










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 no.3 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i3.12
Horn growth characteristics of sable antelope (Hippotragusnigerniger) in South Africa
C.A. ShepstoneI, #; P.C. LuboutII; J.H.C. Van ZylI; L.C. HoffmanI, III; G.F. JordaanI
IDepartment of Animal Sciences, Stellenbosch University, Private Bag X1, Matieland, Stellenbosch, 7602, South Africa
IIDepartment of Animal Science, Faculty of Natural and Agricultural Science Mahikeng Campus, North-West University, Mafikeng 2735, South Africa
IIICentre for Nutrition and Food Sciences, Queensland Alliance for Agriculture and Food Innovation (QAAFI), The University of Queensland, Australia
ABSTRACT
A study of horn growth characteristics of sable antelope in South Africa investigated the environmental effects of sex, calving year and season on horn development for animals up to 50 months old. Horn growth characteristics were assessed in the age categories of 0 to 15, 15.1 to 36, and 36.1 to 50 months. The growth rate (cm/day) of horn length differed significantly between male and female sable antelope in all age categories. The horns of males grew by 0.89 cm/day and females by 0.68 cm/day between 0 and 15 months. Between 15 and 36 months, horn growth was 0.079 cm/day for males and 0.042 cm/day for females. Between 36 and 50 months horn growth was 0.044 cm/day for males and 0.015 cm/day for females. The base circumference differed significantly between the sexes only between 0 and 15 months, when it was 0.026 cm for males and 0.014 cm for females. The number of horn rings did not differ between sexes. Thus, there was rapid initial growth for both males and females, with the horn growth of females slowing noticeably once they reached sexual maturity. Supplemental feeding regimes introduced on most farms affected horn growth traits positively, resulting in longer horns at maturity.
Keywords: annulations, basal circumference, horn length, vegetation, wildlife
Introduction
The sable antelope has four recognised subspecies in Africa (Matthee & Robinson, 1999). Historically the South African population (Hippotragus niger niger) was distributed in the Magaliesberg, west of Pretoria, and at Pilgrims Rest, Waterberg, Letaba and the Kruger National Park on the eastern side of South Africa (Basson, 1989; Du Toit, 1992). The Gravelotte/Letaba population, excluding that in Kruger National Park, declined from 15 000 to 20 000 in the 1930s to about 400 animals in 2000. The population in the Letaba/Gravelotte area was deemed healthy in the early 1900s because few people lived in the area because malaria was endemic (Du Toit, 1992; Warren, 2021). But the successful eradication of malaria led to livestock and crop farmers moving into the region. Therefore the sable antelope population declined because of increased livestock and crop farming, decreased natural habitat, competition with other species for water, and because the prevention of natural veld fires resulted in an increased tick burden and tick-borne diseases (Du Toit, 1992; Rabie, 2011). The reduced population size produced lower levels of genetic variation and increased inbreeding, which led to higher levels of calf mortality (± 20%) (Warren, 2021).
The Game Theft Act, No. 105 of 1991, allowed private ownership of game (Juta, 2011), so herds of sable antelope were established on game ranches. With few unrelated animals being available in South Africa, sable antelope were imported from Malawi, Zimbabwe and Zambia (Kriek, 2017). Game farmers who bred sable antelope in controlled environments saved the species from local extinction, and increased genetic variation and reduced inbreeding. Total income from game sales in 2011 was about R4 million in 2012, which increased to more than R5 billion (Saayman et al., 2018). This increase in value resulted in more farmers entering the game breeding sector (Cloete et al., 2015). From 2012 to 2013, these superior breeding animals were well cared for to ensure calf survival and longer horns. Therefore, animal husbandry practices were adapted to include strategic feeding, and pedigree and phenotype records as an integral part of sable antelope management (Josling et al., 2019).
With the advent of sable antelope ranching with the focus on horn quality, questions arose over the efficacy of genetic selection and the influence of nutrition on horn growth. Thus, the aim of this research was to investigate the growth of horn characteristics of the sable antelope with particular reference to sex and the environmental effects.
