










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 n.1 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i1.11
Effects of flushing and various doses of β-Carotene and vitamin E applications on the reproductive performance of ewes
S. Ozis Altincekic#; M. Koyuncu; S. Duru
Department of Animal Science, Faculty of Agriculture, Bursa Uludag University, 16059, Bursa, Turkey
ABSTRACT
The aim of this study was to determine whether fertility could be enhanced in Kivircik ewes during the anoestrous period by flushing or by administering an injectable (Ovostim) containing β-Carotene and vitamin E. Three-year-old Kivircik ewes (n = 99) were used. The ewes were randomly divided into four groups: flushing, a single dose of Ovostim (5 ml/ewe) before the breeding season, a dose of Ovostim 10 days before the breeding season and a second dose at its start, and an untreated control. Results showed that pregnancy rate was higher in the treated groups than in the control (P <0.001). The single dose of Ovostim produced more multiple births (72%) and the highest litter size (1.64) compared with the other treatments. Although the double-dose of Ovostim influenced the ewes in the anoestrous period to the point of becoming pregnant, it failed to favourably affect the number of lambs born per ewe lambing. In conclusion, flushing the ewes or administering a single dose of Ovostim was shown to be advantageous in comparison with untreated ewes or those given two doses of Ovostim.
Keywords: β-Carotene, fertility, flushing, sheep, vitamin E
Introduction
Adequate nutrition plays a vital role in the reproductive function of animals. Supplemental feeding can affect fertility in ruminants directly and indirectly. This effect can be attributed to regulation of the secretion of the luteinizing hormone (LH) by the gonadotropin releasing hormone (GnRH), which is secreted from the hypothalamus (Schillo, 1992; Robinson et al., 2006). For sheep, flushing is common practice (Cirne et al., 2016) before and during the breeding season by providing additional energy in the form of supplemental feed. Flushing encourages ovulation in ewes and increases rates of ovulation, survival of embryo, and fecundity (Morley et al., 1978; Scaramuzzi et al., 2006; Sabra & Hassan, 2008). For anoestrous ewes, flushing might be sufficient to induce ovulation and increase fertility without administering exogenous hormones (Öziş Altinçekiç et al., 2018).
Vitamin A is an essential nutrient and its deficiency can affect reproduction negatively in males and females (McDowell, 2000). Vitamin A does not exist as such in plants, but its antecedents, such as β-Carotene (carotenoids), are available in vegetable materials (Chew, 1993). As the antecedent of vitamin A and retinoid, β-Carotene encourages a series of physiological events that are important at cellular level with multiple biological actions (Harrison et al., 2012; Eroglu & Harrison, 2013). The β-Carotene is turned into vitamin A in the intestinal mucosa, liver, and other body tissues (Borel et al., 2005). One international unit (IU) of vitamin A is equal to 0.03 μg of vitamin A or 0.6 μg of β-Carotene (Altınışık,, 2021). Because β-Carotene is fat soluble, its absorption is generally similar to that of fat (Deming & Erdman, 1999).
The provision of β-Carotene via an injection during sexual activity increased the observed oestrus and oestradiol-ΙΤβ concentrations (Salem et al., 2015). In addition, positive correlations were reported between the β-Carotene level in plasma and the amounts of follicle liquid, luteal tissue, and corpus luteum weight (Arikan & Rodway, 2001; Ayaşan & Karakozak, 2009). β-Carotene is used as the source of vitamin A in the ovaries, affecting the tearing of the follicle membrane during ovulation and giving the corpus luteum its characteristic bright yellow colour (Haliloglu et al., 2002). In farm animals, β-Carotene decreases the risk of early embryonic death, which may result from stimulating progesterone release from luteal cells (Jackson et al., 1981; Graves-Hoagland et al., 1989). Deficiencies in β-Carotene were also reported to lead to delayed ovulation and formation of the corpus luteum, decreased progesterone synthesis, increased follicular and luteal cysts, and higher incidences of embryonic and foetal deaths in the first months of pregnancy (Hemken & Bremel, 1982; Arikan & Rodway, 2000). Schams et al. (1977) stated that the time interval between the emergence of the LH peak and ovulation is twice as long in animals with β-Carotene deficiency. Not all of the results in the literature pertaining to the effects of β-Carotene supplementation were in agreement. Findings observed a negative effect (Folman et al., 1987), a positive effect (Özpinar et al., 1995; Sonat et al., 2014) and no effect on reproduction (Wang et al., 1987; Kaewlamun et al., 2011; TrojaCanec et al., 2012) .
The purpose of this study was to investigate the effects of flushing, and providing β-Carotene via injection on the reproductive performance of ewes. Secondarily, early growth of the lambs from treated ewes was monitored.
