










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 n.6 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i6.12
Estimates of variance components for feedlot traits of the Simmentaler breed in South Africa
J. HendriksI, II; F.W.C. NeserII; J.B van WykII; F.J. JordaanI, II; M.M. ScholtzI, II, #
IARC-Animal Production, Private Bag X2 Irene 0062, South Africa
IIUniversity of the Free State, PO Box 33009, Bloemfontein, 9300, South Africa
ABSTRACT
Breeding of beef cattle is changing, with more emphasis on efficiency of production. Feed cost is the highest expense, and reducing it has the potential to increase profitability. Common measures of efficiency are ratio traits such as feed conversion (feed consumed/weight gain) and feed efficiency (weight gain / feed consumed). Feed conversion ratio is commonly used in South Africa in an attempt to improve feed efficiency. These ratio traits are associated with growth rate. Selection for them would result in higher growth as a correlated response and might also increase the mature size of the cows and their maintenance cost. Thus, alternative efficiency traits such as residual feed intake and residual daily gain have been proposed. In this study, variance components, and genetic parameters for feedlot traits for the South African Simmentaler breed were estimated, with emphasis on the efficiency traits. The focus was to evaluate the use of residual feed intake as an alternative trait. The results indicate non-significant correlations between residual feed intake and body weight and growth traits, implying that residual feed intake should have little effect on the other traits. This is in contrast to the moderate to strong correlations of feed conversion ratio with the same traits. The study demonstrates that considerable genetic variation exists for residual feed intake, which can be exploited. Selection for residual feed intake can reduce the carbon footprint of beef due to the associated lower methane emissions.
Keywords: feed conversion ratio, heritability, residual feed intake, weight
Introduction
Feed cost is extremely important in beef production since it has a major influence on profitability (Koch et al., 1963; Parnell et al., 1994). It has been shown that a 10% improvement in feed efficiency can improve profitability more than twice as much as when a 10% improvement in growth was achieved (Fox et al., 2004). In addition, the conversion of feed into human food is an important role of livestock production. If the efficiency of this process can be improved, livestock production will become more sustainable. Increasing efficiency would lead to an increase in production of nutritious food for people from a limited resource (MacNeil et al., 2013).
Common measures of efficiency are feed conversion ratio (FCR) (feed consumed / weight gain) and feed efficiency (weight gain / feed consumed), which are ratio traits, with FCR still being used to select for efficiency. However, efficiency is highly associated with growth rate, thus when selecting for better efficiency, indirect selection for higher growth takes place (Koch et al., 1963). This can lead to an increase in mature size, which increases the maintenance cost of the breeding herd (Koots et al., 1994). Another problem when selecting for ratio traits is that the change that occurs in the component traits (growth and feed intake) is unknown (Arthur et al., 2001).
Koch et al. (1963) first described efficiency traits in which efficiency was computed as i) feed consumption adjusted for differences in gain, and ii) gain adjusted for differences in feed consumption. The first trait is now referred to as residual feed intake (RFI) and is defined as the difference between the actual feed intake and the expected feed intake of an animal to fulfil its maintenance requirements and growth. It is sometimes also referred to as net feed intake. The second trait is referred to as residual daily gain (RDG) (Arthur et al., 2001, MacNeil et al., 2013, Retallick, 2013). Residual feed intake differs from ratios composed of feed intake and gain in that it is independent of growth and maturity patterns (Sainz & Paulino, 2004). In a study conducted by Smith et al. (2010), high RFI bulls consumed 17.3% more feed than low RFI bulls. Lancaster et al. (2009) also found that high RFI bulls consumed 18% more feed. Factors that contribute to variations in RFI between animals include differences in protein turnover, tissue metabolism, stress, and heat increment of fermentation (Richardson & Herd, 2004). Castro et al. (2006) indicated that dry matter intake (DMI) was correlated positively with RFI (r =0.44, P <0.001). This showed that selection for RFI could be used as a way to reduce input costs in beef production, as animals with low RFI eat less than animals with high RFI. Selection for reduced RFI would have the advantage not only of reduced feeding costs but, since it is unrelated to growth rate, would not lead to a larger mature size, especially in cows.
