










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.52 n.1 Pretoria 2022
http://dx.doi.org/10.4314/sajas.v52i1.10
Quality of ram semen cryopreserved in egg yolk extender supplemented with myricetin
L.C.P. ArrudaI, #; R.A.J.A. SilvaI; F.C.C. MergulhãoI; M.M. MonteiroI; J.A.S. CostaI; A.M. BatistaI; P.L.J. Monteiro JúniorII; D.C.M. SealI; M.M.P. GuerraI
IAndrology Laboratory (ANDROLAB), Veterinary Medicine Department - UFRPE, Recife, Pernambuco, Brazil
IIDepartment of Animal and Dairy Science, University of Wisconsin-Madison, Madison, WI 53706 USA
ABSTRACT
The aim of this study was to evaluate the ability of the antioxidant myricetin to protect ovine spermatozoa during freezing-thawing. Five semen pools, obtained from four rams, were diluted and frozen in Tris-egg yolk extender (5% glycerol) supplemented with myricetin at 0, 20, 30, 40, 100, 200, 300, and 400 nM. After thawing at 37 °C for 30 seconds and incubation (37 °C) for two hours, these variables were measured: kinematics, plasma and acrosome membrane integrity, mitochondrial membrane potential, intracellular reactive oxygen species (ROS) levels, lipid peroxidation and membrane stability. There was no interaction between treatment and time, and no direct effect of treatments for the kinematics parameters (P >0.09; P >0.2113, respectively) and flow cytometry (P >0.41; P > 0.52, respectively). The means of progressive motility, curvilinear velocity, straight-line velocity, average path velocity, linearity, amplitude of lateral head displacement, percentage of intact plasma membrane and acrosome membrane, oxidative stress and membrane stability were reduced during incubation (P <0.04), whereas the cross-beat frequency increased (P =0.0001) throughout incubation. Thus, the addition of myricetin to the semen extender does not produce an antioxidant effect on ram semen in vitro.
Keywords: antioxidant, cryopreservation, flavonoids, sheep, sperm
Introduction
Flavonoids may interact with the lipid components of biological membranes. Lipid interactions, in most of the cases, are limited to the polar region of the lipid bilayer. However, the depth of penetration in the membrane depends on its structure. Owing to the distribution of these compounds in the lipid bilayer of cell membranes, these agents can protect the membrane from oxidation (Hendrich, 2006). Myricetin is a flavonoid that has antioxidant activity. It is commonly found in fruits, vegetables, nuts, berries, teas, and red wine (Huang et al., 2015). Its antioxidant activity has been reported in various cell types (Huang et al., 2010; Yan et al., 2021; Kimura et al., 2021). Its effect on spermatozoa remains to be clarified, because the antioxidant or pro-oxidant effects of flavonoids depend on their chemical structure, the concentration at which they are used, their capacity to interact with membranes, and the source of free radicals on which they will act (Barreiros et al., 2006; Chobot & Hadacek, 2011). Myricetin is not easily incorporated into the cell membrane. Nevertheless, for both liposomes and the human erythrocytes, it had the highest antioxidant potential compared with other flavonoids (Hendrich, 2006). The incorporation of myricetin in the membrane of spermatozoa can be efficient in protecting sperm against the oxidative damage caused by the semen freezing process owing to its antioxidant potential.
Oxidative damage of the sperm plasma membrane is caused by an imbalance between reactive oxygen species (ROS) and antioxidants, which can occur during semen processing (freezing-thawing) (Naseer et al., 2015; Amidi et al., 2016; Seifi-Jamadi et al., 2016). Oxidation is favoured because of the large amount of polyunsaturated fatty acids present in the sperm membrane, which are the main target of ROS attacks (Aitken et al., 2016; Hammerstedtl, 1993). Such oxidation results in a detrimental effect on the fertilizing capacity of male gametes (Aitken et al., 2016; Cross & Hanks, 1991). For fertilization to occur, sperm cells need to have intact plasma and acrosome membranes (Mogielnicka-Brzozowska et al., 2015).
The fertilization capacity of spermatozoa is affected by oxidative damage caused by ROS production in their membranes (Agarwal et al., 2014).
In cryopreservation, the use of an extender supplemented with substances capable of avoiding the damage caused by ROS can improve post-thaw semen quality (Maia et al., 2009). Therefore, this study aimed to evaluate the antioxidant effect of myricetin on ovine spermatozoa during freezing and thawing.
Materials and Methods
All experimental procedures were approved by the Committee on Ethics in the Use of Animals (CEUA) of the Universidade Federal Rural de Pernambuco - Brazil (licence 047/2015 CEUA-UFRPE). Four sexually mature rams were used (two Dorper and two White Dorper). They had a good history of fertility and were kept on property located in the municipality of Camaragibe, Pernambuco, Brazil (07° 96'93'' S 34°99'52'' W). The animals were submitted to intensive management and were fed commercial concentrate (500 g/day), elephant grass (Pennisetum purpureum), protein mineral salt and water ad libitum. Ejaculates were collected using an artificial vagina in the presence of a female as a dummy. Semen was collected on alternate days, totalling five collections per animal (20 ejaculates).
Immediately after collection, the ejaculates were evaluated using macroscopic and microscopic analyses, under a phase contrast microscope (x100, BX41-Olympus, Japan). Then, ejaculates with >70% motility were used to form a pool (n = 5). Each pool was evaluated for motility (subjective) with a phase-contrast microscope (x100, BX41-Olympus, Japan). The sperm concentration was determined in a Neubauer chamber at a dilution of 1:400 in saline formol.
All reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA), except for the fluorochromes CM-H2DCFDA (dichlorodihydrofluorescein 5- (e-6) -carboxy-2 ', 7'-diacetate), C11-BODIPY581/591 (4,4-difluoro-5- (4-phenyl-1,3-butadienyl) -4-bora-3a, 4a-diaza-s-indacene-3-undecanoic acid) and YO-PRO ®-1 iodide, which were purchased from Molecular Probes (Life Technologies, Eugene, USA), and phosphate-buffered saline, Dulbecco's Ca+2 and Mg+2 free (PBS), which were purchased from Gibco (Life Technologies, USA).
Fluorochrome stock solutions were prepared as follows: PI (25 mg/mL), JC-1 (5 mg/mL), CM-H2DCFDA (0.5 mM), C11-BODIPY581/591 (2 mM), merocyanine 540 (M540) (54 mM) and Yo-Pro-1 (1 mM) in DMSO (dimethylsulfoxide) and FITC-conjugated to peanut agglutinin (FITC-PNA) (1 mg/mL) in PBS. The work solutions were JC-1 (153 μM) and C11-BODIPY581/591 (0.4 mM) in DMSO and FITC-PNA (0.04 mg/mL), PI (0.5 mg/mL), CM-H2DCFDA (50 μM), M540 (270 μM) and Yo-Pro-1 (2.5 μM) in PBS. All solutions were maintained at -20 °C until use and stored for a maximum of 30 days. The semen cryopreservation extender was composed of 3.605 g of Tris, 2.024 g of citric acid, 1.188 g of fructose, 100 mL of distilled water, 20% egg yolk, 5% glycerol, 100 IU penicillin and 50 mg streptomycin, with a pH of 7.2. Before the addition of glycerol and antimicrobials, the extender was centrifuged twice (5500 x g/1 h). After centrifugation, the interface formed between the pellet and a thin layer of supernatant was harvested. After this procedure, the extender was filtered (0.45 microns and 0.22 microns), added to glycerol and antimicrobials, and homogenized. The stock solution of myricetin (31.42 mM) was prepared in dymethylsulfide (DMSO). The extender was fractionated and supplemented with myricetin in different concentrations and stored at -20 °C until use.
After evaluation, the semen pool was diluted in the extender supplemented with myricetin at 0, 20, 30, 40, 100, 200, 300, and 400 nM to a final concentration of 200 x 106 spermatozoa/mL and sent to a laboratory at room temperature (27 °C), where they were packaged in straws (0.25 mL) and frozen in an automated system (TK 3000®, TK Tecnologia em Congelação Ltd., Uberaba, Brazil). The interval between harvest and sample packaging in the freezing system was approximately two hours. The refrigeration curve showed a temperature reduction of -0.25 °C/min to 5 °C, then remained at this temperature for 120 min (stabilization time). Then, the freezing curve was started, with a reduction of -20 °C/min until it reached -120 °C, at which time the straws were immersed and stored in liquid nitrogen (-196 °C).
At the time of analysis, two straws from each experimental group were thawed in a water bath (37 °C) for 30 seconds, pooled and incubated at (37 °C) for 15 minutes. Then, the samples were diluted (1:4) in the extender without glycerol, incubated at 37 °C for 5 minutes and evaluated immediately. After two hours of incubation at 37 °C kinematic observations, plasma membrane integrity (iMPA), mitochondrial membrane potential (MMP), intracellular ROS levels (iROS), lipid peroxidation (LPO), and membrane stability (eMP) were recorded.
The kinematic variables were measured with a 2.5 μΙ_ aliquot of semen deposited on a slide and covered with a coverslip. Both the slide and coverslip were preheated to 37 °C. The evaluation took place using a phase-contrast microscope (X 100, Eclipse 50i, Nikon, Japan). Images of the sperm were captured with a Basler A312FC digital camera (Basler Vision Technologies™, Germany). At least five random fields were chosen, recording at least 500 spermatozoa. Total motility (TM) (%), progressive motility (PM) (%), linearity (LIN) (%) and straightness (STR) (%); curvilinear velocity (VCL) (μm/s), straight-line velocity (VSL) (μη/s) and average path velocity (VAP) (μm/s); amplitude of lateral head displacement (ALH) (μm) and beat-cross frequency (BCF) (Hz) were measured using SCA™ software version 5.1 (Microptics, SL, Barcelona, Spain).
