










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 n.3 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i3.13
Feeding Japanese quail diets supplemented with probiotics and enzymes
Ö. Durna AydinI; G. YildizII; O. MerhanIII; S. UluferIV; K. ÖnkV; M.S. BaranVI; O. KaplanVI,#
IKafkas University, Veterinary Faculty, Animal Nutrition, and Nutritional Diseases Department, Kars, Turkey
IIAnkara University, Veterinary Faculty, Animal Nutrition, and Nutritional Diseases Department, Ankara, Turkey
IIIKafkas University, Faculty of Veterinary Medicine, Department of Biochemistry, Kars, Turkey
IVKafkas University, Vocational School of Kagizman, Department of Pharmacy Services, Kars, Turkey
VKafkas University, Veterinary Faculty, Animal Science Department, Kars, Turkey
VIDicle University, Veterinary Faculty, Animal Nutrition, and Nutritional Diseases Department, Kars, Turkey
ABSTRACT
This study assessed the effects of dietary supplementation with probiotics and enzymes on performance, blood antioxidant capacity, and cecal short-chain fatty acid concentration. Two hundred one-day-old quail chicks were randomly divided into four groups, each containing 50 chicks. All the chicks were fed with a basal diet for 35 days. The control group was fed the basal diet, and the experimental groups were fed the basal diet augmented with a commercial probiotic and enzyme supplement at 0.5 g/kg, 1 g/kg and 2 g/kg. At the end of experiment, initial live weight, final live weight, live weight gain and feed intake were affected significantly by the addition of the supplement. However, the supplement did not influence the feed conversion ratio. Increasing the levels of the probiotic and enzyme supplement Diazyme 256 in the diets caused linear and quadratic responses in malondialdehyde, glutathione, superoxide dismutase, and glutathione peroxidase level. Catalase showed a linear increase with the rise in level of the supplement. However, ceruloplasmin, albumin, total protein and globulin were not affected. Significant linear responses were observed in propionic acid, butyric acid and isocaproic acid with the increasing levels of the supplement. There were no significant differences in acetic acid, isobutyric acid, valeric acid, isovaleric acid, caproic acid, branched chain fatty acids, and short chain fatty acid concentrations for quails fed various levels of the supplement. In conclusion, the supplement Diazyme 256 containing probiotics and enzymes could be used in quail diets to improve growth performance, antioxidant capacity and intestinal health.
Keywords: antioxidant capacity, growth performance, short-chain fatty acid
Introduction
Many feed additives have been developed with biotechnology and used to improve the health of poultry. Antibiotics that are used in poultry production are produced by selected microorganisms and natural metabolites (Guo et al., 2004). The prolonged use of antibiotics as growth promotors in poultry production prevented the advance of pathogenic and beneficial microorganisms in the digestive tract (Kahraman et al., 1996). But the use of antibiotics in feed could lead to the development of resistance in pathogenic bacteria. In addition, residues in animal products became a risk to human health (Güler et al., 2019). Because of these undesirable aspects, the use of antibiotics as growth promoters in animal feed was prohibited. As a result of the ban on the use of antibiotics and other growth factor chemical substances, alternative feed additives were sought (Kaplan et al., 2018). Thus probiotics, prebiotics, enzymes, organic acids and products such as essential oils began to be used as alternatives (Pournazari et al., 2017).
The term probiotic is derived from the Latin preposition pro ('for') and the Greek word biotikos and means 'for life'. Probiotics are living microbial feed supplements that affect the host animal beneficially by improving intestinal microbial balance. They strengthen the immune system by increasing the level of antibodies in the digestive system (Fuller, 1989; Abd El-Hack et al., 2020). Probiotic bacteria have significant antioxidant abilities in vivo and in vitro (Persichetti et al., 2014).
The use of Lactobacillus acidophilus and L. casei as probiotics in broiler diets may influence growth performance positively (Angel et al., 2005; Khan et al., 2020). In recent research, the use of probiotics as feed additives in Japanese quails was observed to improve feed bioavailability, health and immune status (Kheiri et al., 2015). Supplementation of Lactobacillus spp. can manipulate fermentation in the digestive tract of broilers resulting in increased short-chain fat acid (SCFA) accumulation in the cecum and thus improve gastrointestinal tract development and prevent disease (Meimandipour et al., 2010).
Enzymes are biocatalysts that are involved in biochemical reactions and can be synthesized by living cells. The aim of adding enzymes to diets is to improve the digestion of carbohydrates by inactivating undesirable substances (Diler, 2007). Some feedstuffs are high in protein, but contain a large number of anti-nutritional factors, such as protease inhibitors, soybean protein and oligosaccharides in soybean meal, non-starch polysaccharides in corn, and phytic acid in bran, which limit their wider application in animal production (Lv et al., 2021). The use of exogenous enzymes to counteract the targeted anti-nutritional properties of dietary components and to increase growth performance in poultry is routine practice (Munir & Maqsood, 2013). Enzymes as a feed additive are much more effective in poultry than in other species owing to the rapid passage of feed through the digestive tract. They enable the digestion of non-digestible polysaccharides in the cell wall of grain, reduce intestinal viscosity caused by non-digestible polysaccharides and increase the bioavailability of phosphorus, feed efficiency and metabolic energy values (Karademir & Karademir, 2003).