Material and Methods
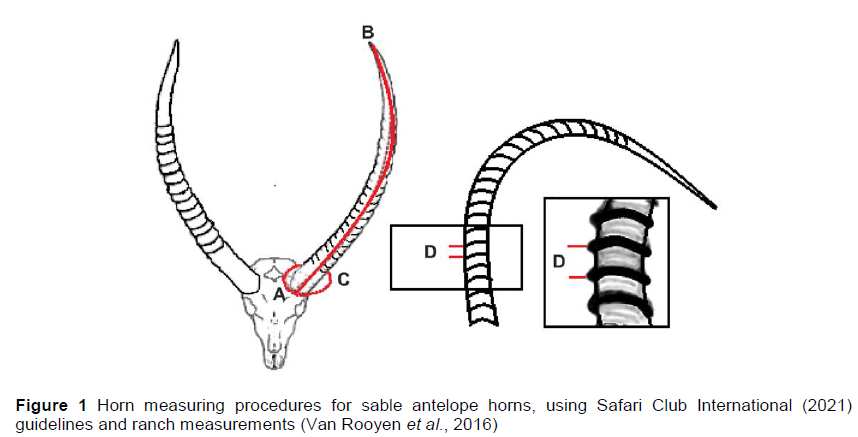
Data were provided by ranchers from routine management practices according to domestic livestock methods and principles (see for example Tucker et al., 2020). The game ranches represented the savannah-sour, savannah-sweet, rooigras-sweet and bushveld-sweet vegetation types. The data were collected from individual animals that were darted with a tranquilizer under the supervision of a wildlife veterinarian. Because sable antelope were scarce and expensive, darting was kept to a minimum to ensure least stress. The horns were measured with a measuring tape by the game rancher or wildlife veterinarian. Horn measurements were recorded using system standards set by Safari Club International for horn length (HL), basal circumference (BC), and the number of rings (HR). Horn length was measured from the base along the front length of the horn to the horn tip without pressing the tape down between the annuli. Basal circumference was measured at the base of the horn where it joined the skull. Horn ring count included only those that encircled the whole horn along its entire length (Josling et al., 2019).
Traits analysed in this study were HL, HR, and BC (Figure 1). Because some horns were damaged, only the longer horn or larger measurement was used in the analysis. The dataset was edited to include only those animals born after 2010 and younger than 1500 days (4 years). A total of 3343 observations were recorded from 2011 to 2018. These data were then edited to eliminate outliers and animals with incomplete data. The final data set included 2265 records of HL, BC, and HR for 1599 individuals.
The birth year was recorded for each animal. Age at the time of measurement was categorized as 0 to 15 months, 15.1 to 36 months, and 36.1 to 50 months. The gender of all animals with records in the data was known and males and females were present in each of the age categories. Mature animals, that is, those older than 50 months, were not included in this study. For females that had calved, the year and season of calving were recorded, and calving season was divided into summer (green) (September to April) and winter (dry) (May to August). Vegetation type was included in the statistical analyses. In 2011/2012, many ranchers had just begun to breed with sable and data were recorded only for selected animals. Because of the high costs and physical risks associated with darting, the ranchers were cautious about darting animals merely for measurements. Thus, only those animals that were moved or purchased were measured.
The data were analysed for analysis of variance with SAS for Windows version 9.4 (SAS Institute Inc., Cary, North Carolina, USA) and XLSTAT 2021.2 (Addinsoft Inc., New York, USA). Each age category was analysed separately to determine the effects of sex, calving year, calving season and interactions among these effects. Age category-specific regressions on age (in days) were calculated for each dependent variable. Then separate regressions of HL, BC and HR on age over the whole 50-month period were recorded for each type of vegetation, calving season and calving year category. Where multiple readings were available per individual, care was taken to ensure that only one data point was used per category. Because of the unbalanced nature of the data in this study, Type III sum of squares was used for all significance tests. Bonferroni post hoc tests were used to detect differences among the least squares means.