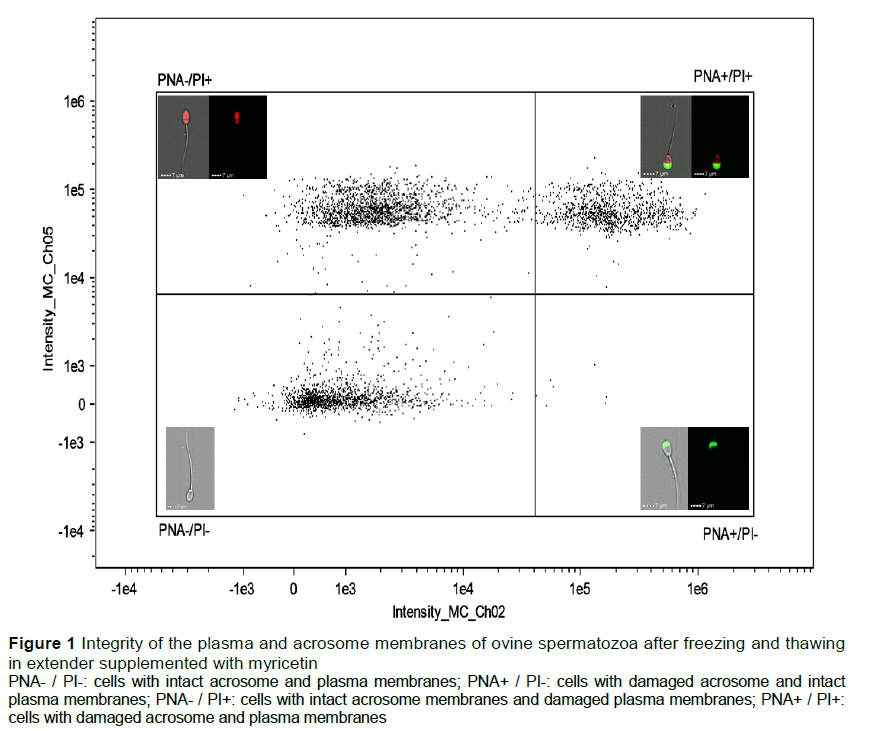
For flow cytometry, two aliquots (300 μL) of semen from each experimental group were placed in micro-tubes (2.0 mL), 1.7 mL of PBS was added slowly through the tube wall, and the aliquots were homogenized. Then, the sample was centrifuged at 100 x g for 5 minutes to remove the extender. The supernatant was discarded, and the pellets were pooled and re-suspended in 250 μL of PBS. Aliquots of 50 μL were transferred into five micro-tubes (1.5 mL) for each of the following analyses. For these evaluations, iMPA, 5 μL of the working solution of FITC-PNA and 5 μL of PI were added to the sample. The sample was incubated for 5 minutes at room temperature and then evaluated. Cells that showed PNA- / PI- labelling without staining were classified as having intact acrosome and plasma membranes. Cells with PNA+ / PI-green labelling were classified as having damaged acrosome membranes and intact plasma membranes. Cells with PNA- / PI+ red labelling were classified as having intact acrosome membranes and damaged plasma membranes. Finally, cells with PNA+ / PI+ green and green labelling were classified as having damaged acrosome and plasma membranes (Figure 1).
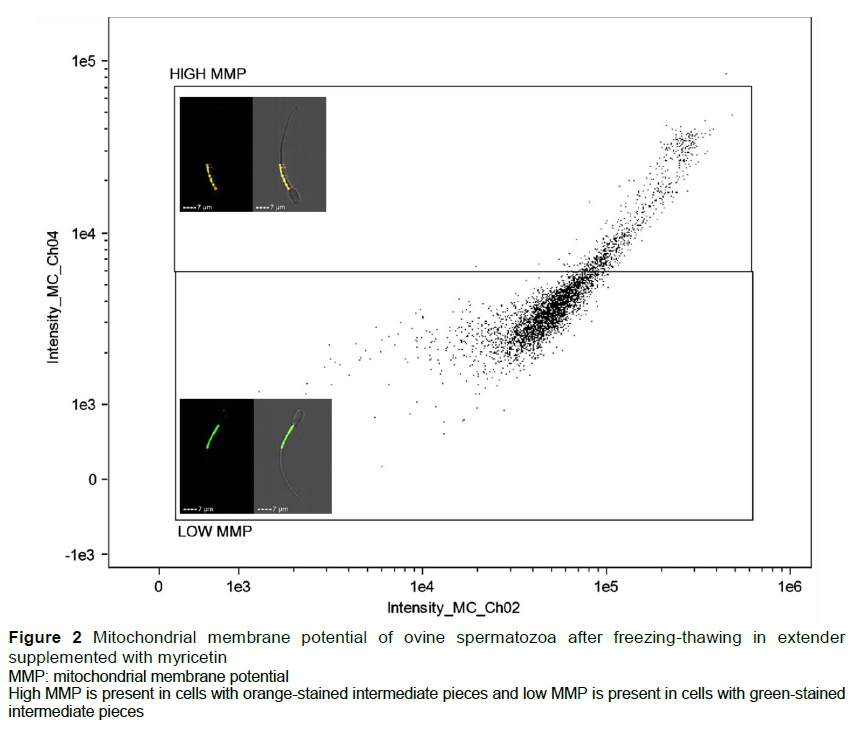
For MMP analysis, 5 μL of the working solution of JC-1 was added to the semen sample. The samples were then incubated for 5 min at room temperature before being read. Orange-stained cells in the midpiece region were classified as having high mitochondrial membrane potential (HMMP) and green-stained cells in the midpiece region were classified as having low mitochondrial membrane potential (LMMP) (Figure 2).
