










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 no.1 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i1.13
Gene expression at different cell stages of in wfro-fertilized bovine embryos
N.M. Al-Malahi#; M.H. Daghestani; M.Q. Al-Ghadi; A.R. Alhimaidi
King Saud University, College of Science, Zoology Dept. Riyadh, Saudi Arabia
ABSTRACT
The birth rate of embryos produced in vitro (IVF) is still lower than that of embryos produced in vivo. Three major steps for the success of the IVF technique are maturation of immature oocytes, fertilization of matured oocytes, and culture of the resulting embryos. Studying mRNA expression in early embryonic development stages is important and can help to assess embryo quality and optimize production protocols in vitro. The current study aimed to determine the expression levels of developmentally important genes in different stages of bovine embryos produced in vitro. Cumulus-oocyte complexes (COCs) were collected from bovine ovaries and cultured in synthetic oviduct fluid (SOF) medium for 7 - 9 days. Embryos were collected at the time-points listed above, and mRNA expression of genes involved in pluripotency (OCT4), DNA methylation (DNMT1), apoptosis (BAX), and metabolism (GLUT1) and a heat shock protein (HSP70) was estimated from the 2-cell stage to the blastocyst stage of embryos. The results showed statistically significant differences in the relative abundance (RA) of OCT4, DNMT1, BAX, and GLUT1 gene transcripts among the different stages, whereas there were non-significant differences in the RA of HSP70 between these stages. In conclusion, gene expression levels differ among the developmental stages of embryos produced in vitro, possibly because of the timing of embryonic genome activation (EGA).
Keywords: bovine in vitro maturation, embryo production, gene expression, in vitro fertilization
Introduction
In vitro embryo production is an important biotechnology in cattle husbandry and breeding, and the use of this technique has increased greatly. Bovine embryos are produced around the world by commercial companies (Camargo et al., 2006; Abd El-Aziz et al., 2016; Stoecklein et al., 2021; Blaschka et al., 2021).
Certain steps are performed in in vitro production of embryos to mimic the in vivo conditions in different types of animals. These steps begin with the maturation of oocytes in vitro, which takes place within 20 - 24 hours in bovines (Wrenzycki, 2018; Damayanti et al., 2020), and the extrusion of the first polar body, which are prerequisites to fertilization and the initiation of embryonic development (Mehlmann, 2005; Sirard, 2016; Turhan et al., 2021). Therefore, the maturation of the oocyte is important for fertilization and preimplantation development (Barakat et al., 2018).
In vitro fertilization is the second step in in vitro production, which is characterized by the extrusion of the second polar body and the formation of the male and female pronuclei (Parrish, 2014). The in vitro culture of bovine embryos is the last step of in vitro production, which requires approximately 7 - 9 days of culture from the zygote stage. The events that occur in the embryo during this step include the first cleavage division, embryonic genome activation, morula compaction, and blastocyst formation (Wrenzycki, 2018; Ramos-Deus et al., 2020; Nogueira et al., 2021). Maternal transcripts stored within the oocyte during oogenesis regulate early embryonic development. Maternally derived transcripts are degraded as development progresses, whereas embryonic genome activation begins from the time of maternal to zygotic transition (MZT) (Graf et al., 2014).
Embryo quality can be assessed using genes that serve as genetic markers and play roles in the pre-and post-implantation development of embryos, where the expression of these genes correlates with the timing of embryonic genomic activation (Sadeesh et al., 2014a). These genes are involved in biological processes such as DNA methylation, which is accomplished by adding a methyl group to the fifth carbon atom of cytosine with the help of a group of enzymes known as DNA methyltransferases. This mechanism is critical in maintaining genome stability in preimplantation embryos (Sagirkaya et al., 2006; Urbanek-Olejnik et al., 2014; Uysal et al., 2015; Chen & Zhang, 2020). Pluripotent cell populations are maintained by the gene OCT4, which belongs to the POU (Pit-Oct-Unc) transcription factor family. This transcription factor is required for maintaining inner cell mass pluripotency, which is present in all cells at the morula stage, and is downregulated in the trophectoderm of the bovine blastocyst (Kurosaka et al., 2004; Hess et al., 2019). Glucose transport across the cell plasma membrane as a primary source of energy is mediated by the transporter GLUTI gene. After entering embryonic cells, glucose is metabolized via glycolysis, which generates ATP. This metabolism increases with glucose uptake, which is correlated with the timing of compaction and blastulation (Lopes et al., 2007; Ostrowska et al., 2015).
During preimplantation, genomic stability is necessary in the embryo, which is achieved by the maintenance of normal methylation patterns (Urbanek-Olejnik et al., 2014; Uysal et al., 2015; Chen & Zhang, 2020), homeostasis via apoptosis initiation (Korsmeyer, 1999; Li et al., 2009), and stress protection (Luft & Dix, 1999; Chen et al., 2018), all of which are critical in embryo development. This study aimed to determine the gene expression levels of developmentally important genes in different stages of bovine embryos produced in vitro during the preimplantation period.
Materials and methods
In vitro-fertilized bovine embryos were studied in different cleavage stages during the 7 - 9 days of culture. Three replicates of each embryonic stage (2-cell stage (18 embryos for each replicate), 4-cell stage (14 embryos), 8-16-cell stage (12 embryos), morula (8 embryos), and blastocyst (6 blastocysts) were collected and washed at least twice in 0.1% PVA-PBS, then frozen in 5 μ! of the same solution and held at -80 °C until RNA was extracted. (De Oliveira et al., 2005). The following genes were selected for the measurement of gene expression during the in vitro development of bovine embryos: OCT4 (pluripotency gene), DNMT1 (DNA methyltransferase), BAX (apoptosis gene), GLUTI (glucose transporter), and HSP70 (heat shock protein).