Another big concern of beef production is the contribution to global warming. Selection for feed efficiency through RFI, where animals consume less feed than expected, would not only improve feed efficiency, but would lead to lower methane emissions, since there is a positive correlation between RFI and methane production, thus reducing the carbon footprint of beef cattle (Nkrumah et al., 2006). Selection for (lower) RFI differs from other feed efficiency traits such as FCR (feed consumed/weight gain) and feed efficiency (weight gain/feed consumed), as it appears to be independent of bodyweight. It should thus not lead to increased maintenance requirements in mature animals, which would ultimately limit the carbon footprint. Any increase in mature cow weight, especially under extensive primary beef production systems, as encountered in South Africa and many other tropical and subtropical countries, would result in higher cow maintenance requirements, and thus increased release of enteric methane into the atmosphere.
Nkrumah et al. (2006) reported that beef cattle with low RFI produced up to 28% less methane than those with high RFI. Residual feed intake is calculated as the difference between actual feed intake and the expected feed requirements for maintenance of bodyweight and a certain level of production (Hegarty et al., 2007). The lower methane production was attributed to differences in the ruminal microbial population between animals, and these differences could be heritable (Nkrumah et al., 2006).
The Simmentaler is the most numerous European breed in South Africa (Scholtz, 2010). Since individual feed intake had already been measured for the breed in the central growth tests of the National Beef Recording and Improvement Scheme in South Africa, it was possible to calculate RFI from the existing information. The aim of this study was to estimate genetic variance and co-variance components with the aim of estimating breeding values for feedlot traits.
Material and Methods
The data of the South African Simmentaler breed for this study were obtained from the South African INTERGIS (Integrated Registration and Genetic Information System), which stores all data from the National Beef Recording and Improvement Scheme. Data for this scheme were collected according to approved standard operating procedures and were accredited with the International Committee for Animal Recording (ICAR).
Weaning weights were recorded on-farm and post-weaning performance from the young Simmentaler bulls at centralized growth test stations. This information was used to calculate the variance components of 10 pre-weaning weight, post-growth, feed efficiency and body measurement traits. The data were collected over 40 years. In the centralized tests animals were subjected to an adaptation period of 28 days, after which they were tested for 84 to 140 days after weaning. The data for this study were restricted to the first 84 days of the test. Data were collected from the Agricultural Research Council (ARC) testing stations at Irene near Pretoria (Gauteng), Armoedsvlakte at Vryburg (North West) Glen Agricultural College near Bloemfontein (Free State), Cedara, near Pietermaritzburg (KwaZulu-Natal), Stellenbosch (Western Cape), and a number of private test stations. These test stations are distributed throughout South Africa.
The bulls were tested under partly controlled environments, and much of the data were collected from animals receiving a 'standardized' diet. All test stations under the control of the ARC used a diet with an energy requirement of approximately 11 MJ/kg DM and between 13.5% and 15% crude protein. Factors that might have an influence on performance were test station, the age of the animals, body condition at start of the test and the season in which the test took place.
During the first 14 days of adaption, the intake was restricted to ensure adequate time for the animals to adapt to the ration, and in the last 14 days of adaptation the animals were fed ad libitum. During the adaptation period, the animals were 'taught' to open their gates to gain access to their own feed troughs, since the Calan gate system was used to collect feed intake data.
Traits that were included in the study were weaning weight (WW), end of test weight (ETW), ADG, daily feed intake (DFI), FCR, RFI, RDG, metabolic mid-test weight (MWt), shoulder height (SH), and scrotal circumference (SC). The equations for RFI and RDG were determined using the test data collected at centralized test stations.