Material and methods
Approval to carry out the study was given by the Ethics Council of Animal Experiments of Bursa Uludag University (No 2020-08-02). The study used 99 three-year-old multiparous Kivircik ewes weighing between 57 and 62 kg that were bred on the Research and Application Farm of the Faculty Agriculture of Bursa Uludag University. This farm is located in the humid tropical lowland 100 m above sea level at longitude 29 °E and latitude 40 °N The average minimum temperature at the farm is 10.4 °C, average maximum temperature is 20.5 °C, and annual rainfall is 522.3 mm. This study started in August, during which all the ewes were in the anoestrous period.
The ewes were randomly assigned to four groups. Group I (n = 25) was flushed for two weeks prior to mating and during the first week of mating. Group II (n = 25) was injected with beta-carotene and vitamin E (Ovostim 5 ml/head, Provet, Turkey) as a single dose two weeks before mating. Group III (n = 25) was given two injections of Ovostim, 5 ml/ewe two weeks before mating and 5 ml/ewe on the day the rams were introduced. Group IV (n = 24) was not subjected to either treatment, and served as the control. Ovostim is a commercial injectable solution, which contains 15 mg of beta-carotene and 20 mg of vitamin E per ml. The mating period lasted 35 days.
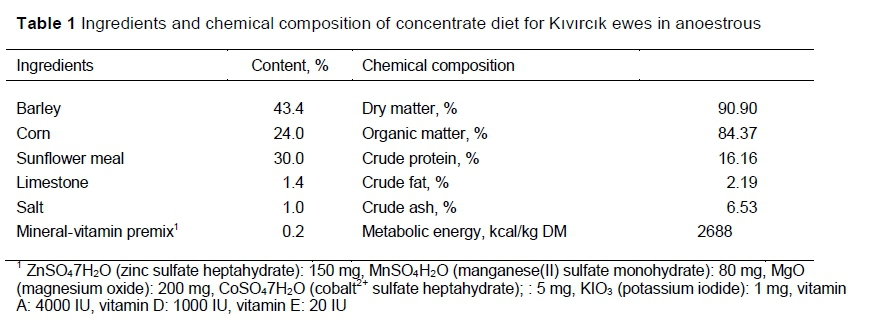
Before mating, all the ewes except for those in group I grazed the rangeland during the day and were kept in a shelter at night. The ewes were grazed on a pasture consisting of a mixture of common vetch (Vicia sativa L), Hungarian vetch (Vicia pannonica L), alfalfa (Medicago sativa) and sainfoin (Onobrychis sativa). The ewes in group I were given an unlimited supply of hay and alfalfa and fed twice a day with 750 g/ewe/day of a concentrate ration (Table 1) that was prepared on the farm. The metabolizable energy content of the supplement was calculated according to NRC (2007). The ewes were allowed to reach clean water and licking blocks in their partitions.
Lambing occurred in January and February. To meet their increased nutrient needs in late pregnancy, the ewes were given 300-400 g of concentrate feed per ewe in addition to ad libitum access to pasture according to the seasonal conditions. Within the first hour of being born, the lambs were made to take colostrum, weighed, and individually identified. Groups were formed in the data to indicate light (2.0 - 3.3 kg), medium (3.4 - 4.7 kg), and heavy (4.8 - 6.2 kg) birth weights. The lambs were kept with their mothers for about two weeks after birth. From the age of 2 - 3 weeks, the lambs were kept in the shelter during the day when their mothers went out to pasture and rejoined their mothers when they returned. The lambs were weaned when they were about two months old. Weaning age was divided into short, medium, and long groups of 56 - 67 days, 68 - 79 days, and 80 - 90 days, respectively.
The mating, birth and the lamb growth data were recorded regularly. These metrics were calculated from the data: pregnancy rate, %: number of pregnant ewes / number of ewes mated χ 100; lambing rate, %: number of ewes lambing / number of pregnant ewes χ 100; multiple rate, %: number of multiple lambing ewes / number of pregnant ewes χ 100; fecundity: number of lambs born / ewe mated; litter size: number of lambs born / ewe lambing; survival rate, %: number lambs weaned / number of lambs born χ 100; gestation productivity, kg: total weight of lambs live born from each 100 mated ewes; total productivity, kg: total weight of lambs at weaning time from each 100 mated ewes; and daily weight gain (DWG), g: weaning weight (WW) (kg), kg - birth weight (BW) (kg) / weaning age (WA) (days).
The categorical data were analysed using a chi-square (χ2) test to determine the significance of differences among treatments. To determine the factors affecting BW, WW and DWG an analysis of variance was conducted using the generalized linear model procedure of Minitab 19.0 (Minitab, LLC, State College, Pennsylvania, USA).Tukey's multiple comparison test was used to determine which means were significantly different. The statistical models used for the birth and weaning weights and daily weight increases were as follows:

where: μ = the mean, ai = the effect of the ith treatment, bj = the effect of the jth birth type (single vs multiple), ck = the effect of the kth sex (male vs female), dl =the effect of the lth birth weight group, fm = the effect of the mth weaning age group, and eijkl and eijklmn = the residual error.
