementation had little effect on pH, moisture, crude protein content, ether extract, crude ash content, WHC, and TBARS in the strip loin of Hanwoo steers.
Table 7 depicts the effects of lupin flake supplementation on dipeptide and nucleic acid content of the strip loin of Hanwoo steers. Lupin flake supplementation had little effect on anserine and creatine contents of the strip loin. Carnosine and creatinine contents were higher in the T3 and T4 groups than in the T1 group (P <0.05). The HX, inosine, and IMP contents of the strip loin were similar in all the treatment groups and ATP and AMP contents were higher in the T3 and T4 groups than in the T1 group (P <0.05).
The dipeptides, including carnosine, anserine, creatine, and creatinine, are the functional and bioactive components of meat (Peireti et al., 2012). Creatine and creatinine, in particular, are involved in muscle energy metabolism and provide the energy required for muscle contraction (Mora et al., 2007). Carnosine is involved in imparting the characteristic taste of meat and the savoury taste (Ahn & Park, 2002) and, like anserine, is known to influence the antioxidant properties of tissues (Decker, 2000). The results of the current study suggest that lupin flake supplementation can help to improve the flavour of beef by increasing carnosine content in the strip loin of Hanwoo steers. The nucleotides in meat are converted to IMP by AMP generated in the muscle during ATP hydrolysis. Inosine, produced from IMP, is converted to HX during the storage period (Komatsu et al., 2019). IMP has been reported to influence the taste of meat (Rotola-Pukkila et al., 2015; Li et al., 2016). Although changes in the levels of nucleic acid-related substances (IMP and inosine) were not investigated during the storage period in this study, it was observed that ATP and AMP contents of the strip loin increased with lupin flake supplementation. Therefore, it is expected that lupin flake supplementation might lead to an increase in the contents of IMP and inosine.
Because the activity of AMP deaminase remains constant regardless of the site (Aberle and Merkel, 1968), 5'-nucleotidase, which degrades IMP, exhibits a constant activity (Tsai et al., 1972). In this study, the increase in the contents of ATP and AMP in the strip loin following 6% or 9% lupin flake supplementation was regarded as being caused by increased muscle ATP accumulation during the fattening period. Therefore, it can be considered that the increase in the energy content of the feed ration, rather than the supplementation of lupin, affects the content of the nucleic acid substances (ATP and AMP) that determine the flavour of beef.
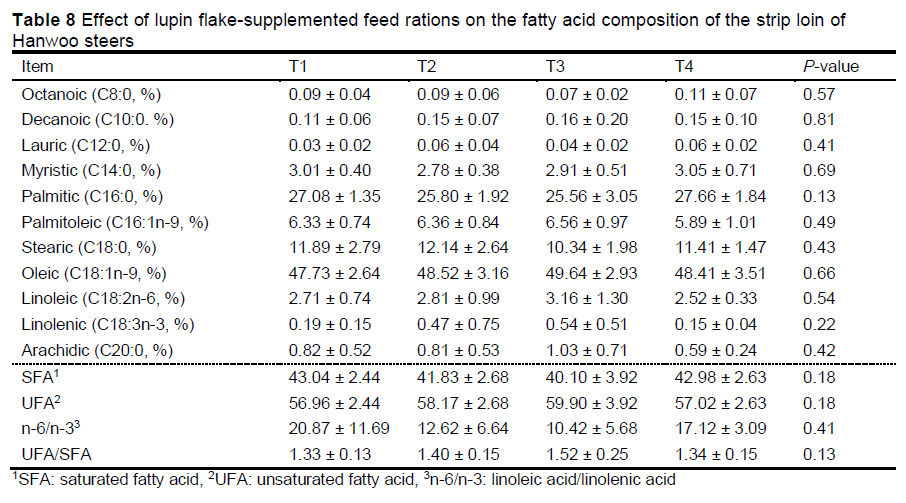
Table 8 depicts the effects of lupin flake supplementation on fatty acid composition of the strip loin of Hanwoo steers. The oleic acid and unsaturated fatty acid (UFA) contents of the strip loin were slightly, but not significantly, higher in the lupin flake-supplemented formulated feed groups (especially the T3 group) than in the T1 group. The n-6/n-3 ratios in the strip loin tended to be lower in the lupin flake-supplemented feed ration groups (especially the T3 group) than in the T1 group, but the difference was not significant.
