










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 n.6 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i6.6
Performance and intestinal histology of sheep fed detoxified castor bean meal in sugarcane silage
P.R. Dantas JúniorI; J.S. OliveiraI; N.L. RibeiroII; L.D. RolaIII; E.G. SilvaIII; A.C. OliveiraIV; V.V.S. AlmeidaIV; D.M. Lima JúniorIV; R.R. GuerraI, #
IPrograma de Pós-Graduação em Zootecnia, Centro de Ciências Agrárias, Federal University of Paraíba, Areia, Paraíba, Brazil
IINational Semi-Arid Institute, Campina Grande, Paraíba, Brazil
IIIPrograma de Pós-Graduação em Ciência Animal, Centro de Ciências Agrárias, Federal University of Paraíba, Areia, Paraíba, Brazil
IVCampus de Arapiraca, Federal University of Alagoas, Arapiraca, Alagoas, Brazil
ABSTRACT
This study evaluated histological changes in the digestive organs and kidneys of sheep fed sugarcane silage augmented with detoxified castor bean meal (CBM). Forty Santa Inês sheep (initial weight 20 ± 0.3 kg) were adapted to the experimental regime for 14 days, after which data were collected for 60 days. The treatments consisted of S0: sugarcane silage (SC); S5: SC augmented with 5% CBM; S10: SC augmented with 10% CBM; S15: SC augmented with 15% CBM; and S20: SC augmented with 20% CBM. After 74 days of confinement, the animals were slaughtered, and then fragments of the liver, rumen, intestine, and kidney were collected for histomorphometric analyses. The intake of dry matter and the average weight gain increased linearly with the level of CBM (P <0.05), as did the rumen absorption area, the height of the papillae and the width of the papillae. The thickness of the keratinized portion of the epithelium was not influenced, Thus, no effect was detected of the anti-nutritional factor of CBM on this organ. In the intestine, CBM increased the height of the villi and the number of goblet cells. However, S20 caused some kidney damage and decreased hepatic glycogen stock, but not sufficiently to reduce performance. Thus, augmentation of sugarcane with CBM, up to 20%, is viable in feeding sheep.
Keywords: detoxification, morphometry, Ricinus communis L., rumen histology
Introduction
Food accounts for about 63% of the total cost sheep production in Brazil (Raineri et al., 2015). Almost half of the grains produced in the world are fed to livestock (Sansoucy, 1995). Irregularity in the availability of forage reduces the herd, prompting late slaughter and a supply of meat that is periodically incompatible with the demand of consumers (Pereira et al., 2013). Therefore, alternative feedstuffs are needed to reduce costs and increase sheep production.
The conservation of sugarcane as silage aroused interest because of its logistical and operational benefits. However, in the sugarcane ensiling process, a chemical or bacterial additive should be used that favours fermentation to reduce total losses and improves its nutritional value (Torres et al., 2003; Balieiro Neto et al., 2007) and thus achieves superior results (Freitas et al., 2006). Among these additives, biodiesel wastes such as castor residue (Furtado et al., 2011 ; Gomes et al., 2017) and detoxified castor meal (Oliveira et al., 2016, are promising, since castor meal has approximately 87.3% dry matter, 94.7% organic matter (based on dry matter), 26.9% crude protein, 5% ether extract, 49.8% neutral detergent fibre, 40.1% acid detergent fibre, 62.8% total carbohydrates, 15% non-fibrous carbohydrates, 72.3% total digestible nutrients (percentage of dry matter), and 4.59 Mcal/kg of metabolizable energy (Valadares Filho et al., 2006).
The castor plant (Ricinus communis L.) has high drought tolerance, and produces an oilseed that is economically valuable. The oil has several industrial applications, and the husk, bran, and castor bean meal are processed, yielding 50% oil and 50% by-products (Bomfim et al., 2009). Increased biodiesel production would be economically viable if new applications were found for these by-products. Their use in feed for livestock would be an important asset (Gomes et al., 2017; Oliveira et al., 2016).
The literature indicated that high intake of castor beans by sheep can cause weakness, salivation, profuse watery diarrhoea, dehydration, mydriasis, gnashing of teeth, hypothermia, and decubitus because of the ricin content (Aslani et al., 2007). Severe gastroenteritis, haemorrhage, cardiac necrosis, liver necrosis and acute tubular necrosis in kidneys have also been reported (Santos et al., 2018). Since castor meal has high nutritional value and low commercial value, it could be used as an additive in sugarcane silage and in the feed of small ruminants after detoxification. Therefore, it is necessary to study the form of administration and the possible impacts caused by castor meal as an additive to sugarcane silage on the digestive and renal tissues of sheep Thus, the objective of this research was to evaluate the histological changes of organs of the digestive system and kidneys of sheep subjected to various levels of castor meal in sugarcane silage.