Results and Discussion
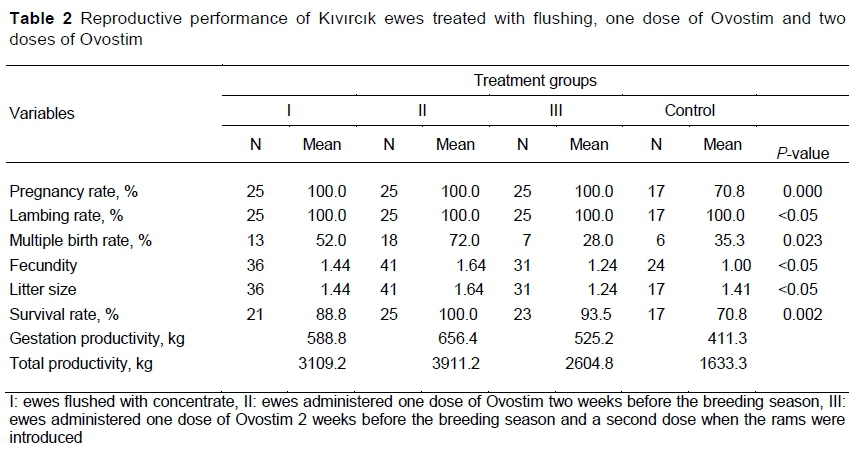
Differences in the pregnancy and multiple birth rates between the treatments were significant (P <0.01) (Table 2). Pregnancy rate of the CON group was about 30% lower than for the other three groups.
Flushing promotes oestrus successfully in ewes. Nutritional deficiencies are known to affect reproductive performance through irregularities in the reproductive cycle, delayed oestrus and ovulation, decreased pregnancy rates and increased embryonic losses (Ayasan & Karakozak, 2009). In ewes, a rise in energy intake before the breeding season increases reproductive performance by enhancing the concentrations of blood metabolites such as glucose to affect folliculogenesis and ovulation (Zabuli et al, 2010). Thus, two to three weeks before the breeding season and over its course, flushing should be used to enhance reproductive efficiency by increasing ovulation rate and consequently birth rate (Habibizad et al, 2015). In present study, Group 1 had a positive influence on pregnancy rate and the frequency of multiple births. Similarly, Abu and Ella (2006), Scaramuzzi et al (2006), and Naqvi et al (2011) reported that the pre-mating flushing diet promoted ovulation in many sheep breeds and increased ovulation rate, embryo viability, and fecundity. While the pregnancy rate obtained in Group 1 was similar to the value obtained by Shahneh et al (2008), it was higher than the 80% observed by Köse et al (2016) and 85% seen by Shaukat et al (2020). The multiple birth rate seen in Group 1 was much higher than the 12% found by Köse et al (2016) and the 32% seen by Mohajer et al (2012) in earlier studies of flushing. While the fecundity value of 1.44 obtained in Group I was lower than the 1.76 and 1.50 observed by Shaukat et al (2020), it was within the range of values seen by Acero et al (2008) as a result of the flushing applications at various energy levels. The results obtained from flushing programmes might have arisen from the body condition of the ewes (Gottardi et al, 2014), ration contents (Roche et al, 2011; Rekik et al, 2012), and the duration of flushing (Habibizad et al, 2015).
Ovulation, oestrus interval, and corpus luteum take shape in their standard courses in animals in the presence of β-Carotene, and reproductive issues can result from the deficiency of β-Carotene (Özpinar et al, 1998). However, Al-Tekreeti and Torun (2020) reported that when β-Carotene was applied alone, it did not affect reproductive performance, and that β-carotene with vitamin E and selenium was more efficient than β-Carotene alone. Groups II and III provided different amounts of Ovostim to the ewes and resulted in a numerical difference in the multiple birth rate, with the higher dose decreasing the frequency of multiple births. However, there were no difference between these treatments in the pregnancy or lambing rates, which were higher than observed in similar earlier studies (Kaçar et al, 2008; Al-Tekreeti & Torun, 2020). The present results were in line with the findings reported by Özpinar et al (1995) that the β-Carotene injection applied to the ewes at intervals of 20 days increased the pregnancy rate to the first insemination, prolificacy and twining rate. The numerically lower multiple birth rate obtained in Group III indicated that the dose of β-Carotene could be important and might lead to very different results among studies. In this study, ewes that received the single-dose of Ovostim were more fecund than those studied by Al-Tekreeti and Torun (2020). Al-Tekreeti and Torun (2020) found β-Carotene alone and in combination with vitamin E and selenium produced multiple birth rates of 55% and 77%, respectively, in Alpine goats, and 50% and 62.5% in Saanen goats. These results could indicate that the increased reproductive performance resulted from the vitamin E rather than the β-Carotene. Several environmental factors (feeding, duration of daylight exposure and treatment doses, etc.) might lead to differences between studies of pregnancy rate.