In recent years, several studies evaluated the effects of probiotics and enzymes as feed additives. However, few studies researched the use of probiotics and enzymes in Japanese quail diets. Therefore, the aim of this study was to investigate the effects of Diazyme 256 in Japanese quail diets on growth performance, some blood parameters, such as glutathione (GSH), superoxide dismutase (SOD), glutathione peroxidase (GPx), catalase (CAT), malondialdehyde (MDA), ceruloplasmin, albumin, total protein, globulin, and SCFA and BCFA (branched chain fatty acid) concentrations in the cecum.
Materials and Methods
The study was approved by Kafkas University Animal Experiments Local Ethics Committee (Decision No: KAU-HAYDEK /2018-085/ 2019-138). Two hundred unsexed one-day-old Japanese quail (Coturnix coturnix japonica) chicks were used. The chicks were randomly allocated to four groups of 50. Each group was then randomly divided into five replications each containing ten chicks.
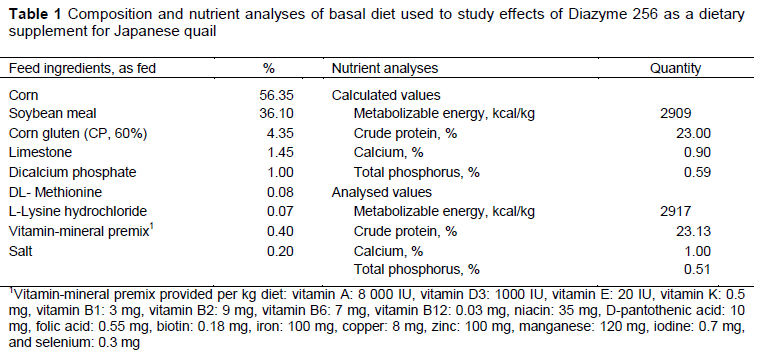
The birds were fed a diet based on corn and soybean meal (Table 1). All diets were calculated to meet the requirements of animals according to NRC (1994) standards. Nutrient analyses of the feed were performed according to methodology of the AOAC (2000). The trial lasted for 35 days.
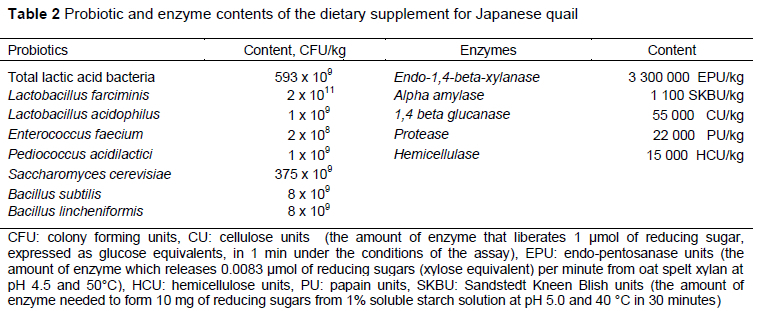
To form the experimental treatments the basal diet was augmented with various levels of a commercial supplement (Diazyme 256®, Tarimsan Chemicals A.S. Istanbul, Turkey), which contained probiotics and enzymes. The diets consisted of CON: unsupplemented basal diet, PE1: basal diet augmented with 0.5 g/kg Diazyme 256; PE2: basal diet with 1 g/kg Diazyme 256; and PE3: basal diet with 2 g/kg Diazyme 256. The probiotic and enzyme content of the supplement is shown in Table 2.
The birds were housed in cages equipped with manual feeders and automatic nipple drinkers. Water and feed were given ad libitum. The house temperature was monitored throughout the study. It was set at 32-35 °C on the first day, lowered gradually, and maintained at 22 °C for the last two weeks of the experiment. Cage dimensions were 95 χ 45 χ 22 cm. Ten quail were housed in each cage. A lighting programme was implemented in accordance with commercial conditions and provided 23 hours of light per day throughout the experiment.
Live weights of each subgroup were recorded at the beginning and end of the study. Live weight gain (LWG) was determined as the difference between these measurements. Feed intake (FI) was recorded weekly, and was determined from the difference between feed delivered and feed remaining in the feeder. Feed conversion (FCR) was calculated as the ratio between FI and LWG for each subgroup. In the fourth week and at the end of the study, blood samples were taken from the wing veins of the animals into tubes containing EDTA as an anticoagulant. After obtaining a sufficient amount, these blood samples were divided into two parts, and plasma was obtained from one of these samples. The samples were centrifuged at 3000 rpm for 15 minutes and stored at -20 oC until analysis. Superoxide dismutase, GPx and CAT antioxidant enzyme activity levels in the plasma were determined by enzyme-linked immunosorbent assay (Epoch, Agilent Technologies, Santa Clara, California, USA) using commercial kits (Cayman Chemical Company, Ann Arbor, Michigan, USA). Whole blood reduced GSH analysis was determined colorimetrically according to Beutler et al. (1963). The MDA in plasma was determined by the method of Yoshioka et al. (1979), ceruloplasmin by the method of Colombo & Ricterich (1964) and albumin and total protein levels were measured with a commercial test kit (Biolabo, Maizy, France). Globulin value was calculated by subtracting albumin from total protein (Doumas et al., 1971).