Material and methods
This study was approved by the Animal Use Ethics Committee of the Federal University of Paraíba (UFPB), Brazil, protocol number 02/2018 and was conducted in the Goat and Sheep Sector of the Federal University of Alagoas - UFAL Campus Arapiraca in the city of Arapiraca- AL, (latitude 9° 69' S, longitude 36° 66' W, and average altitude of 305 m). The climate in Arapiraca is tropical, Köppen climate classification type Aw, with an average temperature of 23.7 °C and average rainfall of 752 mm.
Forty Santa Inês crossbred sheep were used, with an average bodyweight of 20 ± 0.324 kg. After a 14-day adaptation period, they were evaluated over 60 days. The animals had been treated for ecto- and endo-parasites, and were confined in individual covered stalls with concrete floors, and furnished with feeders and drinkers.
The diets (Table 1) had a roughage to concentrate ratio of 60 to 40 to provide for a growth rate of 250 g/day (NRC, 2007). The concentrate consisted primarily of soybean meal and corn. The CBM was detoxified with calcium hydroxide (CaOH) diluted at 1 kg per 10 l water and treated with this solution at 60 grams per kg of organic matter (Oliveira et al., 2010). Treatments consisted of S0: unsupplemented sugarcane silage (control), S5: sugarcane silage augmented with 5% castor meal replacing sugarcane; S10: with 10% castor meal replacing sugarcane, S15: with 15% castor meal replacing sugarcane, and S20: with 20% castor meal replacing sugarcane.
The feed was offered as a complete mixture twice a day (07h30 and 16h30) at will to allow 10% leftovers. The quantities of feed offered and the leftovers were recorded daily to estimate intake. The lambs were weighed at the beginning of the experiment and on the day before slaughter to obtain their initial and final weights. These data were used to calculate the average daily gain and dry matter intake (Lima et al., 2018).
Samples of silage and leftovers were collected daily from each animal. Samples of concentrates were collected weekly for each treatment. The daily samples of silage and leftovers were grouped proportionally during each 14-day period to form a composite sample. Subsequently, these samples were dried in a convection oven at 60 °C, ground in a mill with a 1-mm sieve, conditioned in a flask with a lid, and stored until analysed.
The animals were slaughtered in random order after feed had been withheld for 16 hours. At the time of slaughter, the animals were weighed, desensitized with a compressed air pistol, suspended by the hind limbs, and bled by severing the carotid arteries and jugular veins (CONCEA, 2013). While suspended, the animals were skinned manually (Cezar & Sousa, 2007).
Histomorphometric analyses were performed at the Animal Histology Laboratory, Agricultural Science Centre, Federal University of Paraíba. Fragments of the liver and kidney were collected no larger than 0.5 cm3, and of the rumen and small intestine no larger than 1 cm3, fixed in 10% formaldehyde, and packed in identified containers. These fragments were collected from similar locations in all animals.
Histological processing continued with dehydration, clarification and inclusion in paraffin. Dehydration consisted of serial immersion of the tissue sample in solutions of increasing ethyl alcohol concentrations of 70%, 90%, 100% I, and 100% II for one hour each, and then twice for 30 minutes in 50% alcohol and 50% xylol, after which they were embedded in the paraffin.
A microtome was used to cut the embedded tissue samples to a thickness of 5 μm Haematoxylin and eosin stains were used in the histomorphometric and histopathological studies, and periodic acid Schiff (PAS) was used to quantify hepatic glycogen and the duodenal goblet cell index.
The samples were visualized on an Olympus BX53F microscope (Tokyo, Japan), coupled with a digital photo camera (Olympus DP73, Tokyo, Japan). Measurements were made with cellSens Dimension® software using the 40x objective for the liver and kidneys, and 4x objective for the rumen and small intestine.