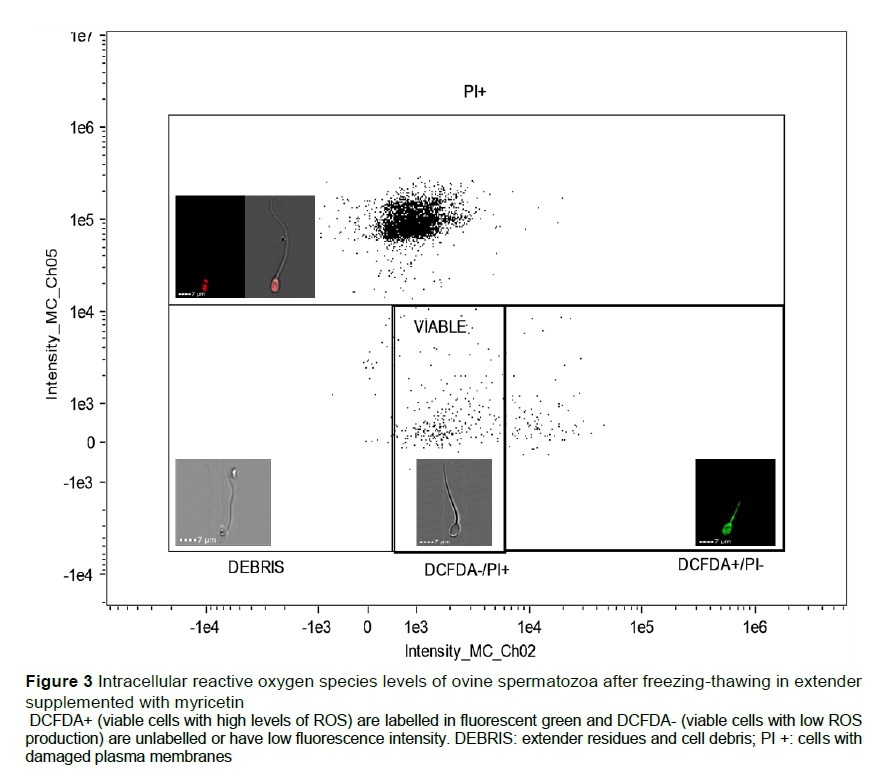
Levels of iROS were measured by labelling the samples with the fluorochrome CM-H2DCFDA. The amount of 5 μΙ_ of the working solution was added to the semen samples, which were incubated in a water bath (37 °C/30 min). Then, 5 μL of working PI solution was added and samples were incubated for 5 min at room temperature, after which time they were read. The results were expressed as a percentage of viable cells with high levels of ROS (DCFDA+) for those labelled with fluorescent green, or viable cells with low ROS (DCFDA-) for unlabelled cells or those with low fluorescence intensity (Figure 3).
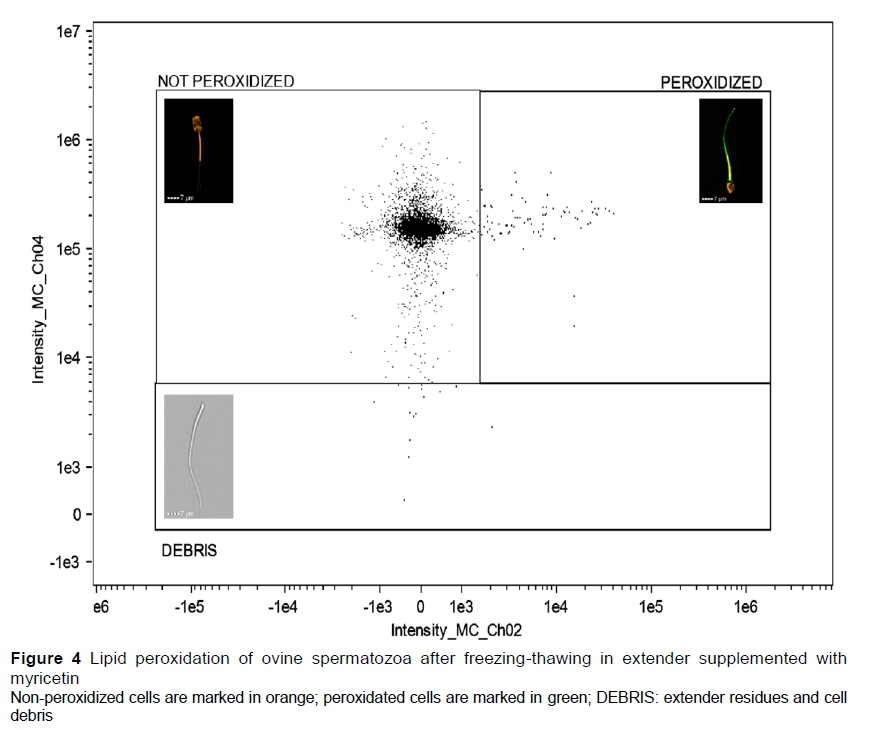
The fluorochrome C11-BODIPY581/591 was used to evaluate the LPO in the spermatozoa, and 5 μΙ_ of its working solution was added to the sample, which was incubated in a water bath (37 °C/30 min). Then, 0.5 mL of PBS was added slowly through the tube wall. The sample was homogenized and then centrifuged (100 x g/5 min) to remove the unbound fluorochrome residues. The supernatant was discarded, and the pellet was re-suspended in 40 μΙ_ of PBS. Then, the samples were evaluated: orange-stained cells (C11-BODIPY-) were classified as non-peroxidized and green-stained cells (C11-BODIPY +) were classified as peroxidized (Figure 4).
The association of the fluorochromes M540 and Yo-Pro-1 was used to evaluate the membrane stability of spermatozoa. To that end, 5 μΐ of the Yo-Pro-1 solution was added to the sample and incubated in a water bath (37 °C/15 min). Then 5 μΐ of M540 was added to the sample, which was incubated for 5 min at room temperature. Three populations were identified: viable cells with stable membranes (M540- / Yo-Pro-1-); viable cells with destabilized membranes (M540 + / Yo-Pro-1-); and non-viable cells (YP +). Results were expressed as a percentage of viable cells with stable (M540-) or destabilized membranes (M540 +) (Figure 5).