To interpret the treatment effects on survival rate, it should be noted that the Ovostim product contained vitamin E, which might decrease mortality by positively affecting growth, whereas the β-Carotene acted as an antioxidant in stimulating immune function (Chew, 1987; Avci et al, 2000; Stephensen, 2013). In the present study, the survival rate of lambs in Group II was the highest, but the survival rate of lambs in Group III also exceeded that of Group 1 and Group IV. The survival rates of the lambs in the present study were slightly higher than those reported for other sheep breeds of Turkey (Akçapinar et al, 2000; Karaoglu et al, 2001).
In this study, although the double dose of Ovostim positively contributed to the pregnancy, the rate of multiple births attributed to that treatment was significantly low. Whereas Folman et al (1987) reported that high carotene intake had a negative effect on the reproductive performance of dairy cows, other researchers stated that β-Carotene did not have any effect on reproductive performance (Wang et al, 1987; Kaewlamun et al, 2011; TrojaCanec et al, 2012). Arikan and Rodway (2000) reported that β-Carotene in high concentrations probably decreased the stimulation of progesterone production in the presence of LH owing to the β-Carotene's luteolytic effect. Brozos et al (2007) found that β-Carotene supplement did not affect the LH concentration in the ewes. Meza-Herrera et al (2013) determined that although the β-Carotene supplement decreased the LH concentration in goats, it increased the follicular growth and the ovulation rate. Thus, the possibility of a stimulating effect of β-Carotene on ovarian function that takes place via an LH-independent mechanism needs to be considered. Lopez-Flores et al (2020) explained that β-Carotene not only affected the ovulation rate and the ovarian modulator function measured as an antral follicular number, but it had a role as the modifier of LH release. Therefore, the dose of Ovostim administered two weeks before the mating period apparently stimulated the hormonal regulation in the desired direction. However, it is likely that the second dose applied during the mating season affected the number of ovulations by markedly altering the release of LH, and thus affected multiple birth rates negatively. Additional studies are needed to reveal the relationships between β-Carotene and vitamin A in the blood and reproductive hormone levels.
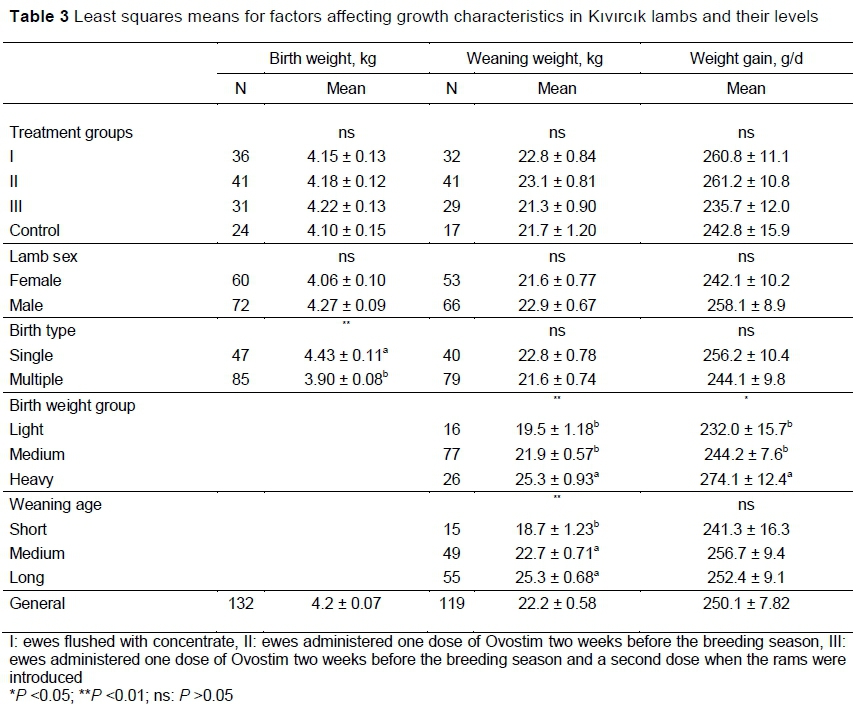
The treatments had no significant effects on BW, DWG, or WW (Table 3). However, Groups II and 1 increased the BW, WW, and DWG relative Group IV. Bodyweight in the present study was higher than that reported by Acero et al. (2008), most likely as a result of better nutrition. As is commonly known, single-born lambs were significantly heavier than twins (P <0.05), but male and female lambs did not differ in their growth. The effects of BW groups were significant for WW (P <0.01) and DWG (P <0.05) with lambs that were heavier at birth growing faster to weaning. Naturally, the lambs that were older at weaning were heavier.
