










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 n.6 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i6.2
ARTICLES
Response of sheep fed urea-treated corncob and supplemented with cassava leaf meal
D. Yulistiani#; W. Puastuti; E. Wina
Indonesian Research Institute for Animal Production, Jl. Veteran III, Ciawi, Bogor.16720. P.O Box 221. West Java, Indonesia
ABSTRACT
The objective of this study was to evaluate the effects of feeding untreated or urea-treated ground corncob and supplementing with cassava leaf meal (CLM) in a total mixed ration on growth, feed intake, nutrient digestibility, nitrogen (N) utilization and rumen fermentation of sheep. Five diet treatments with five replications consisted of untreated corncob + concentrate (CC); urea-treated corncob + concentrate (UCC); and CC and UCC supplemented with CLM and designated as CC+CLM and UCC+CLM, respectively. A diet formulated with a mixture of elephant grass and concentrate was used as control (EG). There were no significant differences in dry matter intake (DMI) between treatments. The average DMI was 4.17% bodyweight. Average daily gain (ADG) of sheep fed the treatment diets was between 146.3 and 176.2 g/h/day, and was higher than EG (89.1 g/head/day). Thus, the treatments improved feed conversion ratio (FCR). Nitrogen retention was lowest for EG and highest for UCC and UCC+CLM. The addition of CLM had no effect on growth, feed intake, nutrient digestibility, nitrogen utilization and rumen fermentation characteristics. It was concluded that corncob could be used as a replacement for elephant grass in postweaning diets for sheep.
Keywords: digestibility, growth, nitrogen utilization, total mixed ration
Introduction
From 2016 to 2018, Indonesia was among the top ten maize producing countries in the world (OECD/FAO, 2019). Maize is one of the strategic food crop commodities that were targeted by the Indonesian government for domestic self-sufficiency to support the food and feed industry. The rise in maize grain production would result in a large increase in by-products, especially corncobs, which can be used as a fibre source in ruminant feed, particularly in a dry season when the availability of grass is limited. However, corncob is generally characterized as having low crude protein (CP) and digestibility, whereas its fibre content is high (Khan et al., 2004; Yulistiani et al., 2012). Urea treatment was an effective method of increasing CP content and in vitro digestibility of corncob (Yulistiani et al., 2012). In a total mixed ration, ground corncob could replace rice straw as a fibre source for crossbred Holstein dairy cows without affecting milk production (Wachirapacorn et al., 2016). Previous studies have evaluated mixtures of corncob and cowpea husk as a basal diet of West African dwarf sheep (Ososanya et al., 2013), and urea-treated corncobs have been used to feed Arsi-Bale sheep (Negewo et al., 2018).
Improving feed intake, feed digestibility, and productivity of ruminants fed low-quality forage can be achieved through supplementing with a natural protein source or non-protein nitrogen (NPN) (McGuire et al., 2013). However, natural protein supplementation is superior to NPN because of its amino acid and metabolizable protein content. Natural protein, particularly rumen degradable protein, provides amino acids that are catabolized into branched-chain volatile fatty acids, which serve as food for fibre-degrading bacteria in the rumen (Arroquy et al., 2004). Increased activity by cellulolytic bacteria in response to protein supplementation enhanced fibre digestibility and provided high-quality rumen microbial protein for the host animal (May et al., 2003). A sufficient supply of RDP optimized rumen microbial growth and increased the inflow of metabolizable protein to the intestine (Naves et al., 2015). Cassava leaves contained 19 - 25% CP, with almost 85% as true protein (Ravindran, 1993; Oni et al., 2010; Jaswandi & Jhon, 2019) and had a higher concentration of amino acids compared with alfalfa hay (Wanapat et al., 2000). Thus cassava leaves could be used as a natural protein supplement to increase microbial protein synthesis and fibre digestibility in the rumen. Various forms of cassava leaf - fresh cassava foliage, cassava leaf hay, CLM, and cassava top silage - have been used as protein sources to supplement low-quality forage (Hue et al., 2008; Ampapon et al., 2016; Odusanya et al., 2016; Wanapat et al., 2018). Fresh cassava foliage supplementation was able to replace concentrate in lambs fed urea-treated rice straw (Hue et al., 2008). Cassava hay was used as a concentrate for dairy cattle fed lime-treated rice straw supplemented with urea (Lunsin et al., 2012). West African dwarf sheep were fed cassava hay as a supplement to fresh chopped Megathyrsus maximus (Jacq.) (Odusonya et al., 2016). Likewise, cassava hay was used with urea-treated rice straw as feed for lactating dairy cattle (Wanapat et al., 2000). Cassava top silage could be used as a supplement to ammoniated rice straw for dairy cattle (Wanapat et al., 2018) and to low-quality grass fed to goats (Phengvichith & Ledin, 2007). Cassava hay meal was also used as rumen enhancer in a rice straw basal diet for dairy steers (Phesatcha et al., 2016).