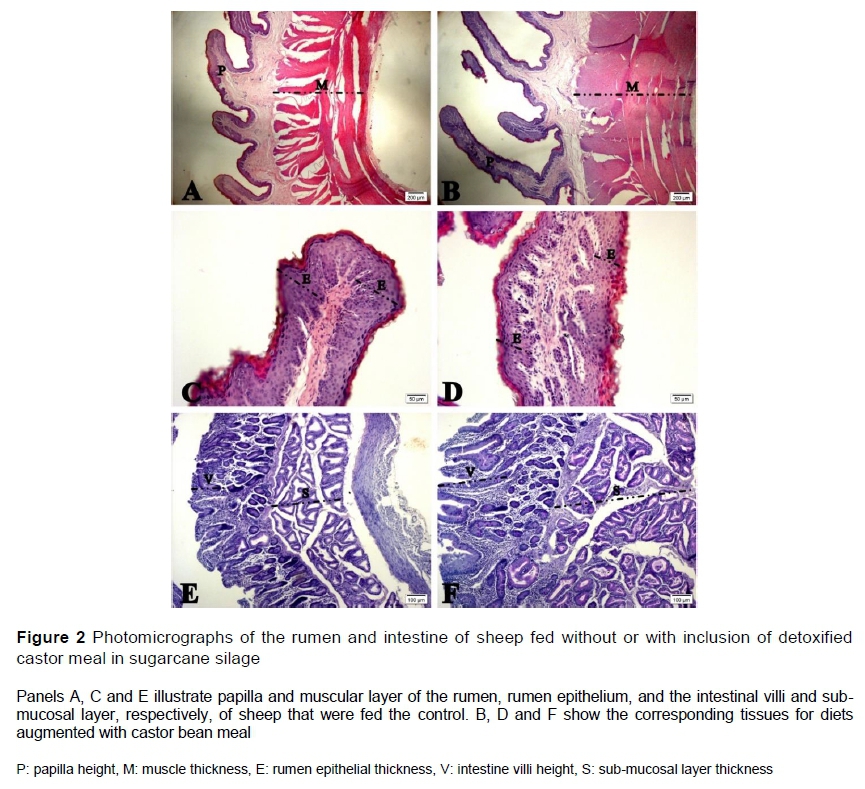
For samples of the rumen and intestine, four photomicrographs per animal were digitized, and three measurements were made of each variable. Thus, there were 96 samples per treatment (8 animals x 4 photomicrograph x 3 measurements). The variables that characterized the rumen consisted of height of the papillae (from the base to the apex), width of the papillae (in the middle region), thickness of the muscular layer, and thickness of the keratinized portion of the epithelium. The variables that characterized the duodenum were villus height (crypt depth and villus to crypt ratio (villus/crypt) (Barboza et al., 2019).
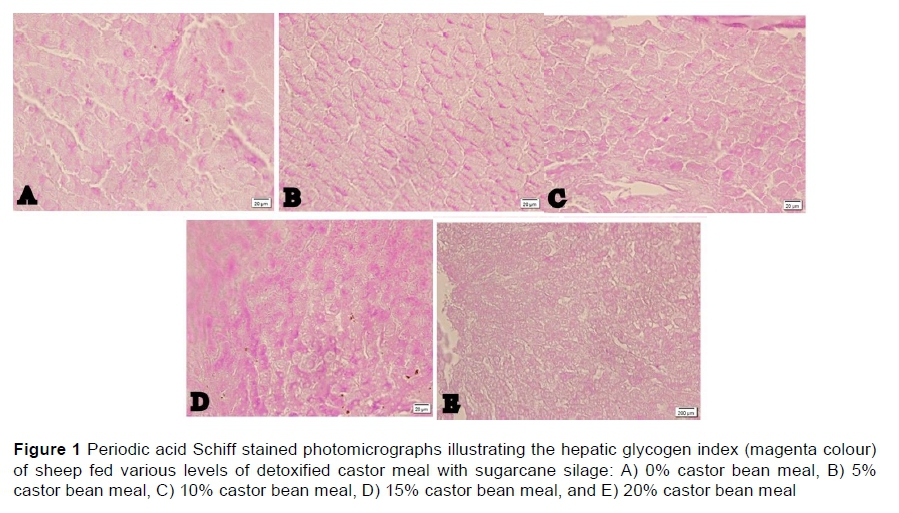
To characterize the liver, five photomicrographs per animal were digitized, totalling 40 observations per treatment. To quantify hepatic glycogen, the same observer verified the degree of positivity to PAS staining (proportional to the amount of hepatic glycogen stock) in each of the photomicrographs, giving a score ranging from 0 to 3, with 3 being the highest degree of deposition of glycogen (Ishak, 1995).
Six photomicrographs per animal were digitized to characterize the kidney, yielding 48 observations per treatment. Each of the photomicrographs was examined for histopathological changes in the renal corpuscle, proximal twisted tubules, Henle loop, and distal twisted tubules to ascertain the possibility of kidney damage because of anti-nutritional factors in the CSM.
Five photomicrographs per animal were also used to determine the goblet cells index of the duodenum. For each animal, the number of goblet cells in a 2000 μπι length of intestinal epithelium was counted, using four segments of 500 μm (Barboza et al., 2019). Thus, the number of observations per treatment was 32.
The experiment followed a completely randomized design with five levels of dietary castor meal (0%, 5%, 10%, 15%, and 20%) and eight repetitions. Data were analysed with analysis of variance and regression using PROC GLM (SAS Institute, Inc., Cary, North Carolina, USA). Effects were declared significant at the 5% level of probability. The linear model was:
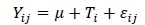
where: 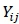 is the dependent variable observed on the jth animal,
is the dependent variable observed on the jth animal,
is the overall mean,
is the effect of treatment ;i, and
is the experimental error.
Results and Discussion
Dry matter intake increased linearly as CBM was added (Table 2), whereas weight gain and the glycogen stock index displayed curvilinear responses. Intake and weight gain were 0.67 kg and 8.9 kg in the control and 1.10 kg and 13.48 kg in S20 (Table 2). The highest values of glycogen stock were observed in S5 and S15. The lowest glycogen stock was observed in S20. S0 and S10 produced values of the glycogen stock index that were intermediate and not different from either extreme.