Semen evaluations were performed using ImageStream® X, Mark II version (Amnis, Seattle, WA, USA). Data acquisitions were performed using INSPIRE® software, version 200.1.388.0 (Amnis, Seattle, WA, USA). The spermatozoa in focus were separated by a histogram of the RMS gradient plotted against the normalized frequency, and the single cells were identified by means of a plot of the area against the aspect ratio, which were verified by the images of the gallery of the clear field channel. All the fluorochromes were excited by a 488 nm laser. For the FITC-PNA + PI association, the laser power used was 55 mW, where FITC-PNA was detected by channel 2 (505 - 560 nm) and PI was detected by channel 5 (640 - 745 nm). For JC-1, the laser power was 130 mW, and detection was performed on channels 2 and 4 (595 - 640 nm). For DCFDA + PI, the laser power was 80 mW and channels 2 and 5 were used, respectively. For C11-BODIPY, the laser power was 60 mW and channels 2 and 4 were used, respectively. Finally, for the association of M540 + Yo-Pro-1, the laser power was 100 mW, and detection was performed with channels 3 (560-595 nm) and 2, respectively. In each of the samples, 5000 events were acquired. IDEAS software version 6.0 (Amnis, Seattle, WA, USA) was used for data analysis. All samples were analysed, and populations were divided by dot charts and histograms, and by visual confirmation of cell images.
The SAS GLIMMIX procedure (System for Windows, version 9.3; SAS Institute Inc., Cary, NC, USA) was used with a generalized linear modelling methodology. For TM, PM, LIN, STR, PNA- / PI-, PNA-, HMMP, DCFDA-, C11-BODIPY-, M540- / Yo-Pro-1-, and M540 + / Yo-Pro-1-, the authors used a binomial distribution with a logit link function. For VCL, VSL, VAP, ALH and BCF, the authors used a Gaussian distribution with an identification link function. Variables were analysed using a mathematical model that included the fixed effects of the treatment. The residual method was used to calculate the denominators of degrees of freedom to approximate the F tests in the mixed models. All statistical comparisons were performed using means adjusted by the least squares method and results are presented as mean squares ± standard error with the distributions shown in their original scales to aid interpretation. Tests with P <0.05 were considered significant.
Results and Discussion
Studies have demonstrated the effects of myricetin on functional aspects of human (Aquila et al., 2013) and ovine (Arruda et al., 2018) spermatozoa. In the present study, microscopic analysis (CASA) and flow cytometry were performed to evaluate the impact of the addition of various myricetin concentrations on kinematics, plasma and acrosome membrane integrity, mitochondrial membrane potential, intracellular ROS levels, lipid peroxidation and membrane stability of ovine spermatozoa in vitro.
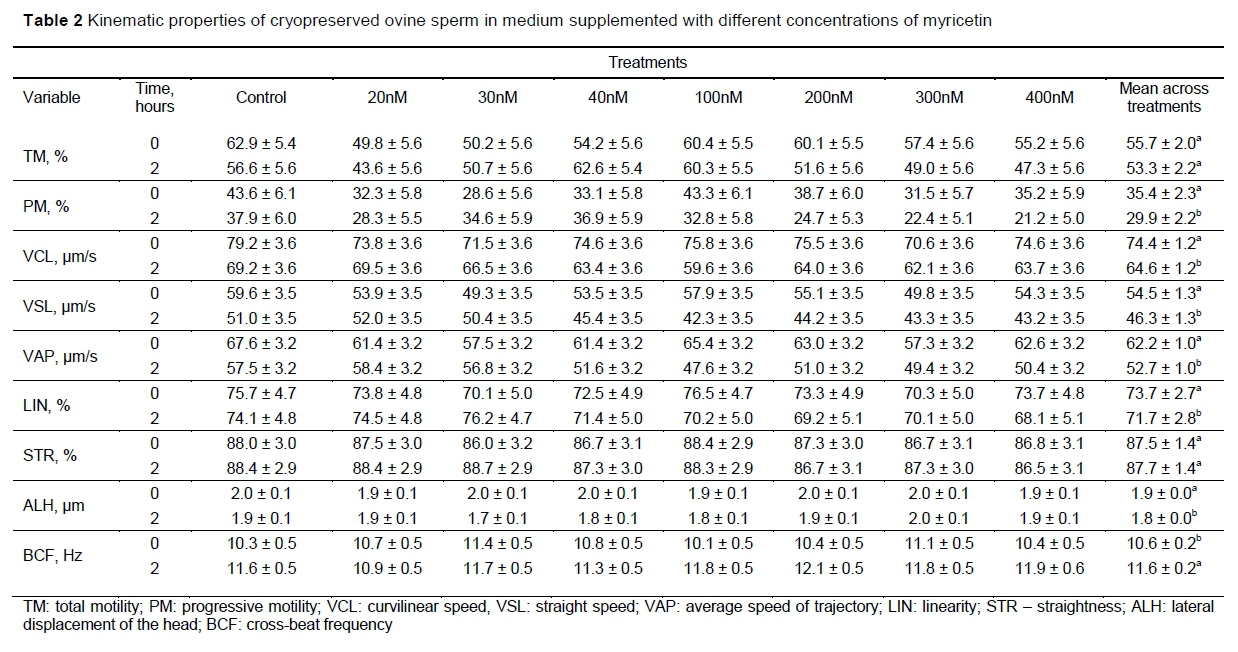
The results of the kinematic analyses are summarized in Tables 1 and 2. There were no interactions between treatment and time (P >0.0965) (Table 1), and no effect of treatments (P >0.2113) (Table 1) was detected on any of the measured variables.
As shown in Table 2, the PM, VCL, VSL, VAP, LIN and ALH values were reduced over time (P <0.0359), while the overall BCF mean increased (P =0.0001). The evaluation of the kinematic properties of spermatozoa has great importance in predicting fertilizing capacity of these gametes through the correlation between their displacement velocity and the success of in vitro and in vivo oocyte fertilization (Cox et al., 2006). In this study, the in vitro results did not demonstrate a positive effect of the addition of myricetin to the frozen-thawed ovine spermatozoa when the parameters were evaluated immediately after thawing.