Conclusion
Flushing or providing extra β-Carotene and vitamin E via the Ovostim injectable could produce a significant improvement in the reproductive performance of anoestrous Kivircik ewes. However, a double dose of the Ovostim injectable did not provide sufficient advantage to offset the associated extra cost and labour required. Future studies evaluating the effects of various β-Carotene levels on reproductive performance should consider the association of β-Carotene and circulating levels of vitamin A as they affect reproductive hormone levels. In addition, since the β-Carotene concentration in animal tissues can be affected by the season, similar studies in different seasons might provide more meaningful results.
Authors' Contributions
SOA and MK designed the study, SD conducted the statistical analyses, SOA collected the data and wrote manuscript, MK reviewed the manuscript.
Conflict of Interest Declaration
The authors declare no conflict of interest.
References
Abu El-Ella, A.A., 2006. Response of Barki ewes to treatment with gonadotrophin hormones and energy supplementation (flushing). Egypt. J. Sheep, Goat and Desert Anim. Sci. 1, 73-88. [ Links ]
Acero, A., Valencia, E., Rodriguez, A. & Randel, P.F., 2008. Effects of flushing with two energy levels on goat reproductive performance. Livest. Res. Rural Dev. 20, 1-8. https://www.researchgate.net/publication/275830691_Effects_of_ flushing_with_two_energy_levels_on_goat_reproductive_performance [ Links ]
Al-Tekreeti, H.Y.T. & Torun, O., 2020. Department of Animal Science. Annual Report. Ç.Ü Fen ve Mühendislik Bilim. Derg. 39, 121-129. [ Links ]
Altinisik, M., 2021. Vitaminler. Vitamin tanimi ve tarihçe. Vitaminler, saglikli beslenme. https://www.mustafaaltinisik.org.uk/89-1-10.pdf [ Links ]
Arikan, Ç. & Rodway, R.G., 2000. Effect of cyclodextrin-encapsulated β-Carotene on progesterone production by bovine luteal cells. Anim. Reprod. Sci. 64, 149-160. https://doi.org/10.1016/S0378-4320(00)00202-5 [ Links ]
Arikan, Ç. & Rodway, R.G., 2001. Seasonal variation in bovine luteal concentrations of β-Carotene. Turkish J. Vet. Anim. Sci. 25, 165-168 [ Links ]
Avci, M., Karakilçik, Z. & Kanat, R., 2000. Vitamin A, E ve Selenyumun Koyunlarda Döl Verimi ve Bazi Biyokimyasal Parametre Düzeyleri ile Kuzularinda Yasama Gücü ve Canli Agirlik Üzerine Etkisi. Turkish J. Vet. Anim. Sci. 24, 45-50 [ Links ]
Ayasan, T. & Karakozak, E., 2009. Hayvan beslemede β-Karoten kullanilmasi ve etkileri. Kafkas Univ. Vet. Fak. Derg. 16(4), 697-705. (InTurkish) https://doi.org/10.9775/kvfd.2010.2008 [ Links ]
Borel, P., Drai, J., Faure, H., Fayol, V., Galabert, C., Laromiguière, M. & Le Moël, G., 2005. Recent knowledge about intestinal absorption and cleavage of carotenoids]. Ann. Biol. Clin. (Paris). 63, 165-177 (Abstract. Article inFrench). [ Links ]
Brozos, C.N., Kiossis, E., Fthenakis, G.C., Tsousis, G. & Boscos, C., 2007. Supplementation of lactating ewes with a glucogenic preparation or β-Carotene in mid- to late lactation on subsequent milk yield and luteinizing hormone secretion. Can. J. Anim. Sci. 87, 185-190. https://doi.org/10.4141/A06-080 [ Links ]
Chew, B.P., 1987. Symposium: Immune Function: Relationship of Nutrition and Disease Control. Vitamin A and β-Carotene on host defense. J. Dairy Sci. 70, 2732-2743. https://doi.org/10.3168/jds.S0022-0302(87)80346-6 [ Links ]
Chew, B.P., 1993. Role of carotenoids in the immune response. J. Dairy Sci. 76, 2804-2811. https://doi.org/10.3168/jds.S0022-0302(93)77619-5 [ Links ]