Figure 1 illustrates the effects of the treatments on the liver glycogen index for sheep fed various levels of CBM in sugarcane silage. A lower index was noted for S20 compared with the other treatments in which the sheep were fed CBM.
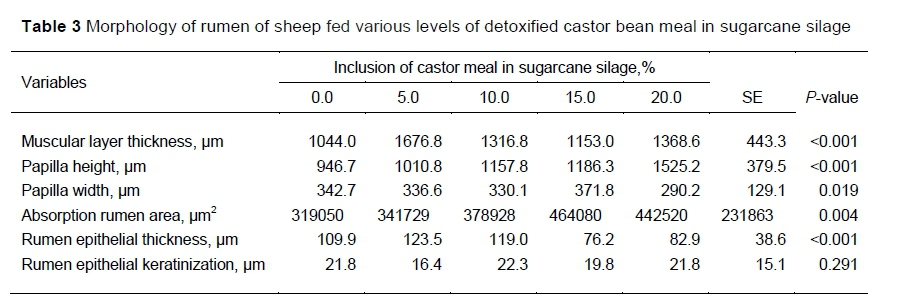
The muscular layer was significantly thicker with CBM at all levels except S15. The height of the rumen papilla was significantly greater when CBM was provided. However, the width of the papilla was unaffected by the treatments. The absorptive surface area of the rumen was increased and the thickness of the rumen epithelium was reduced for S15 and S20. However, the thickness of the keratinized portion of the epithelium was not affected significantly by the treatments (P >0.05). Table 3 presents the data that characterized the rumen morphology.
Figure 2 illustrates the effects of augmenting sugarcane silage with CBM. The comparison of panels A and B shows the greater height of ruminal papilla and muscular layer thickness in animals fed CBM (panel B) compared with the control (panel A). Panels C and D illustrate the change in rumen epithelium from incorporating CBM. Panels E and F show the changes in the intestinal villi and sub-mucosal layer.
Treatment effects were significant on villus height, crypt depth, villus/crypt index, mucosal layer thickness, sub-mucosal layer thickness, and the number of goblet cells (Table 4). With the exception of S15, adding CBM to the sugar can silage increased the villus height. Relative to S0, crypt depth was consistently increased by the treatments. Thus, the ratio of villus height to crypt depth was similar to the control for all treatments except S15. The mucosal layer thickness was increased significantly by S5, S10 and S20 relative to S0. However, S15 and S0 produced similar thicknesses of the mucosal layer. The sub-mucosal thickness was increased by all treatments relative to S0, except for S10. S5 and S10 did not increase the number of goblet cells compared with S0. However, S15 and S20 produced higher numbers of goblet cells than was observed with S0.
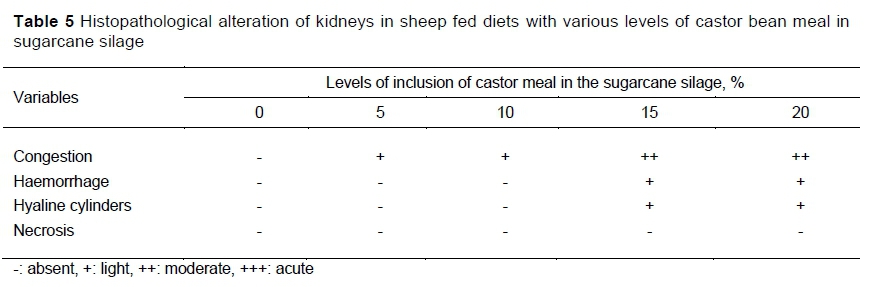
Data in Table 5 summarize the histopathological observations. No renal alterations were found in S0. In S5 slight congestion was found in the cortical and medullary region of the kidneys, and slight congestion in the cortical region in S10. In S15, it was possible to visualize a focal area of haemorrhage and moderate congestion in the medullary region. Finally, S20 provoked a focal area of haemorrhage with moderate congestion in the cortical and medullary region. In both S15 and S20, a slight formation of hyaline cylinders was observed in the glomeruli.
The use of CBM increased the intake of dry matter and the weight gain linearly. Such gains could be explained by the increase in the papillae and the area of ruminal absorption, and in the intestinal villus and goblet cells, which generate greater intestinal health. However, hepatic glycogen stock was affected deleteriously by S20. Ricin in castor bean (Aslani et al., 2007; Santos et al., 2018) was believed to affect the glycogen stock negatively and decrease the conversion of propionate to glycogen. Thus, ricin was not completely removed from CBM by the detoxification process. Glycogen is converted to glucose in the liver, which provides energy or most of the metabolism and production (Brown, 2018). Although S20 had the lowest hepatic glycogen reserve, it did not influence weight gain negatively. On the contrary, sheep in S20 had the greatest weight gain, probably because of the short time for which the animals were confined.