Average daily gain was calculated using linear regression in SAS (SAS Institute Inc., Cary, North Carolina, USA). Weekly weights were regressed against time (measured in days) to give the measure of ADG as the amount of growth (in kg) per day. Feed intake was expressed as the average amount of feed consumed per day during the test, where DFI was calculated by dividing the total amount of feed consumed during the test by the test length. Feed conversion ratio was calculated by dividing DFI by ADG. Residual feed intake was calculated by regressing DFI on ADG and MWT. These regression coefficients were then used to estimate each animal's intake according to its needs for growth (ADG) and maintenance, as measured by MWT. Metabolic mid-test weight was calculated by taking the average between the starting and end weight of test to the power 0.75. This represented the amounts of active tissue in the body using energy and of energy required by the animal for maintenance when at complete rest. Residual daily gain represents a growth trait where ADG is regressed on DFI, which gives an estimate of the expected growth rate of an animal for a specific level of intake. The expected growth rate is subtracted from the actual growth rate and the difference is the RDG value (MacNeil et al., 2013). Animals with a positive value will grow more than expected for a certain level of intake and vice versa. Phenotypic values for RFI and RDG were calculated using these regression equations:
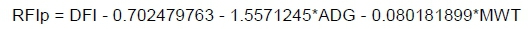
and
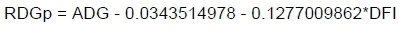
Contemporary groups for the traits measured during the post-weaning growth tests were defined as test centre, test year and test number. For weaning weight, contemporary groups were defined by birth year and season of birth. Only animals belonging to a contemporary group of five or more were included in the analysis. From the initial 4119 growth test records, 941 records were removed, and a total of 3178 records were available for analysis after editing. There were 361 contemporary groups and the average number of animals per contemporary group was 8.8. A total of 63 590 weaning weight records were also used in the analysis. The descriptive statistics are indicated in Table 1. The regression coefficients for the RFI and RDG equations were determined using SAS (SAS Institute Inc., Cary, North Carolina, USA), with contemporary group included in the models set as a class variable.
The following random effects were tested for inclusion in all traits using the log likelihood test (Swalve, 1993): direct additive, maternal additive, permanent environmental effect due to animal and contemporary group. The GLM procedure in SAS (SAS Institute Inc., Cary, North Carolina, USA) was used to examine alternative fixed effects. Age and weight at start of test were included as covariates for ADG, DFI, FCR, RDG, ETW and MWT. Final weight and age at end of test were included as covariates for SH and SC. Contemporary group was regarded as a fixed effect for all the traits. In weaning weight, calf age at weaning and age of the dam were included as covariates, whereas sex and contemporary group were modelled as fixed effects. The fixed and random effects that were included in the models for these traits are summarized in Table 2. The ASREML (Gilmour et al., 1999) program was used to obtain estimates of the (co)variance components. The log likelihood test indicated that the covariance between the direct additive and maternal additive effects was not significant for weaning weight. These final models were used in both the univariate and bivariate analyses for weaning weight and traits in the central growth tests:
WW:
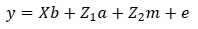
ETW, ADG, DFI, FCR, RFI, RDG, MWT, SH and SC:
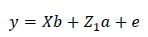
where: y = a vector of observations,
= a vector of fixed effects,
= a vector of direct additive genetic effects,
= a vector of maternal additive effects,
= a vector of residuals,
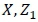 and
and 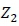 = incidence matrices relating observations to their respective fixed and random effects.
= incidence matrices relating observations to their respective fixed and random effects.
It was assumed that 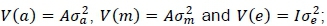 where Iis an identity matrix,
where Iis an identity matrix, 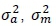 , and
, and 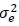 are the direct additive, maternal additive and environmental variance components, respectively.
are the direct additive, maternal additive and environmental variance components, respectively.
Results and Discussion
The best models were determined with the restricted maximum likelihood test, which accounts for the loss of degrees of freedom when estimating fixed effects (Harville, 1977). Some difficulty was encountered in obtaining convergence of the analyses, perhaps because of the lack of connectedness among animals in the contemporary groups (MacNeil et al., 2018). Weaning weight was included in all models for the post-weaning traits to remove the potential for selection bias in the sampling of bulls that entered the test centres.
Multiple heritability estimates were available for each trait, since bi-variate models were used to calculate covariance components and correlations among the traits. The weighted means procedure described by Koots et al. (1994) was used to combine the estimates of heritability, genetic correlation, and their standard error. The heritability estimates varied from 0.14 for weaning weight maternal up to 0.42 for shoulder height. All the heritability estimates had relatively small standard errors (genetic parameter larger than twice the standard error) and varied within the range that is reported in the literature.