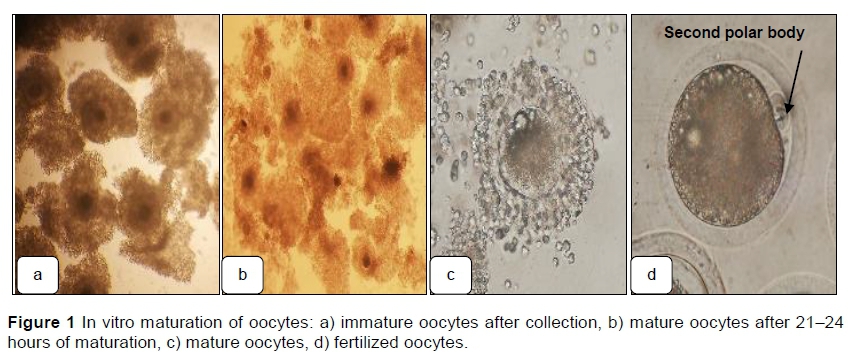
Cumulus oocyte complexes (COCs) were obtained by aspirating follicles from bovine ovaries collected after slaughter (Im et al., 1995; Galli et al., 2003; Saadeldin & Jang, 2018; Lira et al., 2020). After the COCs were washed in TCM-199 Earle's salt medium (Sigma M4530; Merck KGaA, Darmstadt, Germany) supplemented with 0.5 mM sodium pyruvate, 25 μg/ml gentamycin sulfate, 1 μg/ml estradiol-17p (Sigma E2758), 0.02 AU/mL FSH (Sigma F8174), 0.023 lU/mL LH (Sigma L5269), and 10% FBS, groups of 10 - 15 COCs were placed in 50 μl drops of maturation medium under mineral oil and cultured for 21 - 24 hours at 39 °C in an atmosphere of 5% CO2 in air with maximum humidity (Plourde et al., 2012; Cánepa et al., 2014a; Sprícigo et al., 2016; Ferré et al., 2020) (Figure 1). TCM-199 was used as the maturation medium. Photomicrographs of the oocytes and embryos were produced with a cell phone camera and an inverted microscope at 200x and 400x magnifications.
Frozen bull semen was thawed in 37 °C water and transferred to a discontinuous Percoll gradient (2 mL of 45% Percoll over 2 mL of 90% Percoll) (Machado et al., 2009; Saadeldin & Jang, 2018; Vega et al., 2018; Aguiar et al., 2019; Sá et al., 2020; Maziero et al., 2020; Nogueira et al., 2021). Then, the samples were centrifugea at 700 g for 30 min at room temperature (Daniels et al., 2000). The supernatant was discarded, and the spermatozoon pellet was resuspended in in vitro fertilization media (IVF-B.O) and kept in an incubator for three hours in a humidified atmosphere of 5% CO2 at 39 °C to increase capacitation. After 3 hours, sperm were counted using a hemocytometer and prepared at a final concentration of 2x106 cells/ml (Plourde et al., 2012; Cánepa et al., 2014a; Sprícigo et al., 2016; Sá et al., 2020).
Matured COCs were washed and transferred to 60 mm Petri dishes in 10 drops of 50 μ! each per dish, which were then covered with embryo-tested mineral oil. Ten to 15 oocytes were cultured in each droplet of in vitro fertilization medium (IVF-BO) supplemented with 1.25 mM pyruvate, 25 μg/ml gentamycin, 11.12 μg/mL heparin (Sigma H3149), and 3 mg/ml bovine serum albumin (Sigma A6003). Finally, motile sperm were prepared before being added to the COCs at a final concentration of 2x106 cells/mL, with 24 hours co-incubation in a humidified atmosphere of 5% CO2 at 39 °C (Cánepa et al., 2014a).
Fertilized and unfertilized COCs were mechanically denuded by repeated pipetting with a glass Pasteur pipette in a hyaluronidase enzyme solution to completely remove cumulus cells and were washed in the in vitro culture medium (IVC-SOF) (Caisson IVL05). Then, 20 - 25 zygotes were cultured in a droplet of the IVC-SOF supplemented with 0.34 mM sodium pyruvate, 1 mM L-glutamine, 50X MEM-essential amino acids (Sigma B6766), 100X MEM nonessential amino acids (Sigma M7145), 3 mg/mL BSA (Sigma A6003), 25 μg/ml gentamycin, 1.5 mM glucose, and 1 μg/ml EDTA in a 35 mm Petri dish, and the drops were covered with embryo-tested mineral oil. Embryos were cultured in an incubator for 7 - 9 days at 39 °C in 5% CO2, 5% O2, and 90% N2 atmosphere with high humidity inside the incubation chamber (Rodriguez-Alvarez et al., 2010; Cánepa et al., 2014a).