The use of CBM to augment sugarcane silage did not cause pathological morphometric changes to the rumen and intestine, and resulted in better animal performance. Its inclusion may have allowed greater nutrient absorption from the rumen because the increased height of the papillae (Barboza et al., 2019; Silva et al., 2019). The increased size of ruminal papillae could be because CBM increased the amount of concentrate in the diet. It is known that concentrate promotes an increase in volatile fatty acids (VFA), especially when the concentrate has a high carbohydrate content, as in CBM (Caetano et al., 2016; Wang et al., 2017). These VFA have a direct relationship with epithelial growth. Propionate and butyrate are responsible mainly for cell proliferation and consequently an increase in the papillae (Baldwin et al., 2004; Barboza et al., 2019). Thus, the potential for increased production and absorption of VFA provided more energy for ruminants, enabling them to satisfy their requirements (Goularte et al., 2011; Oliveira et al., 2013) and perform better (Monção et al., 2013).
Although rumen lesions were not found in this study, Bianchi et al. (2018) stated that sheep fed castor bean in the trough had discrete multifocal hydropic degeneration in the mucosa of the rumen and other fermentative organs, associated with a mild inflammatory infiltrate composed of lymphocytes and plasma cells in the sub-mucosa. Therefore, the detoxification process used in this study may have reduced the ricin content sufficiently to not cause injury, in this organ at least.
It was expected that the thickness of the muscular layer of the rumen would decrease as the level of the CBM was increased because the roughage content of the diet was diminished. The more bulk in the diet, the greater the need for peristaltic movements for maceration and rumination, which lead to greater hyperplasia and hypertrophy of the smooth muscle fibres of the rumen (Silva et al., 2019). This result can be rationalized by the rise in dry matter intake, which increased linearly with the inclusion of CBM, which provided a greater volume of ingestion, and consequently more peristaltic movements, despite a smaller amount of roughage.
The ruminal epithelium became thinner with the inclusion of 15% and 20% CBM, which can be explained by the reduced abrasion imposed by the CBM in relation to the sugarcane silage. This characteristic can generate energy savings by reducing the turnover rate of these cells (Brandi & Furtado, 2009). The keratinized portion did not change, which may signal that CBM did not present ricin or that the amount was low enough not to cause hypertrophy of that portion of the epithelium (Barboza et al., 2019; Lima et al., 2018).
The use of CBM increased the height of the intestinal villi, except in S15, which had wide variations among animals. In general, the inclusion of CBM in the diet increased the capacity for nutrient absorption (Wang et al., 2009), which was manifest in the linear increase in weight. Villus morphology changes when more nutrients are available or there is a greater demand for energy (Montanholi et al., 2013). Increased crypt depth was also observed, which is associated with a greater need for cell renewal, because of an increase in villus size, or injuries, such as those caused by pathogens, mechanical abrasion, toxins, and anti-nutritional agents (Silva et al., 2010). However, the expected decrease in the ratio of villus height to crypt depth was not observed except in S15 because of a reduction in abrasion relative to S0. This could cause greater cellular desquamation and proliferation in the intestinal crypt and greater depth. However, in this study, CBM inclusion caused an increase in intake which may have enhances rate of passage through the intestine and greater cell desquamation.
The villus to crypt ratio is widely used to check intestinal health, and the higher the ratio, the better the health. That is, a large villus and a shallow crypt were expected. If the crypt was deep, this meant excessive need for cell proliferation owing to aggression to the epithelium, which also leads to greater energy expenditure (Aptekmann et al., 2001; Barboza et al., 2019; Wang et al., 2009). In the present study, CBM led to an increase in intestinal villi, which improved the absorption of nutrients and increased weight gain, but without improving intestinal health.
Goblet cells are also used to measure intestinal health, and the greater the number of these cells, the better the animal's intestine health. The inclusion of CBM at 15% increased these cells, which produce mucin, which has several functions, including aid to intestinal transit (peristalsis), mechanical protection of the intestinal epithelium, protection against infectious agents to the intestinal mucosa, and composition of intestinal glycocalyx (Rocha et al., 2016). In this study, it was inferred that the increase in mucin would protect the intestinal surface from ricin, which causes watery diarrhoea (Aslani et al., 2007) and severe gastroenteritis (Santos et al., 2018). Another reason for the increase in goblet cells is the enhanced intake of dry matter from CBM. In other words, the increases in goblet cells and in mucin improved intestinal health.
The inclusion of CBM in the diet led to mild to moderate renal congestion. In addition, mild haemorrhage and hyaline cylinders were observed in the kidneys of animals in S15. The presence of hyaline cylinders in the glomeruli, with eosinophilic material in the urinary spaces, could suggest an ultra-filtered protein. Ricin can cause necrotic tubular degeneration, glomerulonephritis, and renal congestion (Botha & Penrith, 2009). However, such lesions were not found in this study. Ricin is also cited as promoting haemorrhage and multi-focal necrosis in lymphoid organs (Bianchi et al., 2018), degeneration of hepatocytes .and dilation of hepatic sinusoids (Botha & Penrith, 2009), congestion and haemorrhage in the gastrointestinal tract (Santos & Alessi, 2017), and epithelial cells of intestinal crypts that are focal necrotic and diffusely haemorrhagic sub-mucosal (Roels et al., 2010; Hong et al., 2011). Although the lesions were not found in the present study at the level of the liver and intestine, it was possible to verify the decrease in hepatic glycogen stock in animals that were fed S20.