The heritability estimates for direct and maternal genetic effects on WW were similar (Table 3). It was expected that the additive effect would have a higher heritability than the maternal effect. However, Arthur et al. (2001) also found the heritability estimates of WW direct and WW maternal were similar: 0.17 and 0.13, respectively. The heritability estimates for the postweaning weight traits varied from 0.24 for ETW to 0.23 for MWT, which is in line with previous estimates, which ranged from 0.19 to 0.42 as reported by Afolayan et al. (2007).
Average daily gain is reported to be a moderately heritable trait, with heritability estimates ranging between 0.16 and 0.41 (Herd & Bishop, 2000; Arthur et al., 2001; Herring, 2003; Schenkel et al., 2004; Crowley et al., 2010; Caetano et al., 2013). In in this study, the heritability of ADG (0.26) for Simmentaler was towards the lower end of this range (Table 3). Van der Westhuizen et al. (2004) analysed Bonsmara bulls tested at the same stations and found a heritability of 0.37 for ADG.
Numerous studies have reported that growth is a heritable trait, with estimates of heritability varying between 0.26 and 0.62 (Koch et al., 1963; Rolfe et al., 2011) The heritability estimate for ADG in this study was 0.26, which is at the lower end of the reported heritability estimates. The heritabilities of the traits associated with efficiency were 0.21 for FCR, 0.26 for RFI, and 0.20 for RDG, which are all moderately heritably and in line with the heritabilities of 0.19 - 0.55 for efficiency traits, as reported by Torres-Vazquez et al. (2018). The heritability of DFI was 0.30, which was also in line with the ranges of 0.28 to 0.44 reported in the literature (Koch et al., 1963; Schenkel 2004).
The heritability of SC was within range of what would be expected, based on other studies, which found heritability estimates that varied between 0.39 and 0.44 (Arthur et al., 2001; Boligon et al., 2011). In SH, the heritability was 0.42, but none of the correlations was significant. Since SH is an indication of frame size and thus mature size, it would be expected that ADG would have a positive correlation with SH. Van der Westhuizen et al. (2004) found a moderate correlation of 0.48 between ADG and SH. This indicated that selection for ADG would increase frame size and thus mature weight.
Metabolic mid-test weight (MWT) also had a positive genetic correlation with DFI (0.41), indicating that, as intake levels rise, the bodyweight of the animals will also be higher. This was expected since higher levels of intake should lead to higher growth rates. Positive genetic correlations were also reported in the literature, varying from 0.39 to 0.55 (Arthur et al., 2001; Crowley et al., 2010). Residual daily gain, which is an indication of efficiency of growth according to the level of intake, was strongly correlated with other growth and bodyweight traits, namely ADG (0.81), MWT (0.83), and ETW (0.65). Since ADG is regressed on DFI, one can expect that RDG and DFI would be independent. This was supported by the non-significant genetic correlation of 0.04. The correlation between FCR and RDG was not estimable, whereas the correlation between DFI and FCR was close to zero (-0.08), indicating that DFI did not contribute to the variation found in FCR.
For many years, FCR was the most commonly used trait to select for efficiency. Unfortunately, it affected both metabolic efficiency and growth (indirect selection for better growers), which might increase the mature size of the animals through generations of selection (McGee, 2014). It was therefore important to identify alternative traits. Residual feed intake (RFI) is independent of growth rate and bodyweight and thus measures only metabolic efficiency (Kerley, 2010). Therefore, selection for RFI should not have an effect on growth rate or body size. In this study a negative correlation was found between FCR and ADG (-0.76). From the literature, the correlation was found to range between -0.46 and -0.62 (Arthur et al., 2001 ; Crowley et al., 2010). The correlation in this study was higher than that found in the literature, which was supporting evidence of the antagonism that selection for FCR held.