At the end of the study, the contents of the cecum were sampled and stored at -18 °C. Then frozen cecal digesta were thawed at 4 °C and diluted tenfold with double-distilled water in sterile screwcap tubes before analysis. The cecal digesta were centrifuged at 4000 rpm for 15 min at 4 °C for homogenization. The supernatant was taken into a 750 μl Eppendorf tube and mixed with 150 μl ice-cold 25% metaphosphoric acid solution. Then the tubes were kept in an ice-cold waterbath for 30 min to ensure the collapse of proteins. Subsequently, tubes were centrifuged for 10 min at 10000 rpm at + 4 °C. Supernatants were analysed with a gas chromatograph (Shimadzu GC-2010, Shimadzu Co., Kyoto, Japan) coupled with a 30 m χ 0.53 mm internal diameter column (Teknokroma TRB-FFAP, Teknokroma, Barcelona, Spain) and flame ionization detector to determine SCFA concentrations in cecal digesta. The analysis was performed according to Zhang et al. (2003). The injector-port and flame ionization detector temperatures were fixed at 250 °C. The initial temperature was held at 120 °C for 4 min after injection and then increased at 4 °C/min to 160 °C, where it was held for 4 min. Helium was used as the carrier gas. The injection volume was set at 1 μΙ and analyses were performed in duplicate.
A one-way variance analysis was run to determine the effects of treatments, and polynomial contrasts were used to determine their nature. The statistical analyses used the SPSS version 20 software (IBM Corp., Armonk, New York, USA).
Results and Discussion
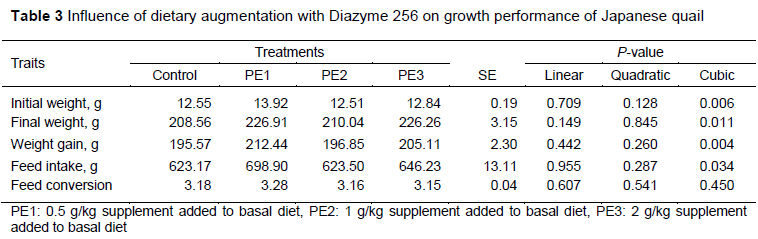
Initial weight, final weight, LWG and FI exhibited cubic responses to the level of supplement (Table 3). However, the supplement did not affect FCR.
In animals, the enzyme system develops with age. Therefore the addition of external enzymes is more effective in young animals (Demirel & Gürbüz, 1999). Probiotics help in the digestion of nutrients by working symbiotically with enzymes produced by the cells of the digestive systems of animals (Vanbelle et al., 1990). In the present study, increasing doses of Diazyme 256 were added to quail basal diets. This had a synergistic effect on digestive enzymes and nutrient digestibility. At the end of the current study, the increasing levels of Diazyme 256 in the diet affected initial LW, final LW, LWG and FI as cubic responses. Low doses (PE1) were more effective than higher doses. However, the addition of Diazyme 256 did not affect FCR. The results of many studies are compatible those of the present study (Huang, 2012). Siadati et al., 2012 found improved LWG using Lactobacillus strains at doses of 50, 100, 150, and 200 mg/kg in quail diets. Abdel-Moneim et al. (2020) reported that whereas the LWG value increased with the use of probiotics in quail diets, the FI value was not affected. Zhao et al. (2020) noted that the use of probiotic Bacillus íicheniformis H2 enhanced affected performance positively. In a study in which probiotics and enzymes were given together in broiler diets positively affected their performance (Wang et al., 2021). Similarly, Li et al. (2014) reported that the use of probiotics affected performance positively by releasing high-active protease, amylase, and lipase. In a study that used enzyme in Japanese quail diets, LWG and FCR improved, but FI was not affected (Kianfar et al., 2013). Jazi et al. (2020) reported that feeding quails with Bacillus subtilis improved growth performance, but the addition of an enzyme to broiler diets decreased LWG, FI and FCR. In a study conducted with quails, probiotics at doses of 0.1 g/kg and 0.3 g/kg did not affect initial LW, final LW, LWG, FI and FCR (Durna Aydin et al., 2021). Enzyme supplementation did not show improvement in growth performance and nutrient digestibility in broilers (Sharifi et al., 2012). When phytase and protease enzymes were added to a diet containing quinoa seeds, the growth performance of broiler chickens and intestinal bacterial ecology were affected positively (Asnaashari Amiri et al., 2021). In a study that used xylanase in a broiler diet, LW and FI were not affected (McCaffery et al., 2019). In another study, 100, 150, 200 and 250 mg/kg probiotics in quail diets improved growth performance and feed efficiency (Gao et al., 2017).The addition of probiotics to poultry diets did not affect growth performance in some studies (Hao et al., 2008). The variations among these studies could be explained by the dose, quality, and chemical composition of the probiotics and enzymes and the differences in maintenance and feeding conditions.