Results and Discussion
Grobler (1980a, b) studied a herd of sable antelope in Matobo National Park in Zimbabwe between 1971 and 1978. Horns were collected from animals at various ages, and horn sections were dissected and studied under a microscope to see how the tissue differed as the horns grew over time. This helped to describe the horn growth phases. The deciduous horn grows from birth until the first ring is produced. This growth is faster than the secondary permanent horn growth that occurs subsequently (Grobler, 1979, 1980b). Both male and female horns grow rapidly through the first 12 months after birth. Horn growth occurs until the animals are about 4 years old. This information was the basis for partitioning age into the categories that were used in this study, namely 0 to 15, 15.1 to 36, and 36.1 to 50 months.
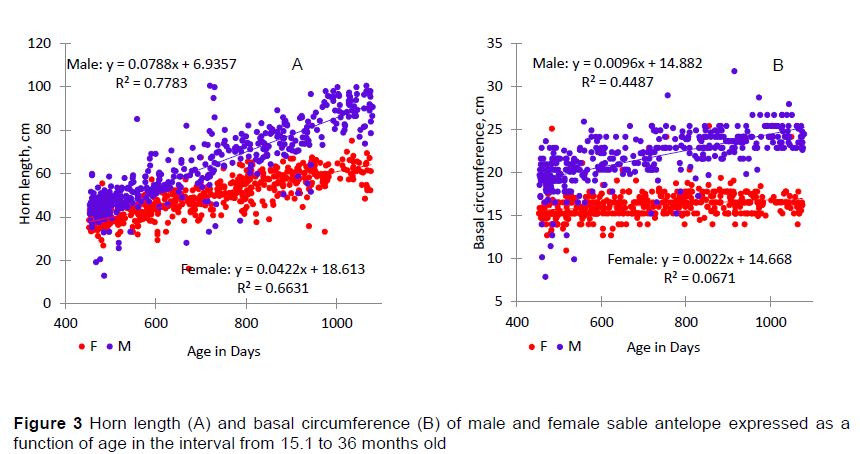
Departures from linearity in HL, HR and BC over time were not noted, but the variation among animals increased as they aged. In the first three years or so of life, the growth of a sable antelope is centred on physical and physiological trait development, when the deciduous and primary horn growth ensues. Historical data (Grobler, 1980b) showed that the horn growth of sable antelope males started to plateau at about 30 months, whereas horn growth in females started to level out about 100 days earlier. From 900 days onwards, the males in this study continued to grow through about 1400 days. Interestingly this plateau at about 1400 days coincided with the blackening of the coat in bulls (Warren, 2021). Improved nutrition possibly had a positive effect on testosterone production and growth, resulting in the 'flattening of the curve' to about 500 days later. Female animals showed a noticeable change from 800 days onwards in which a marked reduction in HL was also seen, coinciding with the time that females would be expected to commence reproducing.
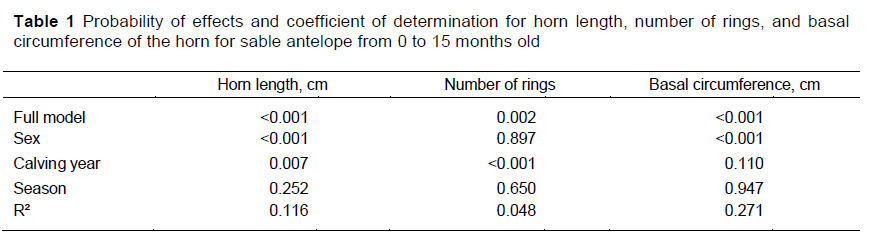
The combined effects of sex, season of birth and calving year explained relatively small amounts of the variation in HL, HR and BC for sable antelope that were 15 months old or younger, indicating substantial variability in these traits even after accounting for the effects that were included in the model (Table 1). The sex of the animal had the most prominent influence on horn development in young sable, particularly for HL (P <0.001) and BC (P <0.001). Significant effects of calving year were also noted for HL and HR. No differences in HL, HR and BC were found to have resulted from being born in summer or winter (P >0.20). Interactions among the effects of sex, season of birth and calving year explained less than 0.3% of the variation in any of these traits and were therefore deemed negligible.