The RNA was isolated from the embryos with the PureLinkTM RNA mini kit (Cat. no. 12183-018A, Thermo Fisher Scientific, Waltham, Massachusetts, USA). Then 0.4 ml lysis buffer was added to each sample, followed by vortexing, and RNA was purified according to the manufacturer's instructions. Complementary DNA (cDNA) was synthesized with a high-capacity cDNA reverse transcription kit (Cat. no. 4368814, Thermo Fisher Scientific), with 10 μ! RNA, 2 μ! of 10X RT buffer, 2 μ! random primers, 0.8 μ! dNTP mix (100 mM), 1 μ! multiscribe reverse transcriptase and 4.2 μ! nuclease-free H2O were mixed in a 200 μ! polymerase chain reaction (PCR) tube. Then, the samples were placed in a thermocycler according to the following program: 25 °C for 10 min, 37 °C for 120 min, and 85 °C for 5 min. The synthesized cDNA was stored at -20 °C prior to real-time PCR.
Amplification with SYBR green master mix (Thermo Fisher Scientific) was performed on an Applied Biosystems ViiA 7 real-time PCR system (Thermo Fisher Scientific) in a 12.5 μ! reaction to assess the gene expression of OCT4, DNMT1, BAX, GLUTI, and HSP70 relative to that of the housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Forward and reverse primers used in these assays are shown in Table 1. All genes of interest were analysed in duplicate in clear 96-well plates containing multiple samples. Amplification was carried out in a 12.5 μ! reaction mixture containing 6.25 μ! of SYBR Green, 0.25 μ! of each forward and reverse primer, 2 μ! of cDNA temp!ate, and 3.75 μ! of nuc!ease-free water. The RT-PCR program was as follows: 50 °C for 2 min; 95 °C for 10 min; 40 cycles of denaturation at 95 °C for 15 seconds, annealing at 60 °C for 1 min and extension at 95 °C for 15 seconds; and a final extension at 60 °C for 1 min.
The comparative CT method (Schmittgen & Livak, 2008) was used for the relative quantification of target gene expression levels, which were normalized to the reference GAPDH gene. The ACT value was obtained by subtracting the GAPDH CT value for each sample from the target gene CT value. The AACT value was calculated by using the highest sample method ACT as an arbitrary constant, which was subtracted from all other ACT sample values. The changes in the gene expression of the target genes were determined by using the 2-AACT formula (Amarnath et al., 2007; EM et al., 2014).
Statistical analysis was performed with SPSS 20 software. For the analysis of relative differential gene expression (IVP) in bovine embryos, differences among means were analysed by one-way ANOVA, followed by multiple pairwise comparisons using Duncan's test. Data were presented as the mean ± standard error). P-values of less than 0.05 were considered significant (EM et al., 2014).
Results and Discussion
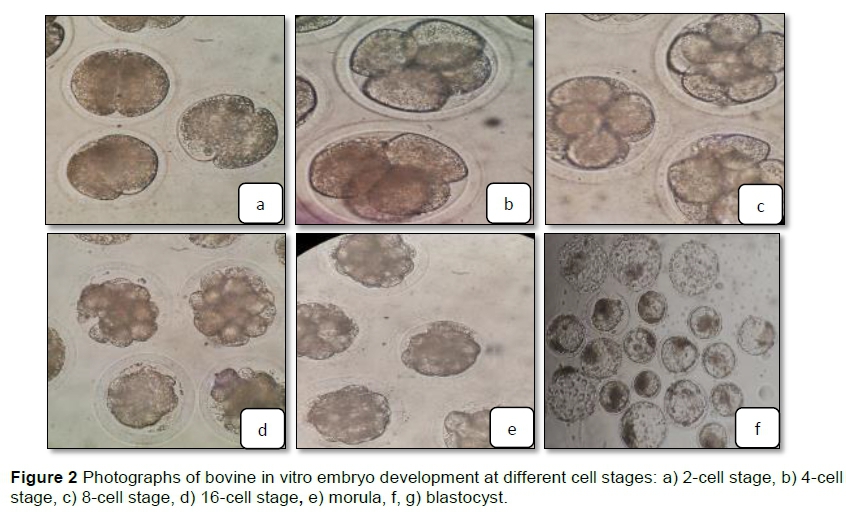
Representative photomicrographs of the IVF bovine embryos at different stages from the 2-cell stage to the blastula stage are shown in Figure 2.