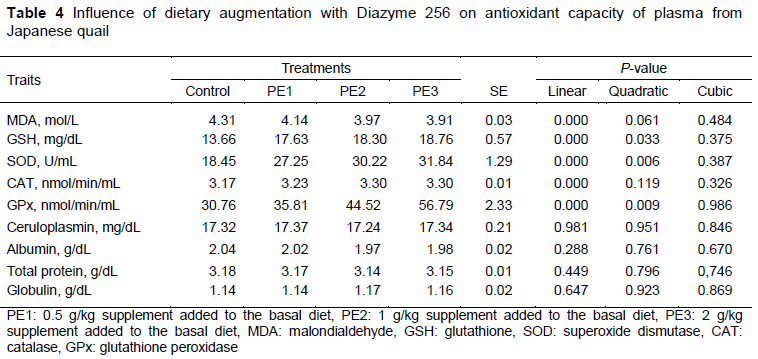
Increasing doses of Diazyme 256 in diets showed linear (P = 0.000, for all) and quadratic (P = 0.061, P = 0.033, P = 0.006 and P = 0.009, respectively) a response in MDA, GSH, SOD and GPx values. Thus the effective dose for GSH, SOD and GPx values in the current study was 0.5 g/kg. Catalase showed a linear increase with rises in the level of Diazyme 256 in diets (P= 0.000). However, ceruloplasmin, albumin, total protein and globulin were not affected. Table 4 shows the influence of the diets on antioxidant capacity.
Probiotics have their own antioxidant enzymatic systems (Landis & Tower, 2005). The culture supernatant, intact cells, and intracellular cell-free extracts of Bifidobacterium animalis cleared up hydroxyl radicals and superoxide anion in in vitro studies by improving antioxidase activities (Shen et al., 2011). In this study, plasma MDA, GSH SOD, CAT and GPx values were affected significantly by the addition of probiotic and enzyme to quail diets, but plasma ceruloplasmin, albumin, total protein and globulin were not affected statistically. The results of many studies were compatible with those of the current study. Dietary augmentation with Lactobacillus fermentum increased serum SOD, GPx and hepatic CAT and muscle SOD in pigs (Wang, et al., 2009). In another study, the addition of probiotics in various doses increased GPx activity in chicks (Aluwong et al., 2013). Addition of 0.1 g/kg and 0.3 g/kg probiotics to quail diets exhibited a linear response in MDA, GSH, SOD, CAT and GPx. However, ceruloplasmin, albumin, total protein and globulin were not affected by the addition of probiotics (Durna Aydin et al., 2021). Abdel-Moneim et al. (2020 increased B. subtilis levels in Japanese quail diets and showed a linear increase in serum total protein, albumin GSH and CAT levels, whereas there was a linear decrease in MDA value. Zhao et al. (2020) reported that the use of probiotic (Bacillus licheniformis H2 enhanced) decreased MDA concentration in the ileum and improved the activities of antioxidant enzymes in the ileum, serum, and liver. In addition, probiotics promoted antioxidant enzymatic activities (for example SOD and GSH) and total antioxidant status of the host (Wang et al., 2017). On the other hand, Jazi et al. (2020) reported that SOD and GSH-Px activities and MDA content in breast muscle were not affected by the addition of Bacilius subtilis to quail diets. The addition of xylanase and phytase to Japanese quail diets affected animals positively against transfer of stress (Mehraei Hamzekolaei et al., 2016). When probiotics and enzymes were given together in broiler diets, antioxidative capacity was affected positively (Wang et al., 2021). In an investigation of the effects of dietary supplementation with Bacillus amyloliquefaciens on antioxidant capacity and digestive enzyme activity of broiler chickens, GSH-Px and SOD activities in serum and liver, and total antioxidant capacity (T-AOC) in liver of broilers on day 42 increased linearly with rising levels of BA, whereas MDA level in serum decreased linearly (Sun et al., 2021).
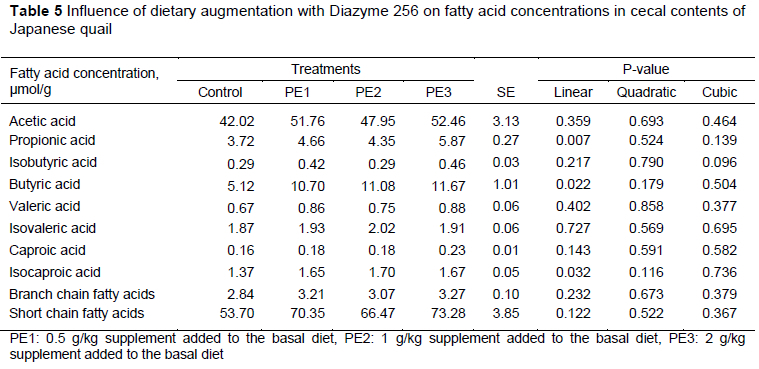
Table 5 shows the effects of the diets on some cecal SCFAs μmol/g). Propionic acid (P = 0.007), butyric acid (P = 0.022) and isocaproic acid (P = 0.032) were affected linearly with the increasing levels of Diazyme 256. There were no significant differences in acetic acid, isobutyric acid, valeric acid, isovaleric acid, caproic acid, and BCFA and SCFA concentrations for quails fed the various levels of Diazyme 256.