The lesions in the kidney and the decrease in hepatic glycogen in S20 demonstrate that at least part of the ricin was not eliminated in the detoxification process. However, the ricin content of the diet was not enough to alter the performance of animals negatively. Animals fed with CBM showed greater weight gain and greater intake of dry matter.
Conclusions
Castor bean meal is a cheap by-product of biodiesel production. When it is detoxified and used to augment sugarcane silage up to 20% based on dry matter, CBM may promote ruminal and intestinal alterations that are favourable to nutrient absorption and increased weight gain when fed to sheep.
Acknowledgements
To CAPES for the scholarship granted to the first author.
Authors' Contributions
PRDJ, VVSA, DMLJ and RRG collected the data for this study, conducted the statistical analyses, collaborated in interpretation of the results, and wrote the initial draft of this manuscript; DMLJ, JSO, and RR. developed the original hypotheses, designed the experiments, interpreted the results, and finalized the manuscript. All authors read and approved the finalized manuscript.
Conflict of Interest Declaration
The authors certify that they have no affiliations with any organization or entity with financial or non-financial interests in the subject matter or materials discussed in this manuscript.
References
Albretsen, J.C., Gwaltney-Brant, S.M. & Kahn, S.A., 2000. Evaluation of castor bean toxicosis in dogs: 98 cases. J. Am. Anim. Hosp. Assoc. 36, 229-233. DOI: 10.5326/15473317-36-3-229 [ Links ]
Aptekmann, K.P., BaraldiArton, S.M., Stefanini, M.A. & Orsi, M.A., 2001. Morphometric analysis of the intestine of domestic quails (Coturnix coturnix japonica) treated with different levels of dietary calcium. Anat. Histol. Embryol. 30, 277-280. DOI: 10.1046/j.1439-0264.2001.00331.x [ Links ]
Aslani, M.R., Maleki, M., Mohri, M., Sharifi, K., Najjar-Nezhad, V. & Afshari, E., 2007. Castor bean (Ricinus communis) toxicosis in a sheep flock. Toxicon 49, 400-406. DOI: 10.1016/j.toxicon.2006.10.010 [ Links ]
Baldwin, R.L., Klotz, J.L. & Heitmann, R.N., 2004. Rumen development, intestinal growth and hepatic metabolism in the pre- and postweaning ruminant. J. Dairy Sci. 87, e55-e65. https://doi.org/10.3168/jds.S0022-0302(04)70061-2 [ Links ]
Balieiro Neto, G., Siqueira, G.R., Reis, R.A., Nogueira, J.R., Roth, M.T.P. & Roth, A.P.T., 2007. Calcium oxide as additive on the sugarcane ensilage. Vet. Bras. Zootec. 36, 1231-1239. https://www.rbz.org.br/article/calcium-oxide-as-additive-on-the-sugarcane-ensilage/ [ Links ]
Barboza, S.C.R., Oliveira, J.S., Souza, M.T.C., Lima Júnior, D.M., Lima, H.B. &Guerra, R.R., 2019. Ovines submitted to diets containing cassava foliage hay and spineless cactus forage: Histological changes in the digestive and renal systems. Trop. Anim. Health Prod. 51, 1689-1697. DOI: 10.1007/s11250-019-01863-9 [ Links ]
Bianchi, M.V., Vargas, T.P., Leite Filho, R.V., Guimarães, L.L.B., Heck, L.C., Pavarini, S.P. & Driemeier, D., 2018.Intoxicação espontânea por Ricinus communis em ovinos. Acta Sci. Vet. 46, 294. https://www.lume.ufrgs.br/handle/10183/180839 [ Links ]
Bomfim, M.A.D., Silva, M.M.C. & Santos, S.F., 2009. Potencialidades da utilização de subprodutos da indústria de biodiesel na alimentação de caprinos e ovinos. Tecnologia e Ciência Agropecuária 3, 15-26. [ Links ]
Brandi, R.A. & Furtado, C.E., 2009. Importância nutricional e metabólica da fibra na dieta de equinos. Vet. Bras. Zootec. 38, 246-258. DOI: 10.1590/S1516-35982009001300025 (in Portuguese, English abstract). [ Links ]
Brown, T.A., 2018. Bioquímica. 1st ed. Guanabara Koogan, Rio de Janeiro, Brazil. [ Links ]
Botha, C.J. & Penrith, M.L., 2009. Potential plant poisonings in dogs and cats in southern Africa: A review article. J. S. Afr. Vet. Assoc. 80,63-74. DOI: 10.4102/jsava.v80i2.173 [ Links ]