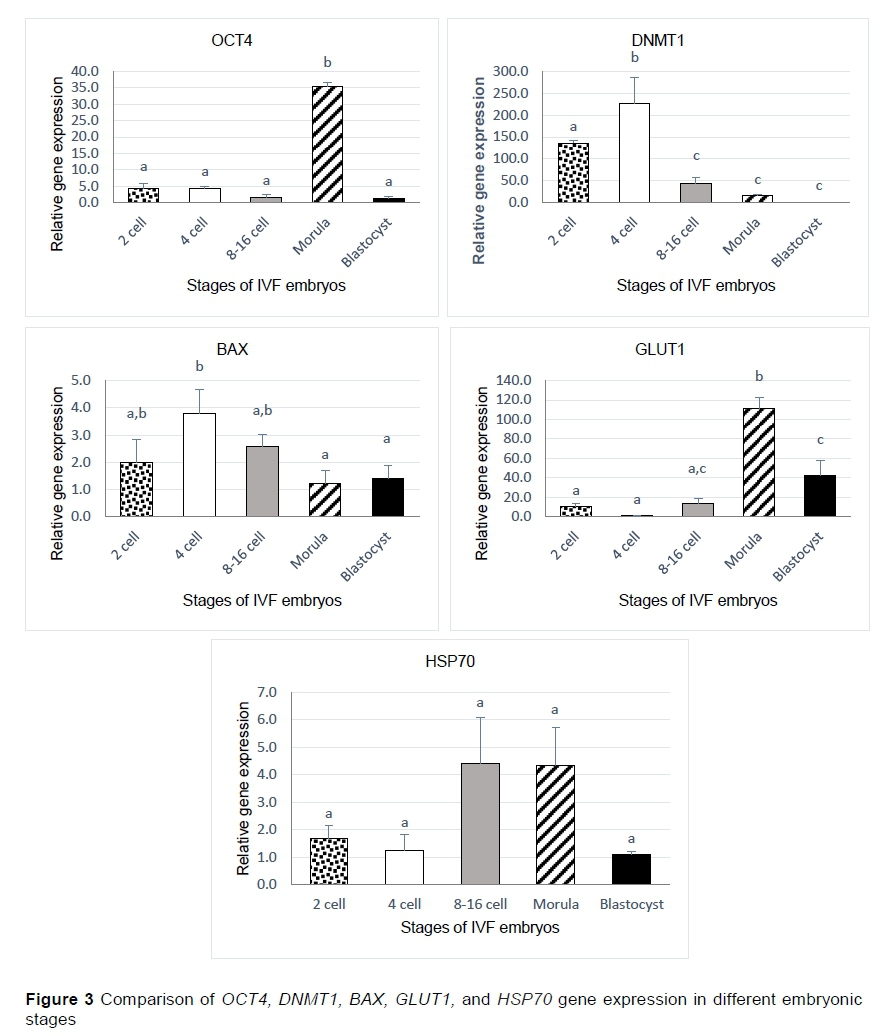
The OCT4, DNMT1, BAX, GLUTI, and HSP70 transcripts were detected in several cleavage stages of bovine embryos collected on different days of culture, including the 2-cell, 4-cell, 8 - 16 cell, morula, and blastocyst stages (Figure 2). The expression of the OCT4 gene was significantly higher in the morula stage than in the other stages according to the findings. Furthermore, relative to the earlier stages, the expression of the DNMT1 gene increased significantly in the 4-cell stage. BAX gene expression was significantly higher in the 4-cell stage than in the morula and blastocyst stages, but its level was nonsignificant in the 2- and 8-16-cell stages. The GLUT1 gene showed much higher expression in the morula stage than in the other stages. There were no significant differences in the expression of the HSP70 gene at any stage (Figure 3).
Embryonic genome activation occurs in waves with the timing varying by species; it occurs at the 2-cell stage in mouse embryos, the 4 - 8-cell stage in pig embryos, the 8 - 16-cell stage in bovine embryos (Sirard, 2012), and the 4 - 8-cell stage in humans (Braude et al., 1988).
The genes studied in this work play a crucial role in early embryonic development, and the current findings revealed changes in gene expression levels at different stages of embryonic development before and after embryonic genome activation. Some of these genes were down-regulated or up-regulated at distinct cell stages, which could be because of oxidative stress under the culture conditions, high maternal genome storage, or the expression of other genes that could alter the expression of these genes (Graf et al., 2014; Sadeesh et al., 2014a).
The OCT4 gene is important for the maintenance of a pluripotent cell population in preimplantation embryos (Kurosaka et al., 2004; Hess et al., 2019), and the expression of this gene was significantly higher in the morula stage than in the other examined stages. This result agrees with those of Kurosaka et al. (2004), who showed that the OCT4 transcript level starts to increase after embryonic genome activation and presents a sharp increase after compaction. Therefore, this gene is present in all cells of the morula stage and is downregulated in the trophectoderm (TE) of bovine blastocysts. These differences may explain the decrease in the expression of this gene in the blastocyst. The DNMT1 protein is responsible for DNA methylation, which is essential to normal embryonic development and cellular differentiation by silencing differentiation-associated genes and activating the critical genes for embryo development (Sagirkaya et al., 2006; Uysal et al., 2018). The expression of this gene in the 2- and 4-cell stages was significantly higher than in the other stages (8 - 16-cell, morula, and blastocyst), which showed gradual decreases in expression relative to the 2- and 4-cell stages. This result was consistent with those of Duan et al. (2019), who reported that although large amounts of DNMT1 mRNA were stored in oocytes, the DNMT1 mRNA level remained very low after embryonic genome activation in bovine embryos (Graf et al., 2014). This result was similar to the findings of Hou et al. (2007), who showed that the methylation level decreased after the 8-cell stage and that this decrease continued through the morula stage. Furthermore, expression was higher in the 4-cell stage than in the 2-cell stage, indicating that DNMT1 expression may have increased to suppress the expression of genes whose expression was not needed at this stage and to maintain the stability of gene expression states (Dor & Cedar, 2018).
Apoptosis induced by the expression of the BAX gene occurs in response to environmental stressors as a normal feature of pre-implantation development (Matwee et al., 2000; Fahrudin et al., 2002). The current study revealed that although the expression of the BAX gene was greatest at the 4-cell stage it was not significantly higher than that in the 2-cell, 8-cell, and 16-cell stages, but was significantly higher than that at the morula and blastocyst stages. These results are not in accord with those of EM et al. (2014), who reported that apoptosis was first observed in bovine in vitro-fertilized embryos at the 8- to 16-cell stages. However, these results do agree with the findings of other researchers (Byrne et al., 1999; Fahrudin et al., 2002) that suboptimal conditions in the culture system can induce apoptosis in bovine embryos produced in vitro and with the observation of cell death resistance gene expression reported by Hardy (1997). These findings may be explained by the observation of Cánepa et al. (2014b) that there is an interaction between HSP70 and BAX gene expression, possibly as a result of stress conditions, causing apoptosis to occur in embryos and thus increasing the expression of HSP70 while BAX expression is down-regulated to protect the embryos.