Significant numbers of studies focused on intestinal microbiota in individual health status. Probiotics improved metabolic diseases such as obesity and diabetes mellitus by modulating the intestinal microorganisms (Wang et al., 2015). Short chain fatty acids are formed by bacterial fermentation in the cecum. They stimulate cell growth, improve intestinal integrity, reduce the digestive tract pH, and prevent the growth of pathogenic microorganisms (Knudsen et al., 2012; Abd El-Hack et al., 2020). Increased SCFA concentrations have beneficial effects on energy, metabolism, microflora and immune responses (Pan & Yu, 2014). Cereal grains may affect cecal microflora and SCFA production. Poultry fed maize-based diets generally consumed lower concentrations of non-starch polysaccharide substrates, which might have affected the development of cecum microflora and fermentative capacity. Resistant starch concentrations indicated the presence of high amylose in cereal grains. The viscous structure of these polysaccharides has a direct effect on nutrient absorption, modifies intestinal physiological function and interacts with intestinal microflora. Increased amounts of NSP in the digestive system affect intestinal microflora negatively. Cecal volatile fatty acid concentration was increased significantly with enzyme supplementation in poultry (Choct et al. 1996). Therefore, adding enzymes to the diet could be advantageous for gut health, especially for young animals. In the current study, significant linear responses in propionic acid, butyric acid and isocaproic acid were observed with increasing levels of Diazyme 256. However, there were no significant differences in concentrations of acetic acid, isobutyric acid, valeric acid, isovaleric acid, caproic acid, BCFAs and SCFAs. Murugesan et al. (2014) observed that cecal butyrate increased with the addition of protease, phytase and Bacillus spp to broiler diets. Durna Aydin et al. (2021) observed no significant differences in acetic acid, propionic acid, butyric acid, isobutyric acid, valeric acid, isovaleric acid and BCFA concentrations in quails fed with various levels of probiotic. However, they did find significant linear responses in ascetic acid, isocaproic acid and SCFA with the addition of 0.1 g/kg and 0.3 g/kg probiotic to quail diet, and a significant quadratic response in caproic acid was also seen. In a study in which various doses (0.5 g/10 l and 1 g/10 l) of probiotics were added to quail drinking water, acetic acid, propionic acid, isobutyric acid, butyric acid, isovaleric acid, valeric acid, caproic acid, total SCFA and BCFA values of the cecum contents were not statistically significant (Durna Aydin & Yildiz, 2020). The use of xylanase in broiler diets did not affect cecal acetic acid, but improved propionic, isobutyric and isovaleric acid concentrations in the cecum (McCaffery et al., 2019). Weng et al. (2019) reported that in a study of the correlation of variables on metabolic and microbiota with diet in intestinal inflammation, the value of isocaproic acid decreased. This showed that pH acidity in the intestine was impaired and pathogenic microorganisms were active in inflammation. In the light of these studies, the increase in cecum SCFAs could be interpreted as having a positive effect on gut health. On the other hand, cecal fermentation was influenced by the use of Diazyme 256. Morever, the addition of Baciííus íicheniformis has been shown to decrease cecal concentrations of propionic, butyric, n-butyric and n-valeric acids in broilers (Novak et al., 2011). Thus, the use of probiotics and enzymes has a positive effect on cecum volatile fatty acids, and seemed to improve intestinal health. Therefore, the use of probiotic and enzyme complexes could be an alternative to antibiotics.
Conclusion
The diets augmented with Diazyme 256 increased GSH, SOD, GPx and CAT, indicating a protective effect against oxidative damage. Moreover, the significant increase in propionic acid, butyric acid and isocaproic acid indicated that these diets could improve intestinal health in quail. Thus, Diazyme 256 could be used safely to avoid oxidative stress and maintain gut health in quail.
Acknowledgement
The authors extend their thanks to Yusuf Kaya from Tarimsan Inc., who provided probiotics and enzymes.
Authors' contributions
ÖDA, GY, KÖ and SUB (ORCIDs: 0000-0003-4532-6795; 0000-0002- 1003- 9254; 0000-0002- 5618- 2988; 00000001-9533-0165) executed the experiment; OK (ORCID: 0000-0001-6143-8987) completed the manuscript; MSB (ORCID: 0000-0002-7297-8556) helped in preparing the manuscript. GY, ÖDA and OM (ORCiD: 0000-0002-3399-0667) were in charge of laboratory analyses. OK was responsible for supervisiing and writing of the manuscript.
Conflict of interest
The authors declare that they have no conflicts of interest.