Least-squares means for factors that had been hypothesized to affect HL, HR and BC of sable antelope are presented in Table 2. These young animals showed significant differences in horn growth, with male horns being almost 5 cm longer and 2.8 cm greater in circumference than those of females (15.2% and 17.5%, respectively). Because game ranchers focused on horn growth, some began to supply a nutritionally balanced diet from 2012/2013 onwards, which might explain the increased HL from 2013 to 2018. The number of HR decreased between 2013 and 2018. However, no corresponding difference in BC was detected over time. The decrease in HR over time also alluded to improved environmental conditions. Season of birth had no effect on HL, HR, and BC. Grobler (1980a) published results for sable antelope in the Matopos in Zimbabwe, which are supported by the observations in the current study. Between 6 and 12 months old (N = 13) male and female sable antelope in Zimbabwe had HL of 28.4 ± 3.0 cm and 27.5 ± 2.0 cm, respectively. Grobler (1980a) also stated that the first annulus (ring) appeared at about 12 months old. In the current study, 84 of the observations had zero rings in this age category.
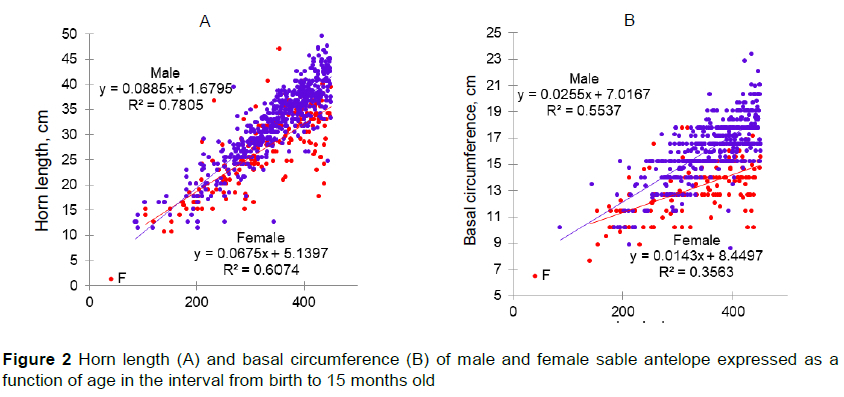
Improved nutrition might increase HL and distance between rings. This could be attributed to the increased supply of nutrients having a positive effect on steroid hormone production and increasing horn growth (Ruiz et al., 2010). Possibly this effect would benefit male animals more than female animals, with this divergence increasing with age (Figure 2A, 2B). A similar effect could be seen in the growth of young livestock, where males have heavier weaning weights than their female contemporaries (Raphaka & Dzama, 2009; Daza et al., 2014).
The amount of variation in HL and BC explained by the model increased markedly from 15.1 months to 36 months old compared with the younger animals (Table 3). The effects of sex and calving year on HL, HR and BC were highly significant for the 15.1 to 36-month-old antelope, but, as in the younger group, no effect of calving season was detected for any of the horn growth traits. Interactions between calving year and sex was significant (P =0.04), but increased R2 by only about 1% compared with the main effects and thus was not considered further.