Fatty acid is an important component that determines the characteristics, quality of fat, and the palatability of beef (Anderson et al., 1975). Amongst fatty acids, oleic acid is critical to the taste and flavour of meat (Lee et al., 2004; Cho et al., 2008). Furthermore, the ratio of MUFA to saturated fatty acid can be an indirect indicator of meat flavour (Anderson, 1976). Lupin has a high unsaturated fatty acid (UFA) content (80%), with more than 35% MUFA (mostly oleic acid) and 10% linolenic acid (Uzun et al., 2007; Boschin et al., 2008; Calabrò et al., 2015). In terms of the fatty acid composition of lupin, the authors predicted that fatty acid composition of the strip loin might be altered following supplementation of the formula feed with lupin. However, in this study, there was no significant effect of lupin flake supplementation on the fatty acid composition of the strip loin. Nevertheless, addition of 6% lupin flakes resulted in some positive effects on the levels of oleic acid and UFA and the n-6/n-3 ratio.
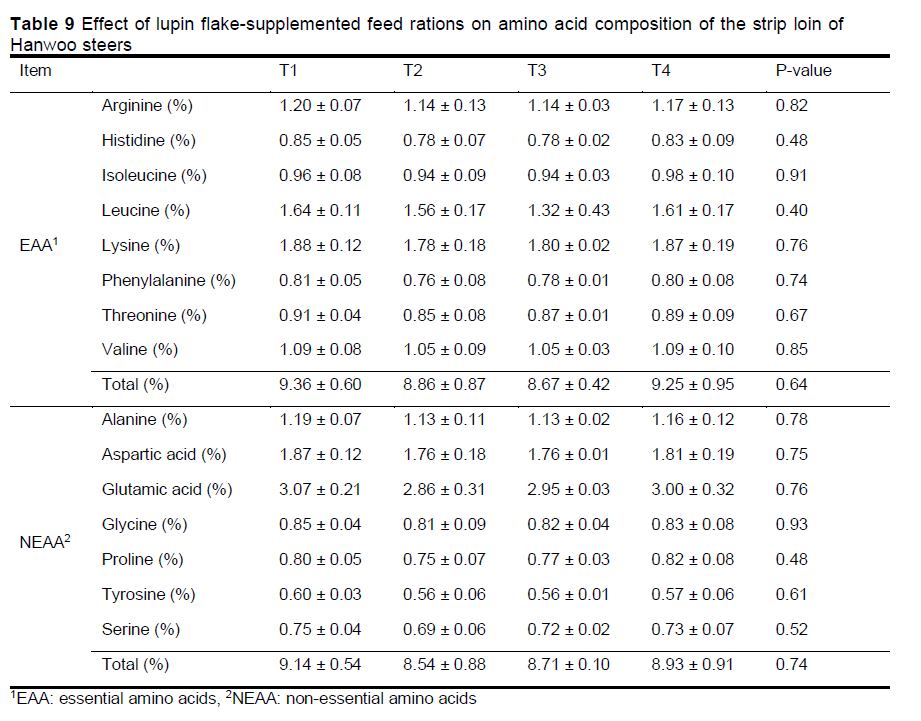
Lupin flake supplementation did not affect the individual, essential, non-essential, or total amino acid composition of the strip loin of Hanwoo steers (Table 9). This could have been because the protein content of the formulated feeds was the same in all the treatment groups. The authors suggest that there was no difference in the amino acid composition of beef because there was no difference in the amino acid supply in the feed rations.
Conclusion
Based on the results of this study, it can be inferred that supplementation of livestock feed with lupin flakes helps to maintain a stable pH without affecting the NH3-N or volatile fatty acid content in the rumen. In addition, it is suggested that supplementation of livestock feed with 6% lupin flakes has a positive effect on the surface lightness and the levels of carnosine, creatinine, ATP, and AMP in the strip loin of Hanwoo steers. Therefore, the results of the present study indicate that the appropriate level of supplementation of lupin flakes is 6% in Hanwoo steers. However, since the size of the group (number of steers) was small in this study, further studies with increased group size may be needed to determine the optimal level of lupin flake supplementation.
Acknowledgements
The authors would like to thank Editage (www.editage.co.kr) for English language editing.
Authors' contributions
KHU collected the data for this study, conducted the statistical analyses, collaborated in the interpretation of the results, and finalized the manuscript; J.S. Shin collaborated in interpreting the results and wrote the initial draft of this manuscript; BKP developed the original hypotheses, designed the experiments, collaborated in interpreting the results, and wrote the initial draft of this manuscript.