References
Abd El-Hack, M.E., El-Saadony, M.T., Shafi, M.E., Qattan, S.Y.A., Batiha, G.E., Khafaga, A.F., Abdel-Moneim A.E. & Alagawany, M., 2020. Probiotics in poultry feed: A comprehensive review. J. Anim. Physiol. Anim. Nutr. 104(6),1835-1850. DOI: 10.1111/jpn.13454 [ Links ]
Abdel-Moneim A.E., Selim D.A., Basuony H.A., Sabic E.M., Saleh A.A. & Ebeid, T.A., 2020. Effect of dietary supplementation of Bacillus subtilis spores on growth performance, oxidative status, and digestive enzyme activities in Japanese quail birds. Trop. Anim. Health. Prod. 52, 671-680. DOI: 10.1007/s11250-019-02055-1 [ Links ]
Aluwong T., Kawu, M., Raji, M., Dzenda, T., Govwang, F., Sinkalu, V. & Ayo, J., 2013. Effect of yeast probiotic on growth, antioxidant enzyme activities and malondialdehyde concentration of broiler chickens. Antioxidants 2, 326339. DOI: 10.3390/antiox2040326 [ Links ]
Angel, R., Dalloul, A. & Doerr, J., 2005. Performance of broiler chickens fed diets supplemented with a direct-fed microbial. Poult. Sci. 84, 1222-1231. DOI: 10.1093/ps/84.8.1222 [ Links ]
AOAC, 2000. Official methods of analysis of the Association of Official Analytical Chemists. 17th ed. AOAC International, Maryland, USA. [ Links ]
Asnaashari Amiri, M.Y., Jafari, M. A. & Irani, M., 2021. Growth performance, internal organ traits, intestinal morphology, and microbial population of broiler chickens fed quinoa seed-based diets with phytase or protease supplements and their combination. Trop. Anim. Health. Prod. 53, 535-543. DOI: 10.1007/s11250-021-02980-0 [ Links ]
Beutler, E., Duron O. & Kelly, B.M., 1963. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 61, 882-888. [ Links ]
Choct, M., Hughes R.J, Wang, J., Bedford M.R., Morgan A.J. & Annison G., 1996. Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of nonstarch polysaccharides in chickens. Br. Poult. Sci. 37, 609621. DOI: 10.1080/00071669608417891 [ Links ]
Colombo, J.P. & Richterich, R., 1964. Zur bestimmung des caeruloplasmin im plasma (On the determination of ceruloplasmin in plasma). Schweiz Med. Wochenschr. 23, 715-720. [ Links ]
Demirel, R. & Gürbüz, Y., 1999. Karma yemlerde enzim kullanimi, s.489-495. V. Poultry Yutav'99 Uluslararasi Tavukçuluk Fuari ve Konferansi 3-6 Haziran Bildiriler Kitabi Istanbul; 1999. [ Links ]
Diler, A., 2007. Probiyotik, enzim kombinasyonunun esmer irki buzagilarda yemden yararlanma ve büyüme performansi üzerine etkileri. Yüksek lisans tezi. Atatürk Üniversitesi Fen Bilimleri Enstitüsü, Erzurum. [ Links ]
Doumas, B.T., Watson, W.A. & Biggs, H.G., 1971. Albumin standards and the measurement serum albumin with bromcresol green. Clinica Chimica Acta. 31(1), 87-96. DOI: 10.1016/0009-8981(71)90365-2 [ Links ]
Durna Aydin, Ö. & Yildiz, G., 2020. Effect of drinking water supplementation of probiotic on cecal short-chain fatty acid concentrations in quail breeders (Coturnix coturnixjaponica). J. Dicle Univ. Faculty of Vet. Med. 13(1), 61-64. [ Links ]
Durna Aydin, Ö., Yildiz, G. & Merhan, O., 2021. Effects of probiotic (Lactobacillus farciminis) supplementation in quail (Coturnix coturnix japonica) rations on growth performance, blood antioxidant capacity and cecal some short-chain fatty acid concentrations. Kafkas Univ. Vet. Fak. Derg. 27 (1), 15-20. DOI: 10.9775/kvfd.2020.24541 [ Links ]
Fuller, R., 1989. A review: Probiotics in man and animals. J. Appl. Bacteriol. 66(5), 365-378. [ Links ]
Gao, Z., Wu, H., Shi, L., Zhang, X., Sheng, R., Yin, F. & Gooneratne R., 2017. Study of Bacillus subtilis on growth performance, nutrition metabolism and intestinal microflora of 1 to 42 d broiler chickens. Anim. Nutr. stegen, M.W.A., 2004. Effects of mushroom and herb polysaccharides, as alternatives for an antibiotic, on growth performance of broilers. Br. Poult. Sci. 45, 684-694. [ Links ]
Güler, A., Kaplan, O. & Bozkaya, F., 2019. Effects of probiotics added to some roughages on in vitro organic matter digestibility and methane production. Harran Univ. Vet. Fak. Derg. 8 (1), 93-98. [ Links ]
Hao, S.H., Tong, J.M., Yang, R.F. & Sa, R.N., 2008. Effect of bacillus licheniformis o broilesr aged 0-3 weeks. Journal of Northwest AF University (Nat. Sci. Ed.), 36(8), 20-24, 30. [ Links ]
Huang, J.H., 2012. Effect of Bacillus subtilis on growth performance of broilers chicken. Heilongjiang Anim. Sci. Vet. Med. 2, 78-79. [ Links ]