When the sable antelope were between 15.1 and 36 months old, males had significantly longer and thicker horns than females (Table 4). This might be expected because females devote nutrients to reproductive functions at this time, redirecting them to initiating and maintaining pregnancy and to uterine involution and lactation. All of these reproductive functions take nutrients away from horn growth. The shorter HL for animals in the 2017 calving year was thought to be the result of the data structure in which the 46 males had an average age of 22 months. In other years, the distribution of animals between 15.1 and 36 months was more uniform. At this time the longer HL and BC in males were expected because they produce more testosterone naturally, especially with optimal nutrition (Ruiz et al., 2010). Testosterone is responsible for the development of male secondary sex characteristics such as larger body size, larger and stronger neck and horn growth (Sempéré & Boissin, 1981), and darkening of the coat (West & Packer, 2002; Warren, 2021). Similar development of secondary sex characteristics is seen in livestock (Daza et al., 2014)
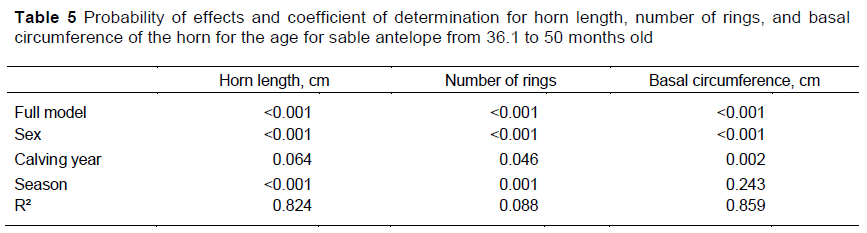
The effects of sex, and the year and season of birth explained large percentages of the variation in HL and BC (Table 5). These effects had a much smaller influence on HR. That being said, the effects of sex, and the year and season of birth were all significant or nearly so for HL and HR, whereas only the effects of sex and year of birth were significant for BC. Although all the interactions between calving year, sex and season in which the animal was born in total were significant (P <0.05), their combined contribution to R2 was 1.4% and these interactions were deemed negligible.
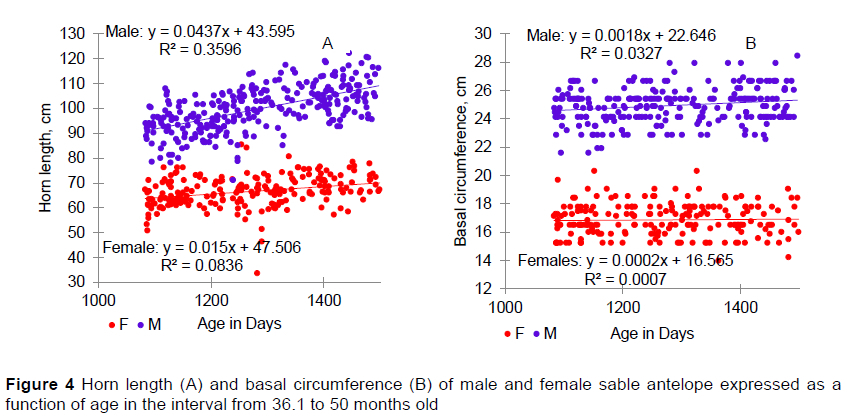
Differentiation between males and females continued in the 36.1 to 50 month category, with males having longer horns with greater BC and more HR than females (Table 6). But females were underrepresented in this age category. They were not measured because of danger to themselves and the foetus during darting. Most of the measurements for older females were taken when they were being sold. Although they were significant, or nearly so, in the analysis of variance, the effects of calving year were detectable only with the mean separation procedure for BC, and even these effects were quite small. Unlike younger ages, animals born in winter had longer horns with more rings than those born in summer. However, these seasonal differences were deemed of little practical importance.
The differences in horn growth between male and female sable antelope had become obvious by the time they were 36 months old (Figure 4). Horn growth by both male and female sable antelope slowed when they were between 36.1 and 50 months old compared with younger stages and the BC of the horns remained mostly static. Females would be expected to have 6.3 cm longer horns at the end of the period compared with the beginning, whereas the HL of males would be expected to increase by 18.4 cm. These reductions in horn growth might be attributed to the general phenomenon of maturation. Males with large horns generally have greater reproductive success (Willisch et al., 2015; Newbolt et al., 2017), which might result from advantages in competing for mates (Lincoln, 1992). Thus, males might partition more nutrients to horn growth than females at the same chronological age.