The cyto-protective factor HSP70 helps embryos recover from stress-induced damage (Cánepa et al., 2014b). This study revealed non-significant increased expression of HSP70 in the 8 to 16-cell and morula stages relative to the 2- and 4-cell and blastocyst stages. These results are consistent with those of Luft and Dix (1999), who reported that HSP70 was expressed beginning in the embryonic period for gene activation in cleavage-stage embryos. Thus, embryos at different developmental stages are exposed to a wide range of environmental stressors leading to the expression of HSP70.
The GLUT1 gene plays an important role in the diffusion of glucose across the cell plasma membrane, and glucose is an important energy substrate for the development of embryos (Lopes et al., 2007; Ostrowska et al., 2015; Arhin et al., 2018). A significant sharp increase in GLUTI gene expression was observed in the morula stage relative to the other stages. This result was corroborated by previous findings (Lequarre et al., 1997), which showed that glucose metabolism in bovine embryos is low during the first cleavages and increases sharply after the resumption genomic activity (8 - 16 cells). Other authors (Lopes et al., 2007) reported that during compaction and blastocyst formation glucose uptake by the embryo increases and causes increased expression of GLUTI. The expression of GLUT1 also increased in the blastocyst stage but was not significantly higher than in the morula stage of development. There was a non-significant difference relative to the 8-16-cell stage, which may be in accord with the return to higher levels of Glut-1 in trophectoderm cells compared with the inner cell mass cells reported in (Wrenzycki et al., 2003) and agrees with the findings of Wrenzycki et al. (2003) and Lopes et al. (2007).
Conclusions
The results of this work indicated that the levels of gene expression differ between different cell stages in embryos produced in vitro due either to the timing of embryonic genome activation or to in vitro conditions that alter the expression level of genes. This gene expression is important in early development to assess the normality of bovine embryos produced in vitro. The longer-term application is development of biomarkers for success of in vitro production of embryos (i.e., embryo quality). These biomarkers could be applied to research studies testing different in vitro embryo production strategies on reproductive success.
Acknowledgments
The authors sincerely acknowledge the Researcher Support Project (RSP 2021/232) for funding this work at King Saud University, Riyadh, Saudi Arabia. In addition, the authors thank the Researcher Support Service Unit at King Saud University for their help with techniques and the review of this manuscript. Our appreciation and thanks also go to the Central Riyadh slaughterhouse for their help in collecting bovine ovary samples.
Authors' Contributions
All of the authors contributed to the conceptualization of this study, ANA-M and MQA-G collected the samples and conducted the laboratory analyses, ANA-M performed the statistical analysis, interpreted the results and drafted the manuscript; MHD and aRa reviewed and edited the article, and ARA administered the project.
Conflicts of Interest Declaration
The authors of this manuscript have declared that there is no conflict of interest pertaining to this work.
References
Abd El-Aziz, A.H., Mahrous, U.E., Kamel, S.Z. & Sabek, A.A., 2016. Factors influencing in vitro production of bovine embryos: A review. Asian J. Anim. Vet. Adv. 11, 737-756. https://doi.org/10.3923/ajava.2016.737.756 [ Links ]
Aguiar, G.B., Caldas-Bussiere, M.C., Maciel, V.L., de Carvalho, C.S.P. & de Souza, C.L.M., 2019. Association of L- arginine with heparin on the sperm capacitation improves in vitro embryo production in bovine. Anim. Reprod. 16, 938-944. https://doi.org/10.21451/1984-3143-AR2019-0022 [ Links ]
Amarnath, D., Li, X., Kato, Y. & Tsunoda, Y., 2007. Gene expression in individual bovine somatic cell cloned embryos at the 8-cell and blastocyst stages of preimplantation development. J. Reprod. Dev. 53, 1247-63. https://doi.org/10.1262/jrd.19096 [ Links ]
Arhin, S.K., Lu, J., Xi, H. & Jin, X., 2018. Energy requirements in mammalian oogenesis. Cell. Mol. Biol. 64, 12-19. https://doi.org/10.14715/cmb/2018.64.10.3 [ Links ]
Barakat, I.A.H., Alajmi, R.A., Zoheir, K.M.A., Salem, L.M. & Al-Hemidiy, A.R., 2018. Gene expression and maturation evaluation of sheep oocytes cultured in medium supplemented with natural antioxidant source. South African J. Anim. Sci. 48, 261-270. https://doi.org/10.4314/sajas.v48i2.7 [ Links ]