Jazi, V., Farahi, M., Khajali, F., Abousaad, S., Ferket, P. & Assadi Soumeh, E., 2020. Effect of dietary supplementation of whey powder and Bacillus subtilis on growth performance, gut and hepatic function, and muscle antioxidant capacity of Japanese quail. J. Anim. Physiol. Anim. Nutr. 104, 886-897. DOI: 10.1111/jpn.13323 [ Links ]
Kahraman, R., Alp, M., Kocabagli, N., Irmak, G. & Çenel, H.S., 1996. The effects of fastrack and sodium bicarbonate on performance of broilers. Turk J. Vet. Anim. Sci. 20, 383-386. [ Links ]
Kaplan, O., Avci, M., Denek, N., Baran, M.S., Hüseyin Nursoy, H. & Bozkaya, F., 2018. Influence of humic acid addition to drinking water on laying performance and egg quality in Japanese quails. Indian J. Anim. Res. 52(9), 2018, 1309-1312. DOI: 10.18805/ijar.B-874 [ Links ]
Karademir, G. &Karademir, B., 2003. Yem katki maddesi olarak kullanilan biyoteknolojik ürünler. Lalahan Hay. Araçt. Enst. Derg, 43(1), 61-74. [ Links ]
Khan, S., Moore, R.J., Stanley, D. & Chousalkar, K.K., 2020. The gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl Environ Microbiol. Jun 17, 86(13):e00600-20. DOI: 10.1128/AEM.00600-20. [ Links ]
Kheiri, F., Rostami, M., Hajiabadi, S., 2015. The effects of Protexin probiotic and chicoridin supplementation on performance and some hematological parameters in Japanese quail (Coturnix coturnixjaponica). Adv. Agric. 4 (10), 115-121. DOI: 10.14196/aa.v4i10.2012 [ Links ]
Kianfar, R., Moravej, H., Shivazad, M. & Taghinejad-Roudbaneh, M., 2013. Effect of enzyme addition, germination, and fermentation on the nutritive value of barley for growing Japanese quails. J. Anim. Feed Sci. 22, 165-171. [ Links ]
Knudsen, K.E.B., Hedemann, M.S. & Laerke, H.N., 2012. The role of carbohydrates in intestinal health of pigs. Anim. Feed Sci. Technol. 173, 41-53. [ Links ]
Landis, G.N. &Tower, J., 2005. Superoxide dismutase evolution and life span regulation. Mech. Ageing Dev. 126, 365-379. DOI: 10.1016/j.mad.2004.08.012 [ Links ]
Li, W., Bai, J, Li, Y., Qin, Y. & Yu, D., 2014. Effects of Bacillus subtilis on meat quality, nutrient digestibility and scrum biochemical index of broilers. Chin J. Vet. Sci. Med. 34, 1682-1685. [ Links ]
Lv, J., Guo, L., Chen, B., Hao, K., Ma, H., Liu, Y. & Min, Y., 2021. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poult. Sci. 98 (7), 2866-2879. DOI:10.1016/j.psj.2021.101570 [ Links ]
McCaffery, K.W., Bedford, M.R., Kerr, B.J. & Doizer, W., 2019. Effects of cereal grain source and supplemental xylanase concentrations on broiler growth performance and cecal volatile fatty acid concentrations from 1 to 40 d of age. Poult. Sci. 98(7), 2866-2879. Doi: 10.3382/ps/pez032 [ Links ]
Mehraei Hamzekolaei, M.H., Zamani Moghaddam, A.K., Tohidifar, S.S., Dehghani Samani, A. & Heydari, A., 2016. The effects of transportation stress on Japanese quail (Coturnix Coturnix japonica) fed corn-based diet in comparison with wheat-based diet supplemented with xylanase and phytase. J. Anim. Physiol. Anim. Nutr. 100(4), 618-622. DOI: 10.1111/jpn.12398 [ Links ]
Meimandipour, A., Shuhaimi, M., Soleimani, A.F., Azhar, K., Hair-Bejo, M., Kabeir, B.M., Javanmard, A., Muhammad-Anas, O. & Yazid, A.M., 2010. Selected microbial groups and short-chain fatty acids profile in a simulated chicken cecum supplemented with two strains of Lactobacillus. Poult. Sci. 89 (3), 470-476. DOI: 10.3382/ps.2009-00495 [ Links ]
Munir, K. & Maqsood, S., 2013. A review on role of exogenous enzyme supplementation in poultry production. Emir. J. Food Agric. 25(1), 66-80. DOI: 10.9755/ejfa.v25i1.9138 [ Links ]
Murugesan, G.R., Romero, L.F. & Persia, M.E., 2014. Effects of protease, phytase and a Bacillus sp. direct-fed microbial on nutrient and energy digestibility, ileal brush border digestive enzyme activity and cecal short-chain fatty acid concentration in broiler chickens. PLoS One 9(7), e101888. [ Links ]
Novak, R., Bogovic Matijasic, B., Tercic, D., Cervek, M., Gorjanc, G., Holkman, A., Levart, A. & Rogelj, I., 2011. Effects of two probiotic additives containing Bacillus spores on carcass characteristics, blood lipids and cecal volatile fatty acids in meat type chickens, J. Anim. Physiol. Anim. Nutr. 95(4), 424-433. [ Links ]
NRC, 1994. Nutrient requirements of poultry. Nineth rev. ed. National Academy Press, Washington DC. Pp. 34-45. [ Links ]