The three main factors that determine the size of antlers or horns are genetics, nutrition and age. Even with the best genetics, trophy-sized antlers and horns are not guaranteed and good range or veld is required. Unless there has been a severe winter or drought, good quality range or veld with a diversity of palatable forage species would supply the nutrients horns and antlers need for maximum growth (Knight, 2008). An adequate supply of nutrients results in more naturally produced testosterone (Ruiz et al., 2010). Testosterone plays a critical role in developing male characteristics, as shown in adult roe deer, where increased plasma concentrations of testosterone were correlated positively with the mineralization of the antlers (Sempéré & Boissin, 1981). The darkening of the coat in males is testosterone driven, as seen in the darkening of a male mane as the lion ages (West & Packer, 2002). Similarly, a jet black male sable antelope was castrated at 4.2 months old, reducing his ability to produce testosterone and within six months this bull went from pitch black to a red calf-like colour (Warren, 2021). In the present study, only characteristics of horn growth were recorded and it was assumed that game ranch managers would provide adequate nutrition to their animals because it was in their financial interest to do so.
Conclusion
There were significant differences in horn growth trait measurements between male and female sable antelope. Male and female sable antelope were most similar in the first year, with sexual dimorphism becoming more apparent afterwards. The horns of males increased in length over this study. In contrast, horn growth of females appeared to plateau coincident with the time that they would be expected to reach sexual maturity and begin to reproduce.
Acknowledgements
The authors would like to thank G. Jordaan for helping with statistical analysis in this article. Second, the authors would like to thank R.A.I. Morton from Tembani Wildlife (Pty) Ltd and Wildlife Stud Services for the measurement data used in this article.
Authors' contributions
CAS drafted and wrote the manuscript, questioning whether there were significant differences in horn growth traits between male and female animals. PCL guided the statistical analysis and suggested the methods. JHCVZ, LCH and GFJ assisted in supervising and editing the manuscript.
Conflict of interest declaration
The authors declare there is no conflict of interest regarding the publication of the manuscript.
References
Basson, S.R., 1989. 'n Voorlopige studie van die swartwitpens (Hippotragus niger Harris, 1838) in die Letabadistrik, Transvaal en moontlike faktore wat bevolkings groei beinvloed. Gravellotte. [ Links ]
Cloete, P.C., Van der Merwe, P. & Saayman, M., 2015. Game ranch profitability in South Africa. Second edition. ABSA, Cape Town. [ Links ]
Daza, A., Rey, A.I., Lopez-Carrasco, C. & Lopez-Bote, C.J., 2014. Effect of gender on growth performance, carcass characteristics and meat and fat quality of calves of Avileña-Negra Ibérica breed fattened under free-range conditions. Spanish J. Agric. Res. 12, 683-693. https://doi.org/10.5424/sjar/2014123-4693 [ Links ]
Du Toit, J.G., 1992. The sable antelope as a game ranch animal. In: B.L. Penzhorn (ed). Proceedings of a Symposium on 'The sable antelope as a game ranch Animal'. South African Veterinary Association, Onderstepoort. Pages 19-25 i [ Links ]
Grobler, J.H., 1979. The use of horn sections to determine the age of sable. Koedoe 22, 219-223. [ Links ]
Grobler, J.H., 1980a. Breeding biology and aspects of social behaviour of sable Hippotragus niger niger (Harris, 1838) in the Rhodes Matopos National Park, Zimbabwe. South African J. Wildl. Res. 10, 150-152 https://journals.co.za/doi/pdf/10.10520/AJA03794369_3018 [ Links ]