Cirne, L.G.A., Da Silva Sobrinho, A.G., Oliveira, M.E.F., Barbosa, J.C., De Oliveira, G.J.C., Bagaldo, A.R., De Carvalho, G.G.P. & Moreno, G.M.B., 2016. Reproductive performance of Ile de France ewes under dietary supplementation before and during the breeding season. Semin. Agrar. 37, 269-278. DOI:10.5433/1679-0359.2016v37n1p269 [ Links ]
Deming, D.M. & Erdman, J.W., 1999. Mammalian carotenoid absorption and metabolism. Pure Appl. Chem. 71, 2213-2223. https://doi.org/10.1351/pac199971122213 [ Links ]
Eroglu, A. & Harrison, E.H., 2013. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 54, 1719-1730. https://doi.org/10.1194/jlr.R039537 [ Links ]
Folman, Y., Ascarelli, I., Kraus, D. & Barash, H., 1987. Adverse effect of β-Carotene in diet on fertility of dairy cows. J. Dairy Sci. 70, 357-366. https://doi.org/10.3168/jds.S0022-0302(87)80017-6 [ Links ]
Gottardi, F.P., Souza Júnior, A., Barbosa, Y.G.S., Marques, C.A.T., Bezerra, L.R., Araújo, M.J., Mingoti, G.Z. & Torreão, J.N.C., 2014. Efeito do flushing sobre o desempenho reprodutivo de ovelhas Morada Nova e Santa Inês submetidas à inseminação artificial em tempo fixo. Arq. Bras. Med. Vet. e Zootec. 66, 329-338. https://doi.org/10.1590/1678-41626103 (Abstract in English) [ Links ]
Graves-Hoagland, R.L., Hoagland, T.A. & Woody, C.O., 1989. Relationship of plasma β-Carotene and vitamin A to luteal function in postpartum cattle. J. Dairy Sci. 72, 1854-1858. https://doi.org/10.3168/jds.S0022-0302(89)79303-6 [ Links ]
Habibizad, J., Riasi, A., Kohram, H. & Rahmani, H. R., 2015. Effect of long-term or short-term supplementation of high energy or high energy-protein diets on ovarian follicles and blood metabolites and hormones in ewes. Small Rumin. Res. 132, 37-43. https://doi.org/10.1016/j.smallrumres.2015.10.004 [ Links ]
Haliloglu, S., Baspinar, N., Serpek, B., Erdem, H. & Bulut, Z., 2002. Vitamin A and β-Carotene levels in plasma, corpus luteum and follicular fluid of cyclic and pregnant cattle. Reprod. Domest. Anim. 37, 96-99. https://doi.org/10.1046/j.1439-0531.2002.00338.x [ Links ]
Harrison, E.H., Dela Sena, C., Eroglu, A. & Fleshman, M. K., 2012. The formation, occurrence, and function of β- apocarotenoids: β-Carotene metabolites that may modulate nuclear receptor signaling. Am. J. Clin. Nutr. 96,1189-1192. https://doi.org/10.3945/ajcn.112.034843 [ Links ]
Hemken, R.W. & Bremel, D.H., 1982. Possible role of beta-carotene in improving fertility in dairy cattle. J. Dairy Sci. 65, 1069-1073. https://doi.org/10.3168/jds.S0022-0302(82)82314-X [ Links ]
Jackson, P.S., Furr, B.J.A. & Johnson, C.T., 1981. Endocrine and ovarian changes in dairy cattle fed a low β-Carotene diet during an oestrus synchronisation regime. Res. Vet. Sci. 31, 377-383. https://doi.org/10.1016/s0034-5288(18)32475-5 [ Links ]
Kaçar, C., Kamiloglu, N. N., Gürbulak, K., Pancharci, S. M., Güngör, Ö., Güvenç, K. & Saban, E., 2008. The Effect of administration of testosterone antibody, β-Carotene and Vitamin E on multiple pregnancy and MDA (Malondialdehyde) in Tuj breed sheep in non-breeding season. Kafkas Univ. Vet. Fak. Derg. 14, 51-56. https://doi.org/10.9775/kvfd.2008.03-a (In Turkish, summary in English) [ Links ]
Kaewlamun, W., Okouyi, M., Humblot, P., Techakumphu, M. & Ponter, A. A., 2011. Does supplementing dairy cows with β-Carotene during the dry period affect postpartum ovarian activity, progesterone, and cervical and uterine involution? Theriogenology 75, 1029-1038. https://doi.org/10.1016/j.theriogenology.2010.11.010 [ Links ]
Karaoglu, M., Macit, M., Emsen, H., 2001. A study on the growth properties and livability of Tushin lambs. Turk. J. Vet. Anim. Sei. 25, 261-266. [ Links ]
Kaymakçi, M. 2006. Üreme Biyolojisi. Ege Üniversitesi Ziraat Fakültesi Yayin no: 503, izmir. [ Links ]