References
Aberle, E.D. & Merkel, R.A., 1968. 5'-Adenylic acid deaminase in porcine muscle. J. Food Sci. 33(1), 27-29. doi.org/10.1111365-2621.1968.tb00878.x. [ Links ]
Ahn, D.H. & Park, S.Y., 2002. Studies on components related to taste such as free amino acids and nucleotides in Korean native chicken meat. J. Korean Soc. Food Sci Nutr. 31(4), 547-552. doi.org/10.3746/jkfn.2002.31.4.547. [ Links ]
Anderson, B.A., Kisellan, J.A. & Watt, B.K., 1975. Comprehensive evaluation of fatty acids in foods. II. Beef products. J. Amer. Diet Assoc. 67, 35-41. [ Links ]
Anderson, B.A., 1976. Comprehensive evaluation of fatty acids in foods, VII, Pork products. J. Amer. Diet Assoc. 69, 44-49. [ Links ]
AOAC, 1995. Association of Official Analytical Chemists. Official methods of analysis, 16th ed. Washington DC, USA. [ Links ]
Bertoglio, J.C., Calvo, M.A., Hancke, J.L., Burgos, R.A., Riva, A., Morazzoni, P., Ponzone, C., Magni, C. & Duranti, M., 2011. Hypoglycemic effect of lupin seed γ-conglutin in experimental animals and healthy human subjects. Fitoterapia 82(7), 933-938. doi.org/10.1016/j.fitote.2011.05.007 [ Links ]
Boschin, G., & Arnoldi, A., 2011. Legumes are valuable sources of tocopherols. Food Chem. 127(3), 1199-1203. doi.org/10.1016/j.foodchem.2011.01.124. [ Links ]
Boschin, G., D'Agostina, A., Annicchiarico, P., & Arnoldi, A., 2008. Effect of genotype and environment on fatty acid composition of Lupinus albus L. seed. Food Chem. 108(2), 600-606. doi.org/10.1016/j.foodchem.2007.11.016 [ Links ]
Brewer, M.S., Ikins, W.I.G. & Harbers, C.A.A., 1992. TBA values, sensory characteristics, and volatiles in ground pork during long-term frozen storage: Effects of packaging. J. Food Sci. 57(3), 558-563. doi.org/10.111365-2621.1992.tb08042.x. [ Links ]
Buege, J.A. & Aust, S.D., 1978. Microsomal lipid peroxidation. Methods Enzymol. 52, 302-310. doi.org/10.1016/S0076-6879(78)52032-6. [ Links ]
Calabrò, S., Cutrignelli, M.I., Lo Presti, V., Tudisco, R., Chiofalo, V., Grossi, M., Infascelli, F. & Chiofalo, B., 2015. Characterization and effect of year of harvest on the nutritional properties of three varieties of white lupine (Lupinus albus L.). J. Sci. Food Agri. 95(15), 3127-3136. doi.org/10.1002/jsfa.7049. [ Links ]
Chaney, A.L. & Marbach, E.P., 1962. Modified reagents for determination of urea and ammonia. Clin. Chem. 8, 130-132. doi.org/10.1093/clinchem/8.2.130 [ Links ]
Cho, H.U., Ko, W.S., Son, H.W., Lee, M.J., Song, H.J. & Park, J.H., 2008. Hematological and biochemical analysis of Korean indigenous cattle according to age. Kor. J. Vet. Serv. (KOJVS). 31(1), 137-147 [ Links ]
Cho, W.K., Jeong, C.H., Kim, H.J., Kim, J.E., Kim, B.R., Lee, S.S. & Moon, Y.H., 2019. Effects of supplemental concentrate during the late fattening phase on performance, carcass characteristics, biochemical composition, and sensory test of loin in TMR feeding of Korean steers. Ann. Anim. Resour Sci. 30(4), 156-164. doi.org/10.12718/AARS.2019.30.4.156. [ Links ]
Decker, E.A., Livisay, S.A. & Zhou, S.A., 2000. A re-evaluation of the antioxidant activity of purified carnosine. Bio Chem. 7, 901-906. [ Links ]
Duncan, D.B., 1955. Multiple range and multiple F tests. Biometrics 11, 1-42. doi.org/10.2307/3001478 [ Links ]
Elbandy, M. & Rho, J.R., 2014. New flavone-di-C-glycosides from the seeds of Egyptian lupin (Lupinus termis). Phytochem Lett. 9, 127-131. doi.org/10.1016/j.phytol.2014.05.006. [ Links ]