Blaschka, C., Diers, S., Aravina, M., Geisler, S., Schuler, G. & Tetens, J., 2021. Evaluation of a small volume oil-free in vitro production system for bovine embryos. Vet. Med. Sci. 00, vms3.42. https://doi.org/10.1002/vms3.428 [ Links ]
Braude, P., Bolton, V. & Moore, S., 1988. Human gene expression first occurs between the four- and eight-cell stages of preimplantation development. Nature 332, 459-461. https://pubmed.ncbi.nlm.nih.gov/3352746/ [ Links ]
Byrne, A.T., Southgate, J., Brison, D.R. & Leese, H.J., 1999. Analysis of apoptosis in the preimplantation bovine embryo using TUNEL. J. Reprod. Fertil. 117, 97-105. https://doi.org/10.1530/jrf.0.1170097 [ Links ]
Camargo, L., Viana, J., Sá, W., Ferreira, A., Ramos, A. & Vale Filho, V., 2006. Factors influencing in vitro embryo production. Anim. Reprod 3, 19-28. http://www.cbra.org.br/pages/publicacoes/animalreproduction/issues/download/AR072%20Camargo%20pag19-28.pdf [ Links ]
Cánepa, M.J., Ortega, N.M., Monteleone, M.C., Mucci, N., Kaiser, G.G., Brocco, M. & Mutto, A., 2014a. Expression profile of genes as indicators of developmental competence and quality of in vitro fertilization and somatic cell nuclear transfer bovine embryos. PLoS One 9, 1-8. https://doi.org/10.1371/journal.pone.0108139 [ Links ]
Chen, B., Feder, M.E. & Kang, L., 2018. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 27, 3040-3054. https://doi.org/10.1111/mec.14769. [ Links ]
Chen, Z. & Zhang, Y., 2020. Role of mammalian DNA methyltransferases in development. Annu. Rev. Biochem. 89, 135-158 https://doi.org/10.1146/annurev-biochem-103019-102815. [ Links ]
Damayanti, E., Sonjaya, H., Hasbi, H. & Baco, S., 2020. Strategies to increase growth early embryo stages of bovine in achieving blastocysts in vitro: A review. IOP Conf. Ser. Earth Environ. Sci. 492, 012082. https://doi.org/10.1088/1755-1315/492/1/012082 [ Links ]
Daniels, R., Hall, V. & Trounson, A. O., 2000. Analysis of gene transcription in bovine nuclear transfer embryos reconstructed with granulosa cell nuclei. Biol. Reprod. 63, 1034-1040. https://doi.org/10.1095/biolreprod63.4.1034 [ Links ]
De Oliveira, A.T.D., Lopes, R.F.F. & Rodrigues, J.L., 2005. Gene expression and developmental competence of bovine embryos produced in vitro under varying embryo density conditions. Theriogenology 64, 1559-1572. https://doi.org/10.1016/j.theriogenology.2005.03.019 [ Links ]
Dor, Y. & Cedar, H., 2018. Principles of DNA methylation and their implications for biology and medicine. Lancet 392, 777-786. https://doi.org/10.1016/S0140-6736(18)31268-6 [ Links ]
Duan, J., Zhu, L., Dong, H., Zheng, X., Jiang, Z., Chen, J. & Tian, X.C., 2019. Analysis of mRNA abundance for histone variants, histone- and DNA-modifiers in bovine in vivo and in vitro oocytes and embryos. Sci. Rep. 9, 1-13. https://doi.org/10.1038/s41598-018-38083-4 [ Links ]
EM, S., Kataria, M., Balhara, S. & Yadav, P.S., 2014a. Expression profile of developmentally important genes between hand-made cloned buffalo embryos produced from reprogramming of donor cell with oocytes extract and selection of recipient cytoplast through brilliant cresyl blue staining and in vitro fertilized embryos. J. Assist. Reprod. Genet. 31, 1541-1552. https://doi.org/10.1007/s10815-014-0316-y [ Links ]
Fahrudin, M., Otoi, T., Karja, N.W.K., Mori, M., Murakami, M. & Suzuki, T., 2002. Analysis of DNA fragmentation in bovine somatic nuclear transfer embryos using TUNEL. Reproduction 124, 813-819. https://doi.org/10.1530/rep.0.1240813 [ Links ]
Ferré, L.B., Kjelland, M.E., Str0bech, L.B., Hyttel, P., Mermillod, P. & Ross, P.J., 2020. Review: Recent advances in bovine in vitro embryo production: Reproductive biotechnology history and methods. Animal 14, 991-1004 https://doi.Org/10.1017/S1751731119002775 [ Links ]
Galli, C., Duchi, R., Crotti, G., Turini, P., Ponderato, N., Colleoni, S., Lagutina, I. & Lazzari, G., 2003. Bovine embryo technologies. Theriogenology 59, 599-616. https://doi.org/10.1016/S0093-691X(02)01243-8 [ Links ]
Graf, A., Krebs, S., Zakhartchenko, V., Schwalb, B., Blum, H. & Wolf, E., 2014. Fine mapping of genome activation in bovine embryos by RNA sequencing. Proc. Natl. Acad. Sci. USA., 111, 4139-4144. https://doi.org/10.1073/pnas.1321569111 [ Links ]
Hardy, K., 1997. Cell death in the mammalian blastocyst. Mol. Hum. Reprod. 3, 919-925. https://doi.org/10.1093/molehr/3.10.919 [ Links ]