The genetic correlation between RFI and ADG in this study was non-significant (0.19 ± 0.16), indicating that there was no genetic correlation between these traits. The correlation between RFI and ETW (-0.26 ± 0.16) or MWT (0.19± 0.16), which are both measures of bodyweight, was also low. This implies that selection for RFI should not have an effect on growth rate or mature weight over time, as was reported in the literature (Herd & Bishop, 2000; Arthur et al., 2001; Van der Westhuizen et aí., 2004). Because FCR is strongly correlated with MWT and ETW (-0.86 and -0.89 respectively), this implies that FCR has a strong association with traits influenced by growth, which might indicate that ADG contributed to most of the variation found in FCR. These strong negative correlations indicated that animals with good FCR values would have higher bodyweights.
Since RFI and FCR are both efficiency traits aimed at lowering intake, it was expected that they would have a positive correlation, as reported in other studies in which the genetic correlation varied between 0.45 and 0.85 (Arthur & Herd, 2008). The genetic correlation of 0.51 found in this study indicated that animals with better FCR values might be those with good RFI values. This correlation is similar to the phenotypic correlation obtained by Arthur et al. (1997). Because the correlation was not very strong (0.51), this indicated that it was not only animals with better FCR values that had good RFI values. Since FCR is a ratio trait, and because better metabolic efficiency and better growth improve FCR, this presents a problem, since it is not possible to know the amount of selection pressure that is placed on each of the component traits.
Freetly et al. (2020) reported that selection for RFI in growing animals would probably have the same effect on mature cows. This implied that selection for reduced RFI would have the advantage of reduced feeding costs not only in growing animals, but also in breeding cows.
In this study the genetic correlation between RFI and RDG was -0.39. This was in agreement with the results of MacNeil et al. (2013), which found that animals for selection based on RFI and RDG provided quite different rankings in merit. These measures could be combined, as suggested by Berry & Crowley (2012), to produce a unified efficiency metric. Alternatively, a selection index may be an efficient approach to manage the antagonism between feed intake and growth rate (MacNeil et al., 2013)
In this study the genetic variance and covariance components for RFI and other efficiency traits have been estimated with the aim of obtaining their estimated breeding values (EBVs). However, a priori calculation of RFI followed by single-trait prediction of its EBV is not recommended, because this strategy compromises the value of the indicator traits, resulting in reduced accuracy of selection and increasing the possibility of selection bias (MacNeil, et al., 2011). The recommended approach is to do a genetic evaluation on the component traits with the EBVs for RFI and other efficiency traits being computed using the EBVs and (co)variance components feed intake, growth rate and other traits as needed, similar to what MacNeil et al. (2013) recommended for the Bonsmara breed.
The impact of global warming and continued excess production of greenhouse gases has twofold implications for the livestock industry. First, the continuous increase in ambient temperature is predicted to have both direct and indirect effects on animals and, second, the livestock industry has the responsibility to limit the carbon footprint to ensure future sustainability (Scholtz et al., 2021). Climate change projections for southern Africa are in agreement that the sub-continent will become drier and warmer, and the average temperature may increase by 1.5 °C to 2 °C, whereas the rainfall in certain areas will decrease by 40 mm per annum (Meissner et al., 2013). Climate-smart beef production is therefore essential, and this should include alternative breeding objectives for cow-calf and post-weaning efficiencies that can optimize it. Selection for alternative measures of post-weaning efficiency, such as RFI and RDG and in combination, or a selection index that includes feed intake and performance measures, is therefore important. Low RFI animals produce about 16 100 L/year less methane than high RFI animals (Nkrumah et al., 2006). Furthermore, selection for a trait such as RFI would not increase cow weight and thus maintenance requirements. Under the extensive conditions in South Africa this would support the improvement in cow-calf efficiency and thus sustainability.
Conclusion
It is desirable to increase efficiency of production and not production per se to ensure sustainability in the face of climate change. For example, the strong negative correlation between FCR and post-weaning growth and bodyweight traits indicated that growth and bodyweight would increase in the South African Simmentaler breed when selecting for FCR. These changes could be undesirable in a warmer and drier South Africa. The results from this study provided the necessary genetic parameters to estimate breeding values that could be used by Simmentaler breeders to manage the genetic antagonism of feed intake and gain. Selection for RFI has the potential to reduce the carbon footprint of beef owing to the associated lower methane emissions. With approximately four to six cattle generations before the full impact of climate change occurs, substantial progress can be made to dampen its negative effects if an appropriate breeding strategy is implemented.