It is essential to perform these analyses after an incubation period, when these gametes seem to have a higher correlation with fertility than in the evaluations performed immediately after thawing (Del Olmo et al., 2013). These analyses were also repeated after two hours of sperm incubation (at 37 °C), but no difference was observed between the control and myricetin-treated groups.
Comparing the effect of time on each sperm parameter, total motility (TM) and rectilinearity (STR) remained stable after two hours of incubation. However, there was a reduction in the progressivity (PM and LIN) and velocity (VCL, VSL, and VAP) of the gametes, in the general mean of the time, after two hours of incubation, parameters correlated with sperm fertility (Verstegen et al., 2002; Del Olmo et al., 2013). The general mean of BCF increased during the incubation period, which could imply in better migration capacity and penetration of the spermatozoa in the cervical mucus (Mortimer, 2000), whereas LIN, a parameter also related to the efficiency of sperm migration in the cervical mucus (Mortimer, 2000), decreased after two hours of incubation.
In an experiment conducted by Arruda et al. (2018), which evaluated ram semen immediately after thawing, 10 nM of myricetin was found to reduce the percentage of fast cells and mean path velocity (VAP), and 10, 100, and 1000 nM of myricetin increased BCF, with no beneficial effect on any of the other parameters evaluated immediately after thawing. Kinematic differences between the data of Arruda et al. (2018) and the results of this study may have occurred because of some modifications in the protocol, such as the preparation of the semen cryopreservation extender, dilution process, time from semen processing to freeze, incubation before the analyses in the computerized system (CASA) and evaluation after the incubation period.
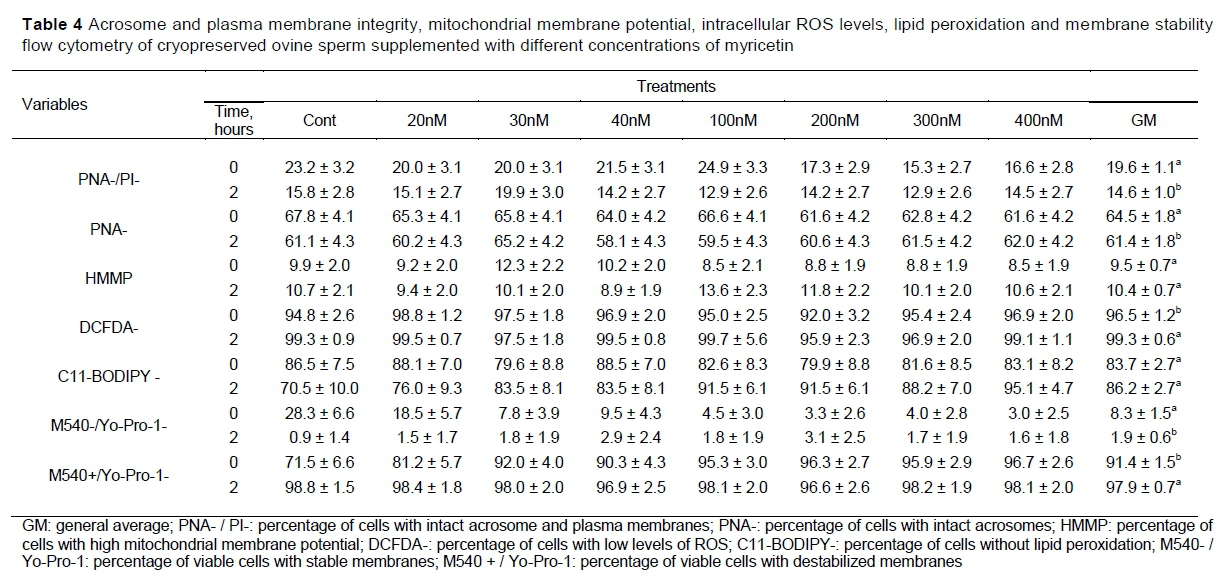
Results pertaining to flow cytometry are presented in Tables 3 and 4. There was no interaction between treatment and time (P >0.4109), and the treatments had no effect (P >0.5185) on the variables.
The overall mean of time (fixed time effect) on percent intact plasma membrane and acrosome membrane (PNA-/PI-), intact acrosome membrane (PNA-), cells with low levels of ROS (DCFDA-) and cells with viable membranes (M540-/Yo-Pro-1) reduced (P <0.0335) (Table 4) during incubation.
Aquila et al. (2013), tested the phytoestrogens effect of myricetin on human sperm (semen in natura), observing that an exposure to different concentrations of myricetin (10, 100, and 1000 nM) was able to alter motility and viability, as well as to cause biochemical changes associated with sperm capacitation, such as cholesterol efflux and tyrosine phosphorylation. In addition, these authors observed that myricetin increased acrosine activity and glucose utilization, demonstrating that sperm function was potentiated through the oestrogen receptor. In contrast, although cholesterol efflux and acrosin activity have not been evaluated, in the present study, it no changes were observed in membrane stability and acrosome integrity in the groups treated with myricetin. In this sequence of findings, it was evident that, in the human and ovine species, the various tested concentrations of myricetin have different effects on spermatozoa under the conditions in which they were evaluated.
Likewise, flow cytometery revealed no positive effect of adding myricetin to ovine semen on the integrity or stability of sperm membranes, or on aspects related to oxidative stress. High levels of ROS can affect the integrity of sperm membranes and increase lipid peroxidation, which results in loss of viability, reduction of motility and loss of plasma and acrosome membrane integrity, leading to a reduction in the quality of these cells and, consequently, a reduction of their fertilizing capacity (Alvarez & Storey, 1992; Aitken, 1995; Peris et al., 2007).