Grobler, J.H., 1980b. Body growth and age determination of the Sable Hippotragus Niger Niger (Harris, 1838). Koedoe 23, 131 -156. https://doi.org/10.4102/koedoe.v23i1.641 [ Links ]
Josling, G.C., Lepori, A.A., Neser, F.W.C., Lubout, P.C. & Van Wyk, J.B., 2019. Evaluating horn traits of economic importance in sable antelope (Hippotragus niger niger). S. Afr. J. Anim. Sci. 49, 40-49. [ Links ]
Juta 2011. Game Theft Act 105 of 1991. Juta & Co. Ltd. Cape Town, South Africa. [ Links ]
Knight, J.E. 2008. Developing trophy antlers. In: Manage your land for wildlife: A wildlife management handbook for Rocky Mountain landowners. Montana State University Extension, Bozeman, Montana. [ Links ]
Kriek, J.C., 2017. Manattu Private Game Reserve. https://mattanu.com/ [ Links ]
Lincoln, G.A., 1992. Biology of antlers. J. Zoology (London) 226, 517-528. https://zslpublications.onlinelibrary.wiley.com/doi/pdf/10.1111/j.1469-7998.1992.tb07495.x [ Links ]
Matthee, C.A., & Robinson, T.J., 1999. Mitochondrial DNA population structure of roan and sable antelope: implications for the translocation and conservation of the species. Mol. Ecol. 8, 227-238. https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1365-294X.1999.00556.x [ Links ]
Newbolt, C.H., Acker, P.K., Neuman, T.J., Hoffman, S.I., Ditchkoff, S.S. & Steury, T.D., 2017. Factors influencing reproductive success in male white-tailed deer. J. Wildlife Manage. 81, 206-217. 10.1002/jwmg.21191 [ Links ]
Rabie, J.H., 2011. Die herlewing en genetiese herstel van die swartwitpens. Deel 1. Wild Jag, 53-57. [ Links ]
Raphaka, K., & Dzama, K., 2009. Sex of calf and age of dam adjustment factors for birth and weaning weight in Tswana and composite beef cattle breeds in Botswana. S. Afr. J. Anim. Sci. 39, 296-300. [ Links ]
Ruiz, M., French, S.S., Demas, G.E. & Martins, E.P., 2010. Food supplementation and testosterone interact to influence reproductive behavior and immune function in Sceloporous graciosus. Horm. Behav. 57, 134-139 https://doi.org/10.1016/j.yhbeh.2009.09.019 [ Links ]
Saayman, M., van der Merwe, P. & Saayman, A., 2018. The economic impact of trophy hunting in the south African wildlife industry. Global Ecology and Conservation 16, e00510. https://doi.org/10.1016/j.gecco.2018.e00510. [ Links ]
Safari Club International, 2021. Method 1 entry form. https://safariclub.org/wp-content/uploads/2021/02/1-SCI-Simple-Horn-Method-Form-2021.pdf [ Links ]
Sempéré, A.J. & Boissin, J., 1981. Relationship between antler development and plasma androgen concentrations in adult roe deer (Capreolus capreolus). J. Reprod. Fertil. 62, 49-53. https://doi.org/10.1530/JRF.0.0620049. [ Links ]
Tucker, C.B., MacNeil, M.D. & Webster, A.B. (eds), 2020. Guide for the care and use of agricultural animals in research and teaching. 4th edition. American Dairy Science Association, American Society of Animal Science, and Poultry Science Association, Champaign, Illinois, USA. [ Links ]
Van Rooyen, N., Van Rooyen, J. & Van Rooyen, N., 2016. Handling and measuring trophies. In: J. du P. Bothma. & J.G. du Toit (eds). Game ranch management.. Sixth edition. Van Schaik, Pretoria. Pp. 719-753. [ Links ]
Warren, P., 2021. Personal communication. Piet Warren Plase, P.O. Box 1, Gravelotte, 0895. [ Links ]
West, P. M. & Packer, C., 2002. Sexual selection, temperature, and the lion's mane. science 297(5585), 1339-1343 DOI 10.1126/science.1073257 [ Links ]
Willisch, C.S., Biebach, I., Marreros, N., Ryser-Degiorgis, M.P. & Neuhaus, P., 2015. Horn growth and reproduction in a long-lived male mammal: No compensation for poor early-life horn growth. Evolutionary Biology 42, 1-11. 10.1007/s11692-014-9294-3 [ Links ]
Submitted 26 November 2021
Accepted 14 February 2022
Published 14 June 2022
# Corresponding author: craig.shepstone@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}