Köse, M., Kirbas, M., Bülbül, B., Dursun, Ç. & Demirei, U., 2016. Akkaraman irki koyunlarda flushing + koç etkisi ya da farkli dozlarda gebe kisrak serum gonadotropini uygulamalariyla kuzu üretiminin arttirilabilirliginin arastirilmasi. Ataturk Univ. Vet. Bilim. Derg. 11, 54-59. https://dergipark.org.tr/tr/pub/ataunivbd/issue/25027/264255 [ Links ]
Lopez-Flores, N.M., Meza-Herrera, C.A., Perez-Marin, C., Blaehe, D., Arellano-Rodríguez, G., Zuniga-Gareia, S., Navarrete-Molina, C., De La Pena, C.G., Rosales-Nieto, C.A. & Veliz-Deras, F.G., 2020. Precision beta-earotene supplementation enhaneed ovarian funetion and the LH release pattern in yearling erossbred anestrous goats. Animals 10, 1-10. https://doi.org/10.3390/ani10040659 [ Links ]
MeDowell, L.R., 2000. Vitamins in animal and human nutrition. Iowa State University Press, Ames. https://www.perlego.eom/book/2788445/vitamins-in-animal-and-human-nutrition-pdf?utm_souree=google&utm_medium=epe&eampaignid=15853719261&adgroupid=133053614878&gelid=CjwK CAiAgbiQBhAHEiwAuQ6BkhD4fMtgLAh2izJwEysb7jmvXUGJTD3W1s0iq7xt17tqYIe-hPK0kxoCKP4QAvD_BwE [ Links ]
Meza-Herrera, C.A., Vargas-Beltran, F., Vergara-Hernandez, H.P., Maeias-Cruz, U., Avendano-Reyes, L., Rodriguez-Martinez, R., Arellano-Rodriguez, G. & Veliz-Deras, F.G. 2013. Beta-earotene supplementation inereases ovulation rate without an inerement in LH seeretion in eyelie goats. Reprod. Biol. 13, 51-57. https://doi.org/10.1016/j.repbio.2013.01.171 [ Links ]
Mohajer, M., Alimon, A.R., Yaakub, H.B., Naslaji, A.N. & Toghdory, A., 2012. Effeets of energy level and PMSG dose on reproduetive performanee of Zel ewes bred to Shal or Zel rams. J. Anim. Vet. Adv. 11, 809-813. https://doi.org/10.3923/javaa.2012.809.813 [ Links ]
Morley, F.H.W., White, D.H., Kenney, P.A. & Davis, I.F., 1978. Predieting ovulation rate from liveweight in ewes. Agrie. Syst. 3, 27-45. https://doi.org/10.1016/0308-521X(78)90004-5 [ Links ]
Naqvi, S.M.K., Soren, N.M. & Karim, S.A., 2011. Effeet of eoneentrate supplementation on performanee, ovarian response, and some bioehemieal profile of Malpura ewes. Trop. Anim. Health Prod. 43, 905-913. https://doi.org/10.1007/s11250-011-9782-8 [ Links ]
NRC (National Researeh Couneil), 2007. Nutrient requirements of small ruminants: Sheep, goats, eervids, and New World eamelids. The National Aeademies Press, Washington, DC. https://www.nap.edu/eatalog/11654/nutrient-requirements-of-small-ruminants-sheep-goats-eervids-and-new [ Links ]
Özis Altinçekiç, Ç., Koyuncu, M. & Duru, S., 2018. Kivircik koyunlarinda flushinge ek olarak farkli dozlarda GKSH uygulamalarinin döl verimine etkisi. Mediterr. Agrie. Sei. 31, 87-91. https://doi.org/10.29136/mediterranean.348592 [ Links ]
Özpinar, H., Çenel, H.S., Özpinar, A. & Çekgül, E., 1998. ineklerde döl verimi ile serumdaki β-Carotene, A ve E vitamin düzeyleri arasindaki iliskiler. Doga Türk Vet Hay Derg 13, 273-282. [ Links ]
Özpinar, H., Özpinar, A., Bilal, T., Türkalp, I. & Firat, A., 1995. Pharmaeokineties of intramuseular administered beta- earotene and its effeets on reproduetion in sheep. Wien. Tierarztl. Monatssehr. 82, 229-231 [ Links ]
Rekik, M., Gonzalez-Bulnes, A., Lassoued, N., Ben Salem, H., Tounsi, A. & Ben Salem, I., 2012. The eaetus effeet: An alternative to the lupin effeet for inereasing ovulation rate in sheep reared in semi-arid regions? J. Anim. Physiol. Anim. Nutr. 96, 242-249. https://doi.org/10.1111/j.1439-0396.2011.01145.x [ Links ]
Robinson, J.J., Ashworth, C.J., Rooke, J.A., Mitehell, L.M. & MeEvoy, T.G., 2006. Nutrition and fertility in ruminant livestoek. Anim. Feed Sei. Teehnol. 126, 259-276. https://doi.org/10.1016/j.anifeedsei.2005.08.006 [ Links ]