However, owing to the scarcity of studies using myricetin in semen extender, the results of this study can be related to those of quercetin, a flavonoid that is structurally similar to myricetin. Seifi-Jamadi et al. (2016), demonstrated that there were no positive effects using quercetin to improve viability and reduce oxidative stress after freezing equine semen. Likewise, Silva et al. (2016) did not observe an effect of quercetin on plasma and acrosome membrane integrity, or on oxidative stress in goat spermatozoa after cryopreservation. The action of these compounds is visible only in situations of extreme stress, which are observed only after induction of oxidation and do not reflect what occurs during cryopreservation (Silva et al., 2016).
The absence of a positive effect regarding the addition of various concentrations of myricetin in this in vitro study may be because of sufficient physiological protection conferred by the antioxidants found in the seminal plasma after no increase in ROS production was observed from the freezing- thawing process. Furthermore, the results obtained with myricetin in the present study can be attributed to the lack of a protective effect of the antioxidant on the physiological conditions (low levels of ROS) that the semen was exposed. These results suggest a need for more in vitro and in vivo studies such as (the evaluation of the phyto-estrogenic and inhibitory effect of ATPase) using other concentrations of myricetin.
Conclusion
Myricetin supplementation of the extender used for freezing ram semen did not produce antioxidant effects.
Acknowledgments
To the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (FACEPE) for granting a scholarship for PhD, the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for financial support.
Author Contributions
LCPA: conceptualization, data curation, investigation, methodology, writing original draft; RAJAS: data curation, methodology, writing review and editing; FCCM: data curation, investigation; MMM: data curation, investigation; JASC: data curation, investigation; AMB: conceptualization, methodology, formal analysis, writing review and editing, PLJMJ: formal analysis; DCMS: data curation, writing review; MMPG: project administration, funding acquisition, supervision, writing review and editing
Conflict of Interest Declaration
All the authors declare not have any conflict of interest.
References
Agarwal, A., Virk, G., Ong, C. & Du Plessis, S. S. 2014. Effect of oxidative stress on male reproduction. World J. Mens Health, 32(1), 1. DOI: 10.5534/wjmh.2014.32.1.1 [ Links ]
Aitken, R.J. 1995. Free radicals, lipid peroxidation and sperm function. Reprod. Fertil. Dev. 7(4),659-668. DOI:10.1071/RD9950659 [ Links ]
Aitken, R.J., Gibb, Z., Baker, M.A., Drevet, J. & Gharagozloo, P. 2016. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 28(2), 1. DOI: 10.1071/rd15325 [ Links ]
Alvarez, J.G. & Storey, T., 1992. Evidence for increased lipid peroxidative damage and loss of superoxide dismutase activity as a model of sublethal cryodamage to human sperm during cryopreservation. J. Androl. 13(3), 232-241. DOI: 10.1002/j.1939-4640.1992.tb00306.x [ Links ]
Amidi, F., Pazhohan, A., Shabani Nashtaei, M., Khodarahmian, M. & Nekoonam, S. 2016. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank 17(4), 745-756. DOI: 10.1007/s10561-016-9566-5 [ Links ]
Aquila, S., Santoro, M., De Amicis, F., Guido, C., Bonofiglio, D., Lanzino, M., Cesario, M.G., Perrotta, I., Sisc,i D. & Morelli, C. 2013. Red wine consumption may affect sperm biology: The effects of different concentrations of the phytoestrogen myricetin on human male gamete function. Mol. Reprod. Dev. 80(2), 155-165. DOI:10.1002/mrd.22145 [ Links ]
Arruda, L.C.P., Silva, R.A.J.A., Monteiro, M.M., Silva, R.P.F., Oliveira, A.S., Mergulhão, F.C.C., Monteiro Jr, P.L.J., Batista, A.M. & Guerra, M.M.P 2018. Avaliação in vitro do sêmen congelado de carneiros com diluidor suplementado com miricetina. Arq. Bras. Med. Veterinária e Zootec. 70(1), 153-159. DOI: 10.1590/1678-41629671 [ Links ]
Barreiros, A.L.B.S., David, J.M. & David, J.P. 2006. Estresse oxidativo: Relação entre geração de espécies reativas e defesa do organismo. Quim. Nova. 29(1), 113-123. DOI: 10.1590/S0100-40422006000100021 [ Links ]
Chobot, V. & Hadacek, F. 2011. Exploration of pro-oxidant and antioxidant activities of the flavonoid myricetin. Redox Rep. 16(6), 242-247. DOI: 10.1179/1351000211y.0000000015 [ Links ]
Cox, J.F., Alfaro, V., Montenegro, V. & Rodriguez-Martinez, H. 2006. Computer-assisted analysis of sperm motion in goats and its relationship with sperm migration in cervical mucus. Theriogenology 66(4), 860-867. DOI: 10.1016/j.theriogenology.2006.01.062 [ Links ]