Acknowledgements
This work is based on research supported in part by Red Meat Research and Development South Africa and the National Research Foundation of South Africa (NRF), under grant UID 75123. The grantholder acknowledges that opinions, findings and conclusions or recommendations expressed in any publication generated by NRF-supported research are those of the authors and that the NRF accepts no liability whatsoever in this regard.
Authors' Contributions
This study is based on the of the MSc dissertation of JH. MMS. FWCN and JBvW conceived the investigation. FJJ withdrew the data from the INTERGIS and assisted with data editing. JH prepared the data for analysis and analysed the data under the guidance of FWCN and MMS. FJJ, MMS, FWCN and JBvW contributed to interpretation of the results. JH drafted the initial manuscript, which was subsequently edited by MMS and FWCN. All authors read and approved the final manuscript.
Conflict of Interest Declaration
The authors declare that they have no conflicts of interest with regard to this work.
References
Afolayan, R.A., Pitchford, W.S., Deland, M.P.B. & McKiernan, W.A., 2007. Breed variation and genetic parameters for growth and body development in diverse beef cattle genotypes. Animal 1, 13-20. DOI: 10.1017/S1751731107257933 [ Links ]
Archer, J.A., Arthur, P.F., Herd, R.M., Parnell, P.F. & Pitchford, W.S., 1997. Optimum post weaning test for measurement of growth rate, feed intake, and feed efficiency in British breed cattle. J. Anim. Sci. 75,2024-2032. DOI: 10.2527/1997.7582024x [ Links ]
Arthur, P.F. & Herd, R.M., 2008. Residual feed intake in beef cattle. R. Bras. Zootec. v.37, suplemento especial. Pp. 269-279. DOI: 10.1590/S1516-35982008001300031 [ Links ]
Arthur, P.F., Archer, J.A., Herd, R.M. & Melville, G.J., 2001. Response to selection for net feed intake in beef cattle. Proc. Assoc. Adv. Anim. Breed. Genet. 14, 135-138. [ Links ]
Berry, D.P. & Crowley, J.J., 2012. Residual intake and body weight gain: A new measure of efficiency in growing cattle. J. Anim. Sci. 90, 109-115. DOI: 10.2527/jas.2011-4245 [ Links ]
Boligon, A.A., Baldi, F. & Albuquerque, L.G., 2011. Genetic parameters and relationships between growth traits and scrotal circumference measured at different ages in Nelorre cattle. Gen. Mol. Bio. 34, 225-230. DOI: 10.1590/s1415-47572011005000004 [ Links ]
Caetano, S.L., Savegnago, R.P., Boligon, A.A., Ramos, S.B., Chud, T.C. S.,Lobo, R.B. & Munari, D.P., 2013. Estimates of genetic parameters for carcass, growth and reproductive traits in Nellore cattle. Livest. Sci. 155, 1-7. DOI: 10.3390/ani9100715 [ Links ]
Castro, B.F.C., Paulino, P.V., Sanches, A.C. & Sainz, R.D., 2006. Growth, carcass quality, and protein and energy metabolism in beef cattle with different growth potentials and residual feed intake. J. Anim. Sci. 85, 928-936. DOI: 10.2527/jas.2006-373 [ Links ]
Crowley, J.J., McGee, M., Kenny, D.A., Crews Jr., D.H., Evans, R.D. & Berry, D.P., 2010. Phenotypic and genetic parameters for different measures of feed efficiency in different breeds of Irish performance-tested beef bulls. J. Anim. Sci. 88, 885-894. DOI: 10.2527/jas.2009-1852 [ Links ]
Fox, D.G., Tedeschi, L.O., Tylutki, T.P., Russell, J.B., van Amburgh, M.E., Chase, L.E., Pell, A.N. & Overton, T.R., 2004. The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion. Animal Feed Sci. Tech. 112, 29-78. DOI: 10.1016/j.anifeedsci.2003.10.006 [ Links ]