Previous studies focused on cassava leaf supplementation of rice straw or grass basal diets, and few reported the effects of supplementation on diets containing corncob. It was expected that cassava leaf would increase rumen fermentation, which would enhance sheep performance. Therefore, the objective of the study was to assess growth performance, feed intake, feed efficiency, nutrient digestibility, nitrogen utilization, and rumen fermentation of sheep fed diets that contained untreated or urea-treated ground corncob with and without CLM.
Materials and Methods
About 500 kg of dry corncob was obtained from maize fields and ground to pass through a 5-mm sieve. The urea-corncob was sprayed with 3% urea solution (1 L/kg dry matter (DM) straw), mixed thoroughly, and put in black plastic bags (5 kg cob/bag), which were pressed carefully to remove the entrapped air, sealed tightly, and stored for a minimum of three weeks. After incubation, the excess ammonia in the corncob was evaporated by spreading it evenly on a concrete floor for a day before offering it to the sheep.
Cassava leaf meal was prepared by harvesting cassava foliage at a cassava farm in Bogor District after the cassava tubers had been gathered at about 10 months old. The foliage consisted of the young stem, leaf, and the tip of the stem from the top down with a length of approximately 50 cm and was chopped to about 5 cm pieces, followed by sun-drying for three days. After drying, the DM content of the foliage was approximately 90%. It was ground to pass through a 2-mm sieve and kept for use in the experiment.
The animal experimentation was conducted after a review by the Institutional Animal Care and Use Committee of the Ministry of Agriculture Indonesia based on the Guide for the Care and Use of Laboratory Animals (NRC 2011). The growth trial was carried out at the Research Station of Indonesian Research Institute for Animal Production (IRIAP), Bogor, Indonesia. The sheep that were used in the study were of the Compass Agrinak breed, which is a composite of 50% local Sumatera, 25% St Croix and 25% Barbados Blackbelly. In total, 25 weaned lambs were used, with an average BW of 15.6 ± 3.28 kg. Each sheep was kept in an individual pen during the 12 weeks of the experiment. The sheep were stratified by initial bodyweight and assigned to one of the five diet treatments.
The experiment was conducted in a completely randomized block design with five replications in each treatment. The treatments consisted of experimental diets that were offered as total mixed rations containing 40% untreated corncob or 40% urea-treated corncob and 60% concentrate (Table 1). In the diets that were supplemented with CLM, the corncob was reduced to 38% and the concentrate to 57%. The control diet was formulated with 40% elephant grass and 60% concentrate. The concentrate mixture contained 12% coconut meal, 30% rice bran, 20% soybean meal, 24% ground maize grain, 20% soybean meal molasses, 1% urea, 1.5% salt, and 1.5% mineral supplement.
Feed consumption were measured daily by subtracting the amount of orts from the amount of feed offered. Then orts from the previous day were weighed before feed was offered in the morning. Data on ADG during the 12 weeks of the experiment was calculated from the weights of the sheep obtained weekly in the morning before feeding. Feed conversion ratio was calculated by dividing daily DMI by ADG. At the end of the growth trial, the sheep were moved to metabolic crates for a digestibility trial.