Roehe, J.R., Burke, C.R., Meier, S. & Walker, C.G., 2011. Nutrition x reproduetion interaetion in pasture-based systems: Is nutrition a faetor in reproduetive failure? Anim. Prod. Sei. 51, 1045-1066. https://doi.org/10.1071/AN10162 [ Links ]
Sabra, H. & Hassan, S.S., 2008. Effeet of new regime of nutritional flushing on reproductive performances of Egyptian Barki ewes. Glob. Vet. 2, 28-31 [ Links ]
Salem, A., El-shahawy, N.A. & Soliman, I.A., 2015. Effeet of beta-carotene injection on estrus, vitamin A and estradiol- 17β concentrations in pubertal Farafra ewe lambs. Egypt. J. Anim. Prod. 52, 123-128. https://doi.org/10.21608/ejap.2015.93629 [ Links ]
Searamuzzi, R.J., Campbell, B.K., Downing, J.A., Kendall, N.R., Khalid, M., Munoz-Gutiérrez, M. & Somehit, A., 2006. A review of the effeets of supplementary nutrition in the ewe on the eoneentrations of reproduetive and metabolie hormones and the meehanisms that regulate follieulogenesis and ovulation rate. Reprod. Nutr. Dev. 46, 339-354. https://doi.org/10.1051/rnd:2006016 [ Links ]
Sehams, D., Hoffman, B., Lotthammer, K. H. & Ahlswede, L., 1977. A speeifie vitamin A independent effeet of beta earotene on eattle fertility. 4. Effeet on hormonal parameters during the eyele. Dtseh. Tierarztl. Woehensehr. 84, 307-310. https://pubmed.nebi.nlm.nih.gov/336337/ [ Links ]
Sehillo, K.K., 1992. Effeets of dietary energy on eontrol of luteinizing hormone seeretion in eattle and sheep. J. Anim. Sei. 70, 1271-1282. https://doi.org/10.2527/1992.7041271x [ Links ]
Shahneh, Z., Sadeghipanah, A., Barfourooshi, H.J. & Emami-mibody, M.A., 2008. Effeets of equine ehorionie gonadotropin (eCG) administration and flushing on reproduetive performanee in Nadooshan goats of Iran. Afriean J. Bioteehnol. 7, 3373-3379. http://www.academicjournals.org/AJB [ Links ]
Shaukat, A., ur Rehman, T., Shukat, R., Rajput, S.A., Shaukat, S., Naeem, M.A., Hassan, M., Fatima, T., Ahmad, F., Saleem, M.U., Arooj, F., Mehfooz, A. & Qureshi, A.S., 2020. Effeets of nutrient flushing on produetion and reproduetive performanee of teddy goats (Capra hircus). Pak. J. Zool. 52, 457-463. https://doi.org/10.17582/journal.pjz/20190330090301 [ Links ]
Sonat, A., Tekin, K. & Akçay, E., 2014. G6G / Ovysnch protokolü ile senkronize edilen ineklerde β -karoten enjeksiyonun döl verimi üzerine etkisi. Vet. Hek. Der. 85, 17-23 [ Links ]
Stephensen, CB., 2013. Provitamin A earotenoids and immune funetion. In: S. Tanumihardjo (ed). Carotenoids and Human Health. Humana Press, New York, NY. Pp 261-270 https://link.springer.eom/book/10.1007/978-1-62703-203-2 [ Links ]
Trojacanec, S., Bobos, S. & Pajic, M., 2012. Influence of β-earotene and vitamin A supplementation on the ovarian activity of dairy eows with ehronie fertility impairment. Vet. Arh. 82, 567-575. [ Links ]
Wang, J.Y., Hafi, C.B., Owen, F.G. & Larson, L. L., 1988. Effeet of beta-earotene supplementation on periparturient health and reproduetion of Holstein eows. Anim. Reprod. Sei. 15, 139-144. https://pubmed.nebi.nlm.nih.gov/3372802/ [ Links ]
Zabuli, J., Tanaka, T., Lu, W. & Kamomae, H., 2010. Intermittent nutritional stimulus by short-term treatment of high-energy diet promotes ovarian performanee together with inereases in blood levels of glueose and insulin in eyeling goats. Anim. Reprod. Sei. 122, 288-293. https://doi.org/10.1016/j.anireprosei.2010.09.005 [ Links ]
Submitted 3 July 2021
Accepted 11 November 2021
Published 22 February 2022
# Corresponding author: senizozis@gmail.com


{kind=link}
{kind=link}
{kind=link}