Pan, D. & Yu, Z., 2014. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 5, 108-119. [ Links ]
Persichetti, E., De Michele, A., Codini, M. & Traina, G., 2014. Antioxidative capacity of Lactobacillus fermentum LF31 evaluated in vitro by oxygen radical absorbance capacity assay. Nutr. 30(7-8), 936-938, DOI: 10.1016/J.NUT.2013.12.009 [ Links ]
Pournazari M., Qotbi A.A., Seidavi A. & Corazzin M., 2017. Prebiotics, probiotics and thyme (Thymus vulgaris) for broilers: Performance, carcass traits and blood variables. Rev. Colombiana de Ciencias Pecuar. 30, 3-10. [ Links ]
Sharifi, S.D., Shariatmadari, F. & Yaghobfar, A., 2012. Effects of inclusion of hull-less barley and enzyme supplementation of broiler diets on growth performance, nutrient digestion and dietary metabolisable energy content. J. Cent. Eur. Agric. 13(1), 193-207. Doi: 10.5513/JCEA01/13.1.1035 [ Links ]
Shen, Q., Shang, N. & Li, P., 2011. In vitro and in vivo antioxidant activity of Bifidobacterium animalis 01 isolated from centenarians. Curr. Microbiol. 62, 1097-1103. DOI: 10.1007/s00284-010-9827-7 [ Links ]
Siadati, Tan L., Yuan, D. & Chen, Y.B., 2012. Differ bacillus probiotics affect growth perform ammon emission excreta broilers, Chin J Anim Sci. 24(5), 877-85. [ Links ]
Sun, Y.,Zhang, Y., Liu, M., Li, J., Lai, W., Geng, S., Yuan, T., Liu, Y., Di, Y., Zhang, W. & Zhang, L., 2021. Effects of dietary Bacillus amyloliquefaciens CECT 5940 supplementation on growth performance, antioxidant status, immunity, and digestive enzyme activityof broilers fed corn-wheat-soybean meal diets. Poult. Sci. 98 (7), 28662879. DOI: 10.1016/j.psj.2021.101585 [ Links ]
Vanbelle, N., Teller, E. & Focant, M., 1990. Probiotics in animal nutrition. A Review. Arch Tierernaehr. 40(7), 543-567. [ Links ]
Wang, A.N., Yi, X.W., Yu, H.F., Dong, B. & Qiosa, S.Y., 2009. Free radical scavenging activity of Lactobacillus fermentum in vitro and its antioxidative effect on growing-finishing pigs. J. Appl. Microbiol. 107, 1140-1148. DOI: 10.1111/j.1365 2672.2009.04294.x [ Links ]
Wang, J., Tang, H., Zhang, C., Zhao, Y., Derrien, M., Rocher, E., Van-Hylckama Vlieg, J. ET., Strissel, K., Zhao, L., Obin, M. & Shen, J., 2015. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J. 9, 1-15. DOI: 10.1038/ismej.2014.99 [ Links ]
Wang, Y., Wu, Y., Wang, Y., Xu, H., Mei, X., Yu, D., Wang, Y. & Li, W., 2017. Antioxidant properties of probiotic bacteria, Nutr. 9(5), 521. DOI: 10.3390/nu9050521 [ Links ]
Wang, Y., Heng, C., Zhou, X., Cao, G., Jiang, L., Wang, J., Li, K., Wang, D. & Zhan, X., 2021. Supplemental Bacillus subtilis DSM 29784 and enzymes, alone or in combination, as alternatives for antibiotics to improve growth performance, digestive enzyme activity, anti-oxidative status, immune response and the intestinal barrier of broiler chickens. Br J Nutr. 14, 125(5):494-507. DOI: 10.1017/S0007114520002755 [ Links ]
Weng, Y.L., Gan, H.Y., Li, X., Huang, Y., Li, Z.C., Deng, H.M., Chen, S.Z., Zhou, Y., Wang, L.S., Han, Y.P.,Tan, Y.F., Song, Y.J., Du, Z.M., Liu, Y.Y., Wang, Y., Qin, N., Bai, Y., Yang, R.F., Bi, Y.J. & Zhi, F.C., 2019. Correlation of diet, microbiota and metabolite networks in inflammatory bowel disease. J. Dig. Dis. 20, 447-459. DOI: 10.1111/1751-2980.12795 [ Links ]
Wyatt, C.L. & Goodman, T., 1993. Utilization of feed enzymes in laying hen rations. J. Appl. Poultry Res..2, 68-74 [ Links ]
Yoshioka, T., Kawada, K., Shimada, T. & Mori, M., 1979. Lipid peroxidation in maternal and cord blood and protective mechanism against activated-oxygen toxicity in the blood. Am. J. Obstet. Gynecol. 135, 372-376. [ Links ]
Zhang, W.F., Li, D.F., Lu, W.Q. & Yi, G.F., 2003. Effects of isomalto-oligosaccharides on broiler performance and intestinal microflora. Poult. Sci. 82(4), 657-663. [ Links ]
Zhao, Y., Zeng, D., Wang, H., Qing, X., Sun, N., Xin, J., Luo, M., Khalique, A., Pan, K., Shu, G., Jing, B. & Ni, X, 2020. Dietary probiotic Bacillus licheniformis H2 enhanced growth performance, morphology of small intestine and liver, and antioxidant capacity of broiler chickens against Clostridium perfringens-induced subclinical necrotic enteritis. Probiotics Antimicrob. Proteins 12(3), 883-895. DOI: 10.1007/s12602-019-09597-8 [ Links ]
Submitted 1 February 2022
Accepted 8 April 2022
Published 24 June 2022
# Corresponding author: okaplan61@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}