Freetly, H.C., Kuehn, L.A., Thallman, R.M. & Snelling, W.M., 2020. Heritability and genetic correlations of feed intake, body weight gain, residual gain and residual feed intake of beef cattle as heifers and cows. J. Anim. Sci. 98, 1-6. DOI: 10.1093/jas/skz394 [ Links ]
Gilmour, A.R., Cullis, B.R., Welham, S.J. & Thompson, R., 1999. ASREML, Biometric Bulletin 3, NSW Agriculture, Orange Agricultural Institute, Orange, NSW. [ Links ]
Harville, D.D., 1977. Maximum likelihood approaches to variance component estimation and to related problems. J. American Stat. Ass. 72, 320-338. DOI: 10.1080/01621459.1977.10480998 [ Links ]
Hegarty, R.S., Goopy, J.P., Herd, R.M. & McCorkell, B., 2007. Cattle selected for lower residual feed intake have reduced daily methane production. J Anim. Sci. 85, 1479-1486. DOI, 10.2527/jas.2006-236 [ Links ]
Herd, R.M. & Bishop, S.C., 2000. Genetic variation in residual feed intake and its association with other production traits in British Hereford cattle. Livest. Prod. Sci. 63, 111-119. DOI: 10.1016/S0301-6226(99)00122-0 [ Links ]
Herring, W.O., 2003. Genetics of feed efficiency in Angus cattle. Florida Beef Report. Pp. 31-34. [ Links ]
Kerley, M.S., 2010. Impact of selection for residual feed intake on forage intake by beef cows and feed efficiency of progeny. Division of Animal Sciences, University of Missouri. Pp. 39-49. [ Links ]
Koch, R.M., Swiger, L.A., Chambers, D. & Gregory, K.E., 1963. Efficiency of feed use in beef cattle. J. Anim. Sci. 22, 486-494. https://doi.org/10.2527/jas1963.222486x [ Links ]
Koots, K.R., Gibson, J.P., Smith, C. & Wilton, J.W., 1994. Analyses of published genetic parameter estimates for beef production traits. 1. Heritability. Anim. Breed. Abstr. 62, 309-338. [ Links ]
Lancaster, P.A., Carstens, G.E., Crews Jr., D.H., Welsh Jr., T.H., Forbes, T.D.A., Forrest, D.W., Tedeschi, L.O., Randel, R.D., & Rouquette, F.M., 2009. Phenotypic and genetic relationships of residual feed intake with performance and ultrasound carcass traits in Brangus heifers. J. Anim. Sci. 87, 3887-3896. DOI: org/10.2527/jas.2009-2041 [ Links ]
MacNeil, M.D., Lewis, R.M., & Scholtz, M.M., 2018. Degree of connectedness among herds and sires of current candidates for selection in Afrikaner cattle with reported weaning weights. Proc. World Cong. Genetics Appl. . Prod. Electronic Poster Session - Genetic Gain - Strategies for Local Breeds 2, 34, [ Links ]
MacNeil, M.D., Lopez-Villalobos, N. & Northcutt, S.L., 2011. A prototype national cattle evaluation for feed intake and efficiency of Angus cattle. J. Anim. Sci. 89:3917-3923. DOI: 10.2527/jas.2011-4124 [ Links ]
MacNeil, M.D., Scholtz, M.M. & Maiwashe, A., 2013, Estimates of variance components for post weaning feed intake and growth in Bonsmara bulls and evaluation of alternative measures of feed efficiency. S. Afr. J. Anim. Sci. 43, 20-24. DOI: 10.4314/sajas.v43i1.2 [ Links ]
McGee, M., 2014. Feed efficiency in beef finishing systems. Teagasc-IGFA Nutrition Conference, June 2014, Portaoise, Ireland. [ Links ]
Meissner, H.H., Scholtz, M.M. & Engelbrecht, F.A., 2013. Sustainability of the South African Livestock Sector towards 2050. Part 2: Challenges, changes and required implementations. South African Journal of Animal Science 43, 298-319. DOI: 10.4314/sajas.v43i3.6 [ Links ]