Hess, D. L., Kelly-Goss, M. R., Cherepanova, O. A., Nguyen, A. T., Baylis, R. A., Tkachenko, S., Annex, B. H., Peirce, S. M. & Owens, G. K., 2019. Perivascular cell-specific knockout of the stem cell pluripotency gene Oct4 inhibits angiogenesis. Nat. Commun. 10, 1-15. https://doi.org/10.1038/s41467-019-08811-z [ Links ]
Hou, J., Liu, L., Lei, T. H., Cui, X. H., An, X. R. & Chen, Y. F., 2007. Genomic DNA methylation patterns in bovine preimplantation embryos derived from in vitro fertilization. Sci. China, Ser. C Life Sci. 50, 56-61. https://doi.org/10.1007/s11427-007-0003-7. [ Links ]
Im, K. S., Kim, H. J., Chung, K. M., Kim, H. S. & Park, K. W., 1995. Effects of ovary type, oocyte grade, hormone, sperm concentration and fertilization medium on in vitro maturation, fertilization and development of bovine follicular oocytes. Asian-Australasian J. Anim. Sci. 8, 123-127. https://doi.org/10.5713/ajas.1995.123 [ Links ]
Korsmeyer, S. J., 1999. BCL-2 gene family and the regulation of programmed cell death. Cancer Res. 59, 1693-1701. https://doi.org/10.1016/0165-4608(96)85244-7 [ Links ]
Kurosaka, S., Eckardt, S. & McLaughlin, K.J., 2004. Pluripotent lineage definition in bovine embryos by Oct4 transcript localization. Biol. Reprod. 71, 1578-1582. https://doi.org/10.1095/biolreprod.104.029322 [ Links ]
Lequarre, A.S., Grisart, B., Moreau, B., Schuurbiers, N., Massif, A. & Dessy, F., 1997. Glucose metabolism during bovine preimplantation development: Analysis of gene expression in single oocytes and embryos. Mol. Reprod. Dev. 48, 216-226. https://onlinelibrary.wiley.com/doi/10.1002/(SICI)1098-2795(199710)48:2%3C216::AID-MRD9%3E3.0.CO;2-V [ Links ]
Li, H. J., Liu, D.J., Cang, M., Wang, L.M., Jin, M.Z., Ma, Y.Z. & Shorgan, B., 2009. Early apoptosis is associated with improved developmental potential in bovine oocytes. Anim. Reprod. Sci. 114, 89-98. https://pubmed.ncbi.nlm.nih.gov/19008057/ [ Links ]
Lira, A. da S., Chaves, R. de M., Moraes Junior, F. de J., Costa Junior, S.H., Amaral, B.K.L. do & Trovão, H.M.P. 2020. Use of melatonin in the in vitro production of bovine embryos. Rev. Bras. Saúde e Produção Anim. 21, 1-9. https://doi.org/10.1590/s1519-9940210322020 [ Links ]
Lopes, A.S., Wrenzycki, C., Ramsing, N.B., Herrmann, D., Niemann, H., L0vendahl, P., Greve, T. & Callesen, H., 2007. Respiration rates correlate with mRNA expression of G6PD and GLUT1 genes in individual bovine in vitro- produced blastocysts. Theriogenology 68, 223-236. https://doi.org/10.1016/j.theriogenology.2007.04.055 [ Links ]
Luft, J.C. & Dix, D.J., 1999. Hsp70 expression and function during embryogenesis. Cell Stress Chaperones 4, 162-70. [ Links ]
Machado, G.M., Carvalho, J.O., Filho, E.S., Caixeta, E.S., Franco, M.M., Rumpf, R. & Dode, M.A.N., 2009. Effect of Percoll volume, duration and force of centrifugation, on in vitro production and sex ratio of bovine embryos. Theriogenology 71, 1289-1297. https://doi.org/10.1016/j.theriogenology.2009.01.002 [ Links ]
Matwee, C., Betts, D.H. & King, W.A., 2000. Apoptosis in the early bovine embryo. Zygote 8, 57-68. https://doi.org/10.1017/S0967199400000836 [ Links ]
Maziero, R.R.D., Guaitolini, C.R.D.F., Paschoal, D.M., Crespilho, A.M., Monteiro, B.A., Lima, J.S. De, Sestari, D.A.O. & Landim-Alvarenga, F.D.C., 2020. Treatment with roscovitine and butyrolactone I prior to in vitro maturation alters blastocyst production. Zygote 28, 24-31. https://doi.org/10.1017/S0967199419000571 [ Links ]
Mehlmann, L.M., 2005. Stops and starts in mammalian oocytes: Recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 130, 791-799. https://doi.org/10.1530/rep.L00793 [ Links ]
Nogueira, B.G.R., Souza, L.F.A. de, Puelker, R.Z., Giometti, I.C., Firetti, S.M.G., Dias, T.S. dos S.B. & Castilho, C., 2021. Factors affecting the in vitro production of bovine embryos in a commercial program. Res. Soc. Dev. 10, e16110212264. https://rsdjournal.org/index.php/rsd/article/view/12264 [ Links ]
Ostrowska, M., Jarczak, J. & Zwierzchowski, L., 2015. Glucose transporters in cattle - A review. Anim. Sci. Pap. Reports 33, 191-212. [ Links ]
Parrish, J.J., 2014. Bovine in vitro fertilization: In vitro oocyte maturation and sperm capacitation with heparin. Theriogenology 81, 67-73. https://doi.org/10.1016/j.theriogenology.2013.08.005 [ Links ]
Plourde, D., Vigneault, C., Laflamme, I., Blondin, P. & Robert, C., 2012. Cellular and molecular characterization of the impact of laboratory setup on bovine in vitro embryo production. Theriogenology 77, 1767-1778.e1. https://doi.org/10.1016/j.theriogenology.2011.12.021 [ Links ]