Cross, N.L. & Hanks, S.E. 1991. Effects of cryopreservation on human sperm acrosomes. Hum. Reprod. 6(9),1279-1283. DOI: 10.1093/oxfordjournals.humrep.a137526 [ Links ]
Del Olmo, E., Bisbal, A., Maroto-Morales, A., García-Alvarez, O., Ramon, M., Jimenez-Rabadan, P., Martínez-Pastor, F., Soler, A.J., Garde, J.J. & Fernandez-Santos, M.R. 2013. Fertility of cryopreserved ovine semen is determined by sperm velocity. Anim. Reprod. Sci. 138(1-2),102-109. DOI: 10.1016/j.anireprosci.2013.02.007 [ Links ]
Hammerstedtl, R.H. 1993. Maintenance of bioenergetic balance in sperm and prevention of lipid peroxidation: A review of the effect on design of storage preservation systems. Reprod. Fertil. Dev. 5(6), 675-690. DOI:10.1071/RD9930675 [ Links ]
Hendrich, A.B. 2006. Flavonoid-membrane interactions: Possible consequences for biological effects of some polyphenolic compounds. Acta Pharmacol. Sin. 27(1),27-40. DOI: 10.1111/j.1745-7254.2006.00238.x [ Links ]
Huang, H., Chen, A.Y., Rojanasakul, Y., Ye, X., Rankin, G.O. & Chen, Y.C. 2015. Dietary compounds galangin and myricetin suppress ovarian cancer cell angiogenesis. J. Funct. Foods 15, 464-475. DOI: 10.1016/j.jff.2015.03.051 [ Links ]
Huang, J.H., Huang, C.C., Fang, J.Y., Yang, C., Chan, C.M., Wu, N.L., Kang, S.W. & Hung, C.F. 2010. Protective effects of myricetin against ultraviolet-B-induced damage in human keratinocytes. Toxicol. Vitr. 24(1),21-28. DOI: 10.1016/j.tiv.2009.09.015 [ Links ]
Kimura, A.M., Tsuji, M., Yasumoto, T., Mori, Y. Oguchi, T., Tsuji, Y., Umino, M., Umino, A., Nishikawa, T., Nakamura, S., Inoue, T., Kiuchi, Y., Yamada,M., Teplow, D.B. & Ono, K., 2021. Myricetin prevents high molecular weight Aß1-42 oligomer-induced neurotoxicity through antioxidant effects in cell membranes and mitochondria. Free Radic. Biol. Med. (In press). https://pubmed.ncbi.nlm.nih.gov/34015458/ [ Links ]
, Maia, M.S., Bicudo, S.D., Azevedo, H.C., Sicherle, C.C., Souza, D.B. & Rodello, L. 2009. Motility and viability of ram sperm cryopreserved in a Tris-egg yolk extender supplemented with anti-oxidants. Small Rum. Res. 85(2), 85-90. DOI: 10.1016/j.smallrumres.2009.07.001 [ Links ]
Mogielnicka-Brzozowska, M., Strzezek, R., Wasilewska, K. & Kordan, W. 2015. Prostasomes of canine seminal plasma - zinc-binding ability and effects on motility characteristics and plasma membrane integrity of spermatozoa. Reprod. Domest. Anim. 50(3), 484-491. DOI: 10.1111/rda.12516 [ Links ]
Mortimer, S.T. 2000. CASA - practical aspects. Andrology Lab Corner. J. Androl. 21(4), 515-24. DOI: 10.1002/j.1939-4640.2000.tb02116.x [ Links ]
Naseer, Z., Ahmad, E., Aksoy, M., Küçük, N., Serin, I., Ceylan, A., Boyacioglu, M. & Kum, C. 2015. Protective effect of cholesterol-loaded cyclodextrin pretreatment against hydrogen peroxide induced oxidative damage in ram sperm. Cryobiology 71(1),18-23. DOI: 10.1016/j.cryobiol.2015.06.007 [ Links ]
Peris, S.I., Bilodeau, J.F., Dufour, M. & Bailey, J.L. 2007. Impact of cryopreservation and reactive oxygen species on DNA integrity, lipid peroxidation, and functional parameters in ram sperm. Mol. Reprod. Dev. 74(7),878-892. DOI:10.1002/mrd.20686 [ Links ]
Seifi-Jamadi, A., Kohram, H., Zare Shahneh, A., Ansari, M. & Macías-García, B. 2016. Quercetin ameliorate motility in frozen-thawed Turkmen stallions sperm. J. Equine Vet. Sci. 45,73-77. DOI: 10.1016/j.jevs.2016.06.078 [ Links ]
Silva, E.C.B., Arruda, L.C.P., Silva, S.V., Souza, H.M. & Guerra, M.M.P. 2016. High resveratrol or quercetin concentrations reduce the oscillation index of frozen goat semen. Arq. Bras. Med. Veterinária e Zootec. 68(5),1237-1243. DOI: 10.1590/1678-4162-8670 [ Links ]
Thiyagarajah, P., Kuttan, S.C., Lim, S.C., Teo, T.S. & Das, N.P. 1991. Effect of myricetin and other flavonoids on the liver plasma membrane Ca2+ pumP kinetics and structure-function relationships. Biochem. Pharmacol. 41(5), 669-675. DOI: 10.1016/0006-2952(91)90065-D [ Links ]
Verstegen, J., Iguer-Ouada, M. & Onclin, K. Computer assisted semen analyzers in andrology research and veterinary practice. Theriogenology. 2002; 57(1):149-179. DOI: 10.1016/S0093-691X(01)00664-1 [ Links ]
Yan, T., Tao, Y., Wang, X., Lv, C., Miao, G., Wang, S., Wang, D. & Wang, Z., 2021. Preparation, characterization and evaluation of the antioxidant capacity and antitumor activity of myricetin microparticles formated by supercritical antisolvent technology. J. Supercrit. Fluid. (In press). DOI: 10.1016/j.supflu.2021.105290 [ Links ]
Submitted 27 May 2020
Accepted 7 July 2021
Published 15 February 2022
# Corresponding author: luciacpa@hotmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}