Caetano Junior, M.B., Caetano, G.A.O. & Oliveira, M.D., 2016. A influência da dieta no desenvolvimento ruminal de bezerros. Nutritime Rev. Eletrôn. 13, 4902-4918 (in Portugese, English abstract). [ Links ]
Cezar, M.F. & Sousa, W.H., 2007. Carcaças ovinas e caprinas: obtenção, avaliação e classificação. 1st ed. Editora Agropecuária Tropical, Uberaba, Brazil. [ Links ]
Freitas, A.W.P., Pereira, J.C., Rocha, F.C., Costa, M.G., Leonel, F.P. & Ribeiro, M.D., 2006. Evaluation of the nutritional quality of sugarcane silage treated with microbial additives and soybean crop residue. Ver. Bras. Zootec. 35, 3847. https://publons.com/publon/7915799/ [ Links ]
Furtado, C.E., Brandi, R.A. & Ribeiro, L.B., 2011 .Utilização de coprodutos e demais alimentos alternativos para dietas de equinos no Brasil. Ver. Bras. Zootec. 40, 232-241 (in Portuguese, English abstract). [ Links ]
Gomes, F.H.T., Cândido, M.J.D., Carneiro, M.S.S., Furtado, R.N. & Pereira, E.S., 2017. Intake, behavior and performance in sheep fed diets containing castor cake. Revista Ciência Agronômica 48, 182-190. DOI: 10.5935/1806-6690.20170021 [ Links ]
Goularte, S.R., Ítavo, L.C.V., Santos, G.T., Ítavo, C.C.B.F., Oliveira, L.C.S., Favaro, S.P., Dias, A.M., Torres Júnior, R.A.A. & Bittar, C.M.M., 2011. Ácidos graxos voláteis no rúmen de vacas alimentadas com diferentes teores de concentrado na dieta. Arq. Bras. Med. Vet. Zootec. 63, 1479-1486. DOI: 10.1590/S0102-09352011000600027 (in Portuguese, English abstract). [ Links ]
Hong, I.H., Kwon, T.E., Lee, S.K., Park, J.K., Ki, M.R., Park, S.I. & Jeong, K.S., 2011. Fetal death of dogs after the ingestion of a soil conditioner. Exp. Toxicol. Pathol. 63, 113-117. DOI:10.1016/j.etp.2009.10.004 [ Links ]
Ishak, K., Baptista, A., Bianchi, L., Callea, F., Groote, J., Gudat, F., Denk, H., Desmet, V., Korb, G. & MacSween, R.N., 1995. Histological grading and staging of chronic hepatitis. J. Hepat. 22, 696-699. DOI: 10.1016/0168-8278(95)80226-6 [ Links ]
Lima, T.J., Costa, R.G., Medeiros, G.R., Medeiros, A.N., Ribeiro, N.L., Oliveira, J.S., Guerra, R.R. & Carvalho, F.F.R., 2018. Ruminal and morphometric parameters of the rumen and intestines of sheep fed with increasing levels of spineless cactus (Napolea cochenillifera Salm Dyck). Trop. Anim. Health Prod. 51, 363-368. DOI: 10.1007/s11250-018-1697-1 [ Links ]
Monção, F.P., Oliveira, E.R.R., Moura, L.V., Tonissi, R.H.H. & Góes, B., 2013. Development of microbiotaruminal calf -literature review. Revista Unimontes Científica 15. [ Links ]
Montanholi, Y., Fontoura, A., Swanson, K., Coomber, B., Yamashiro, S. & Miller, S., 2013. Small intestine histomorphometry of beef cattle with divergent feed efficiency. Acta Veterinaria Scandinavica 55(1), 9. https://doi.org/10.1186/1751-0147-55-9 [ Links ]
National Research Council (NRC), 2007. Nutrient requirements of small ruminants: Sheep, goats, cervids, and New World Camelids. 1st ed. National Academy Press, Washington, USA. [ Links ]
Oliveira, A.S., Campos, J.M.S., Oliveira, M.R.C., Brito, A.F., Valadares Filho, S.C., Detmann, E., Valadares, R.F.D., Souza, S.M. & Machado, O.L.T., 2010. Nutrient digestibility, nitrogen metabolism and hepatic function of sheep fed diets containing solvent or expeller castor seed meal treated with calcium hydroxide. Anim. Feed Sci. Techn. 158, 15-28. DOI: 10.1016/j.anifeedsci.2010.02.009 [ Links ]
Oliveira, V.S., Santana Neto, J.A. & Valença, R.L., 2013. Características químicas e fisiológicas da fermentação ruminal de bovinos em pastejo - Revisão de Literatura. Revista Científica Eletrônica de Medicina Veterinária, 20. [ Links ]