Ramos-Deus, P., Santos Nascimento, P., Vieira, J.I.T., Chaves, M.S., Albuquerque, K.A., Ferreira-Silva, J.C., Grázia, J.G.V., Santos Filho, A.S., Batista, A.M., Teixeira, V.W.& Oliveira, M.A.L., 2020. Application of platelet-rich plasma in the in vitro production of bovine embryos. Trop. Anim. Health Prod. 52, 2931-2936. https://doi.org/10.1007/s11250-020-02307-5 [ Links ]
Rodriguez-Alvarez, L., Sharbati, J., Sharbati, S., Cox, J.F., Einspanier, R. & Castro, F.O., 2010. Differential gene expression in bovine elongated (Day 17) embryos produced by somatic cell nucleus transfer and in vitro fertilization. Theriogenology 74, 45-59. https://doi.org/10.1016/j.theriogenology.2009.12.018 . [ Links ]
Sá, N.A.R., Vieira, L.A., Ferreira, A.C.A., Cadenas, J., Bruno, J.B., Maside, C., Sousa, F.G.C., Cibin, F.W.S., Alves, B.G., Rodrigues, A.P.R., Leal-Cardoso, J.H., Gastal, E.L. & Figueiredo, J.R., 2020. Anethole supplementation during oocyte maturation improves in vitro production of bovine embryos. Reprod. Sci. 27, 1602-1608. https://doi.org/10.1007/s43032-020-00190-x [ Links ]
Saadeldin, I.M. & Jang, G., 2018. Sex differences in single IVF-derived bovine embryo cultured in chemically defined medium. Int. J. Vet. Sci. Med. 6, S78-S80. https://doi.org/10.1016/j.ijvsm.2018.01.003 [ Links ]
Sagirkaya, H., Misirlioglu, M., Kaya, A., First, N.L., Parrish, J.J. & Memili, E., 2006. Developmental and molecular correlates of bovine preimplantation embryos. Reproduction 131, 895-904. https://doi.org/10.1530/rep.L01021 [ Links ]
Schmittgen, T. & Livak, K., 2008. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3, 1101-1108. https://doi.org/10.1038/nprot.2008.73 [ Links ]
Sirard, M.A., 2012. Factors affecting oocyte and embryo transcriptomes. Reprod. Domest. Anim. 47, 148-155. https://doi.org/10.1111/j.1439-0531.2012.02069.x [ Links ]
Sirard, M.A., 2016. Somatic environment and germinal differentiation in antral follicle: The effect of FSH withdrawal and basal LH on oocyte competence acquisition in cattle. Theriogenology 86, 54-61. https://doi.org/10.1016/j.theriogenology.2016.04.018 [ Links ]
Sprícigo, J.F.W., Sena Netto, S.B., Muterlle, C.V., Rodrigues, S. de A.D., Leme, L.O., Guimarães, A.L., Caixeta, F.M.C., Franco, M.M., Pivato, I. & Dode, M.A.N., 2016. Intrafollicular transfer of fresh and vitrified immature bovine oocytes. Theriogenology 86, 2054-2062. https://doi.org/10.1016/j.theriogenology.2016.07.003 [ Links ]
Stoecklein, K.S., Ortega, M.S., Spate, L.D., Murphy, C.N. & Prather, R.S., 2021. Improved cryopreservation of in vitro produced bovine embryos using FGF2, LIF, and IGF1. PLoS One 16, e0243727. https://doi.org/10.1371/journal.pone.0243727 [ Links ]
Turhan, A., Pereira, M.T., Schuler, G., Bleul, U. & Kowalewski, M.P., 2021. Hypoxia-inducible factor (HIF1alpha) inhibition modulates cumulus cell function and affects bovine oocyte maturation in vitrof. Biol. Reprod. 104, 479-491. https://doi.org/10.1093/biolre/ioaa196 [ Links ]
Urbanek-Olejnik, K., Liszewska, M. & Kostka, G., 2014. The effect of phenobarbital on gene expression levels of p53 and Dnmt1 in the liver of Wistar rats. Rocz . Paήstwowego Za kladu Hig. 65, 199-203. https://www.cabdirect.org/globalhealth/abstract/20143371717 [ Links ]
Uysal, F., Akkoyunlu, G. & Ozturk, S., 2015. Dynamic expression of DNA methyltransferases (DNMTs) in oocytes and early embryos. Biochimie 116, 103-113. https://doi.org/10.1016/j.biochi.2015.06.019 [ Links ]
Uysal, F., Ozturk, S. & Akkoyunlu, G., 2018. Superovulation alters DNA methyltransferase protein expression in mouse oocytes and early embryos. J. Assist. Reprod. Genet. 35, 503-513. https://doi.org/10.1007/s10815-017-1087-z [ Links ]
Vega, W.H.O., Quirino, C.R., Bartholazzi-Junior, A., Rua, M.A.S., Serapião, R.V. & Oliveira, C.S., 2018. Variants in the CYP19A1 gene can affect in vitro embryo production traits in cattle. J. Assist. Reprod. Genet. 35, 2233-2241 https://doi.org/10.1007/s10815-018-1320-4 [ Links ]
Wrenzycki, C., 2018. Gene expression analysis and in vitro production procedures for bovine preimplantation embryos: Past highlights, present concepts and future prospects. Reprod. Domest. Anim. 53, 14-19. https://doi.org/10.1111/rda.13260 [ Links ]
Wrenzycki, C., Herrmann, D. & Niemann, H., 2003. Timing of blastocys t expansion affects spatial messenger RNA expression patterns of genes in bovine blastocysts produced in vitro. Biol. Reprod. 68, 2073-2080. https://doi.org/10.1095/biolreprod.102.012104 [ Links ]
Submitted 27 August 2021
Accepted 4 January 2022
Published 22 February 2022
# Correspondence: nalmalahi@ksu.edu.sa, nalmalahi-4@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}