Froidmont, É., Wathelet, B., Beckers, Y., Romnée, J.M., Dehareng, F., Wavreille, J., Schoeling O., Decauwert, V., & Bartiaux-Thill, N., 2005. Improvement of lupin seed valorisation by the pig with the addition of alpha- galactosidase in the feed and the choice of a suited variety. Biotechnologie, Agronomie, Société et Environnement. 9(4), 225-235. hdl.handle.net/2268/60583. [ Links ]
Jeong, J., Seong, N.I., Hwang, I.K., Lee, S.B., Yu, M.S., Nam, I.S. & Lee, M.I., 2010. Ruminant nutrition: Effects of level of CP and TDN in the concentrate supplement on growth performances and carcass characteristics in Hanwoo steers during final fattening period. J. Anim. Sci. Technol. 52(4), 305-312. doi.org/10.5187/JAST.2010.52.4.305 [ Links ]
Keles, G., & Demirci, U., 2011. The effect of homofermentative and heterofermentative lactic acid bacteria on conservation characteristics of baled triticale-Hungarian vetch silage and lamb performance. J. Anim. Sci. Technol. 164(1-2), 21-28. doi.org/10.1016/j.anifeedsci.2010.11.017 [ Links ]
Komatsu, T., Komatsu, M. & Uemoto, Y., 2019. The NT5E gene variant strongly affects the degradation rate of inosine 5'-monophosphate under postmortem conditions in Japanese Black beef. Meat Sci. 158, 107893. doi.org/10.1016/j.meatsci.2019.107893. [ Links ]
Laudadio, V. & Tufarelli, V., 2011. Dehulled-micronised lupin (Lupinus albus L. cv. Multitalia) as the main protein source for broilers: Influence on growth performance, carcass traits, and meat fatty acid composition. J. Sci. Food Agric. 91(11), 2081-2087. doi.org/10.1002/jsfa.4426 [ Links ]
Lee, E.H., Oh, K.S., Ahn, C.B., Chung, B.G., Bae, Y.K. & Ha, J.H., 1987. Preparation of powdered smoked-dried mackerel soup and its taste compounds. Bull. Korean Fish Soc. 20(1), 41-51. [ Links ]
Lee, S.H., Yoon, D.H., Hwang, S.H., Cheong, E.Y., Kim, O.H. & Lee, C.S., 2004. Relationship between monounsaturated fatty acid composition and stearoyl-CoA desaturase mRNA level in Hanwoo liver and loin muscle. J. Anim. Sci. & Technol. 46(1), 7-14. doi.org/10.5187/JAST.2004.46.1.007. [ Links ]
Lestingi, A., Facciolongo, A.M., Jambrenghi, A.C., Ragni, M. & Toteda, F., 2016. The use of peas and sweet lupin seeds alone or in association for fattening lambs: Effects on performance, blood parameters and meat quality. Small Rumin. Res. 143, 15-23. doi.org/10.1016/j.smallrumres.2016.08.006 [ Links ]
Li, D., Qin, N., Zhang, L., Lv, J., Li, Q. & Luo, Y., 2016. Effects of different concentrations of metal ions on degradation of adenosine triphosphate in common carp (Cyprinus carpio) fillets stored at 4 °C: An in vivo study. Food Chem. 211, 812-818. doi.org/10.1016/j.foodchem.2016.05.120 [ Links ]
Livingston, D.J., La Mar, G.N. & Brown, W.D., 1983. Myoglobin diffusion in bovine heart muscle. Science 220(4592), 71-73. DOI: 10.1126/science.6828881 [ Links ]
Maeng, W.J., Van Nevel, C.J., Baldwin, R.L., & Morris, J.G., 1976. Rumen microbial growth rates and yields: Effect of amino acids and protein. Journal of Dairy Science 59(1), 68-79. doi.org/10.3168/jds.S0022-0302(76)84157-4 [ Links ]
McDougall, E.I., 1948. Studies on ruminant saliva. 1. The composition and output of sheep's saliva. Biochem. J. 43, 99-109.DOI 10.1042/bj0430099 [ Links ]
Miyaji, M., Matsuyama, H., Hosoda, K. & Nonaka, K., 2012. Effect of replacing corn with brown rice grain in a total mixed ration silage on milk production, ruminal fermentation, and nitrogen balance in lactating dairy cows. Anim. Sci. J. 83(8), 585-593. doi.org/10.1111/j.1740-0929.2011.00996.x [ Links ]
Mora, L., Sentandreu, M.A. & Toldrá, F., 2007. Hydrophilic chromatographic determination of carnosine, anserine, balenine, creatine, and creatinine. J. Agric. Food Chem. 55(12), 4664-4669. [ Links ]