Oliveira, H.C., Garcia, R., Almeida, V.V.S., Oliveira, A.C., Pires, A.J.V., Nascimento Filho, C.S., Veloso, C.M., Silva, R.R. & Oliveira, U.L.C., 2016. Comportamento ingestivo de cordeiros alimentados com farelo de mamona. Semin. Ciênc. Agrár. 37, 1451-1460. [ Links ]
Pereira, L.G.R., Aragão, A.L.S., Santos, R.D., Azevedo, J.A.G., Neves, A.L.A., Ferreira, A.L. & Chizzotti, M.L., 2013. Productive performance of confined sheep fed mango meal. Arq. Bras. Med. Vet. Zootec. 65, 675-680. DOI: 10.1590/S0102-09352013000300009 [ Links ]
Raineri, C., Stivari, T.S. & Gameiro, A.H., 2015. Lamb production costs: Analyses of composition and elasticities analysis of lamb production costs. Asian-Australasian J. Anim. Sci. 28(8), 1209-1215. https://doi.org/10.5713/ajas.14.0585 [ Links ]
Rocha, P.M.C., Barros, M.E.G. & Evêncio-Neto, J., 2016. Morphometric analysis of the intestinal wall and the dynamic of mucins secreted in the jejunum of broilers supplemented with Bacillus subtilis strain C3102. Pesqui. Vet. Bras. 36, 312-316. DOI: 10.1590/S0100-736X2016000400010 [ Links ]
Roels, S., Coopman, V., Vanhaelen, P. &Cordonnier, J. 2010. Lethal ricin intoxication in two adult dogs: Toxicologic and histopathologic findings. J. Vet. Diagn. Invest. 22, 466-468. DOI: 10.1177/104063871002200325 [ Links ]
Sansoucy, R., 1995. Livestock - a driving force for food security and sustainable development. World Anim. Rev. 84/85,.5-17. [ Links ]
Santos, C.B., Araújo, M.J., Bezerra, L.R., Marques, C.A.T., Torreão, J.N.C., Freitas, N.E., Oliveira Neto, C.B. & Morais, J.S., 2018. Hematological and biochemical parameters of lactating goats fed diets containing crude glycerin from biodiesel production from waste frying oil. Arq. Bras. Med. Vet. Zootec. 70, 1867-1876. DOI: 10.6084/m9.figshare.7482671 .v1 [ Links ]
Santos, P.A., Ludke, M.C.M., Ludke, J.V., Santos, M.J.B., Melo, A.G.S., Oliveira, A.C. & Cavalcanti, A.S.A., 2013. Castor meal in feeding of non-ruminants. Revista Eletrônica Nutritime 10, 2814-2827. [ Links ]
Santos, R.L. & Alessi, A.C., 2017. Patologia Veterinária. (2nd ed.), Roca, Rio de Janeiro, Brazil. [ Links ]
Silva, L.C.R., Furuya, W.M., Natali, M.R.M., Schamber, C.R., Santos, L.D. & Vidal, L.V.O., 2010. Productive performance and intestinal morphology of Nile tilapia juvenile fed diets with L-glutamine and L-glutamate. Rev. Bras. Zootec. 39, 1175-1179. .org/10.1590/S1516-35982010000600002 [ Links ]
Silva, K.B., Oliveira, J.S., Santos, E.M., Cartaxo, F.Q., Guerra, R.R., Souza, A.F.N., Muniz, A.C.S.& Cruz, G.F.L., 2019. Ruminal and histological characteristics and nitrogen balance in lamb fed diets containing cactus as the only roughage. Trop. Anim. Health Prod. 52, 637-645. DOI: 10.1007/s11250-019-02051-5 [ Links ]
Torres, L.B., Ferreira, M.A., Véras, A.S.C., Melo, A.A.S. & Andrade, D.K.B., 2003. Sugarcane bagasse and urea as replacement of soybean meal in the growing dairy cattle diets. Ver. Bras. Zootec. 32, 760-767. DOI: 10.1590/S1516-35982003000300029 [ Links ]
Valadares Filho, S.C., Magalhães, K.A. & Rocha Junior, V.R., 2006. Tabelas brasileiras de composição de alimentos para bovinos. (2nd ed.), Suprema Gráfica Ltda, Viçosa, BRA. [ Links ]
Wang, B., Wang, D., Wu, X., Xai, J., Liu, M., Huang, X., Wu, J., Liu, J. & Guan, L., 2017. Effects of dietary physical or nutritional factors on morphology of rumen papillae and transcriptome changes in lactating dairy cows based on three different forage-based diets. BMC Genomics, 18, 353. DOI: 10.1186/s12864-017-3726-2 [ Links ]
Wang, Y.H., Xu, M., Wang, F.N., Yu, Z.P., Yao, J.H., Zan, L.S & Yang, F.X., 2009. Effect of dietary starch on rumen and small intestine morphology and digesta pH in goats. Livest. Sci. 122, 48-52. doiDOI: 10.1016/j.livsci.2008.07.024 [ Links ]
Submitted 28 December 2020
Accepted 4 November 2021
Published 26 December 2021
# Corresponding author: ricardo@cca.ufpb.br


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}