Nkrumah, J.D., Okine, E.K., Mathison, G.W., Schmid, K., Li, C., Basarab, J.A., Price, M.A., Wang, Z. & Moore, S.S., 2006. Relationships of feedlot feed efficiency, performance, and feeding behaviour with metabolic rate, methane production, and energy partitioning in beef cattle. J. Anim. Sci. 84, 145-153. DOI: 10.2527/2006.841145x [ Links ]
Parnell, P.F., Herd, R.M., Perry, D. & Bootle, B., 1994. The Trangie experiment responses in growth rate, size, maternal ability, reproductive performance, carcase composition, feed requirements and herd productivity. In: The consequences of selection for growth rate in beef cattle. Proc. Aus. Soc. Anim. Prod. 20, 1726. [ Links ]
Retallick, K.M., 2013. Genetic variance and covariance components for feed intake, average daily gain, and postweaning gain and indices to improve feed efficiency in growing beef cattle. MSc, University of Wisconsin-Madison. [ Links ]
Richardson, E.C. & Herd. R.M., 2004. Biological basis for variation in residual feed intake in beef cattle. 2. Synthesis of results following divergent selection. Aust. J. Exp. Agric. 44, 431-440. DOI: 10.1071/EA02221 [ Links ]
Rolfe, K.M., Snelling, W.M., Nielsen,M.K., Freetly, H.C., Ferrell, C.L. & Jenkins, T.G., 2011. Genetic and phenotypic parameter estimates for feed intake and other traits in growing beef cattle, and opportunities for selection. J. Anim. Sci. 89, 3452-3459. DOI: 10.2527/jas.2011-3961 [ Links ]
Sainz, R.D. & Paulino, P.V., 2004. Residual feed intake. UC Davis: Sierra Foothill Research and Extension Center. https://escholarship.org/uc/item/9w93f7ks [ Links ]
Schenkel, F.S., Miller, S.P. & Wilton, J.W., 2004. Genetic parameters and breed differences for feed efficiency, growth, and body composition traits of young beef bulls. Can. J. Anim. Sci. 84, 177-185. DOI: 10.4141/A03-085 [ Links ]
Scholtz, M.M., 2010. Beef breeding in South Africa. 2nd edition. M.M. Scholtz (ed). ARC, Pretoria. ISBN-13, 978-186849-391-3 [ Links ]
Scholtz, M.M., Chadyiwa, M.C., Pyoos-Daniels, G.M., Jordaan, F.J., Hendriks, J., Wepener, M.P., Theunissen, A., Makgahlela, M.L. & Neser, F.W.C., 2021. Climate-smart livestock production in the era of climate change through targeted interventions. Proceedings Climate-smart Livestock Production in Africa Conference, 22-24 June 2021, Nigeria. [ Links ]
Smith, S.N., Davis, M.E. & Loerch, S.C., 2010. Residual feed intake of Angus beef cattle divergently selected for feed conversion ratio. Livest. Sci. 132, 41-47. DOI:10.1016/j.livsci.2010.04.019 [ Links ]
Swalve, H.H., 1993. Estimation of direct and maternal (co)variance components for growth traits in Australian Simmental beef cattle. J. Anim. Breed. Genet. 110, 241-252. DOI: 10.1111/j.1439-0388.1993.tb00736.x [ Links ]
Torres-Vazquez, J.A., Van der Werf, J.H.J. & Clark, S.A., 2018. Genetic and phenotypic associations of feed effciency with growth and carcass traits in Australian Angus cattle. J. Anim. Sci. 96, 4521-4531. DOI: 10.1093/jas/sky325 [ Links ]
Van der Westhuizen, R.R., Van der Westhuizen, J. & Schoeman, S.J., 2004. Genetic relationship between feed efficiency and profitability traits in beef cattle. S. Afr. J. Anim. Sci. 34, 50-52. DOI: 10.4314/sajas.v34i6.3829 [ Links ]
Submitted 31 March 2021
Accepted 1 January 2022
Published 18 January2022
# Corresponding author: GScholtz@arc.agric.za


{kind=link}
{kind=link}
{kind=link}