Morrison, W.R. & Smith, L.M., 1964. Preparation of fatty acid methyl esters and dimethylacetals from lipids with boron fluoride-methanol. J. Lipid Res. 5(4), 600-608. doi.org/10.1016/S0022-2275(20)40190-7 [ Links ]
Muchenje, V., Dzama, K., Chimonyo, M., Strydom, P.E., Hugo, A. & Raats, J.G., 2009. Some biochemical aspects pertaining to beef eating quality and consumer health: A review. Food Chem. 112(2), 279-289. doi.org/10.1016/j.foodchem.2008.05.103 [ Links ]
Nalle, C.L., Ravindran, V. & Ravindran, G., 2011. Nutritional value of narrow-leafed lupin (Lupinus angustifolius) for broilers. Br. Poult. Sci. 52(6), 775-781. doi.org/10.1080/00071668.2011.639343 [ Links ]
Nishino, N., Wada, H., Yoshida, M. & Shiota, H., 2004. Microbial counts, fermentation products, and aerobic stability of whole crop corn and a total mixed ration ensiled with and without inoculation of Lactobacillus casei or Lactobacillus buchneri. Journal of Dairy Science. 87(8), 2563-2570. doi.org/10.3168/jds.S0022-0302(04)73381-0 [ Links ]
Nowak, W. & Wylegala, S., 2005. The effect of rapeseed oil on the ruminal degradability and intestinal protein digestibility of rapeseed meal, soyabean and lupin seed. J. Anim. Feed Sci. 14, 295. doi.org/10.22358/jafs/70543/2005 [ Links ]
Park, H.O., 2005. Nutritional and feed value of lupine seeds. Feed 55-63. [ Links ]
Putri, E.M., Zain, M., Warly, L., & Hermon, H., 2019. In vitro evaluation of ruminant feed from West Sumatera based on chemical composition and content of rumen degradable and rumen undegradable proteins. Veterinary World. 12(9), 1478. DOI: 10.14202/vetworld.2019.1478-1483 [ Links ]
Rotola-Pukkila, M.K., Pihlajaviita, S.T., Kaimainen, M.T. & Hopia, A.I., 2015. Concentration of umami compounds in pork meat and cooking juice with different cooking times and temperatures. J. Food Sci. 80(12), C2711-C2716. doi.org/10.1111/1750-3841.13127 [ Links ]
Shim, J.M., 1992. Effect of flaking and particle size of corn and lupin seed upon ruminal fermentation rate in vitro and in situ. Konkuk University Graduate School, Dissertation. [ Links ]
Sung, S.H., Bae, Y.S., Oh, S.H., Lee, J.C., Kim, H.J. & Jo, C., 2013. Possibility of instrumental differentiation of duck breast meat with different processing and storage conditions. Korean J. Food Sci. An. 33(1), 96-102. doi.org/10.5851/kosfa.2013.33.1.96 [ Links ]
Tsai, R., Cassens, R.G., Briskey, E.J. & Greaser, M.L., 1972. Studies on nucleotide metabolism in porcine longissimus muscle postmortem. J. Food Sci. 37(4), 612-617. doi.org/10.1111/j.1365-2621.1972.tb02705.x. [ Links ]
Uzun, B., Arslan, C., Karhan, M. & Toker, C., 2007. Fat and fatty acids of white lupin (Lupinus albus L.) in comparison to sesame (Sesamum indicum L.). Food Chem. 102(1), 45-49. Doi.org/10.1016/j.foodchem.2006.03.059 [ Links ]
Vicenti, A., Toteda, F., Di Turi, L., Cocca, C., Perrucci, M., Melodia, L. & Ragni, M., 2009. Use of sweet lupin (Lupinus albus L. var. Multitalia) in feeding for Podolian young bulls and influence on productive performances and meat quality traits. Meat Sci. 82(2), 247-251. Doi.org/10.1016/j.meatsci.2009.01.018 [ Links ]
Trout, G.R., 1989. Variation in myoglobin denaturation and color of cooked beef, pork, and turkey meat as influenced by pH, sodium chloride, sodium tripolyphosphate, and cooking temperature. J. Food Sci. 54(3), 536-540. doi.org/10.1111/j.1365-2621.1989.tb04644.x. [ Links ]
Submitted 28 January 2022
Accepted 10 August 2022
Published 12 December 2022
# Corresponding authors: animalpark@kangwon.ac.kr


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}