The digestibility trial consisted of a 14-day period for adaptation to the metabolic crates, followed by seven days of sample collection with rumen fluid collected on a subsequent day. Feed intake, orts, and the amounts of excreted faeces and urine of individual sheep were measured daily during the collection period. A sample consisting of 10% of the daily total faecal production was oven-dried at 60 °C for 48 hours prior to analysis of chemical composition. At the end of the seven-day collection period, the faeces samples from each sheep were pooled to provide a single sample for each animal and a 10% sub-sample was taken, ground to pass through 1 mm sieve, and kept frozen until it was analysed. Similar to the faeces collection, the total daily urine excretion of each sheep was collected every morning in a bucket containing 100 mL of 10% sulfuric acid to maintain pH below 3 to inhibit microbial activity and N losses. The volume of urine was recorded daily, mixed thoroughly, and a representative sample of 10% was taken and kept in the freezer. After the collection period, the daily samples were pooled for each sheep and 10% sub-samples were taken to measure urinary N. On the last day of the digestibility trial, rumen fluid was collected from each sheep four hours after morning feeding with a stomach tube. After sampling, the ruminal fluid pH was measured immediately using a portable pH temperature meter (LaMotte pH 5 PLUS, Maryland, USA). One drop of concentrated sulfuric acid was added to this ruminal fluid (to halt microbial activity) and then centrifuged at 3000 g for 10 min. After centrifugation, approximately 10 mL of the supernatant was removed, kept in an airtight container, and stored at -20 °C until analysed for ammonia (NH3-N) content and VFAs.
Chemical analysis for feeds, residues and faeces included DM, organic matter (OM), and CP contents according to the procedures of AOAC (2012). The methods of Van Soest et al. (1991) were applied to determine neutral detergent fibre (NDF) and acid detergent fibre (ADF). Rumen NH3-N concentrations were analysed using the micro-diffusion technique of Conway (1962). Total and individual VFA concentrations were assessed with gas chromatography (Chrompack CP-9002, Chrompack, Inc., Raritan, New Jersey, USA). Microbial population referred to the number of protozoa, which was determined with a haemocytometer and the bacterial population was recorded by the roll tube method (Ogimoto & Imai, 1981).
An analysis of variance was implemented for the randomized complete block design using SAS version 9.1 (SAS Inc., Cary, North Carolina, USA). Duncan's multiple range tests were applied to differentiate the treatment means at a probability of 5%.
Results and Discussion
In this study, the CP content of corncob was 56 g/kg (Table 1), which was higher than results reported by previous researchers, which varied from 23 to 44 g/kg (Negewo et al., 2018; Ososanya et al., 2013; Wachirapacorn et al., 2016; Yulistiani et al., 2012), but may have been affected by different maize cultivars. The urea treatment of the corncob increased its CP content by 20% (Table 1), which is much lower than in previous studies, which varied from 73% to 190% (Oji et al., 2007; Kayastha et al., 2012; Negewo et al., 2018). This increase resulted from the rates at which urea was applied. Oji et al. (2007) used 5.3% urea to increase the CP content of corncob from 26 to 45 g/kg. Negewo et al. (2018) used 4% urea to increase its CP content from 44 to 93 g/kg. Other factors that affected the CP content included ambient temperature, moisture content, treatment period, sealing, and quality of the forage (Chenost & Kayouli 1997; Oji et al., 2007).
The CP content of cassava leaf in the current study was 231 g/kg (Table 1), which falls within the range of 160 to 250 g/kg reported by other researchers (Ravindran, 1993; Hue et al., 2008; Ampapon et al., 2016; Wanapat et al., 2018; 2000). The variability of CP content is affected by the leaf to stem ratio, with leaves being higher in CP than stems. Likewise, harvesting age and cutting interval affect the CP content, with the younger leaves having higher CP (Kang et al., 2005).
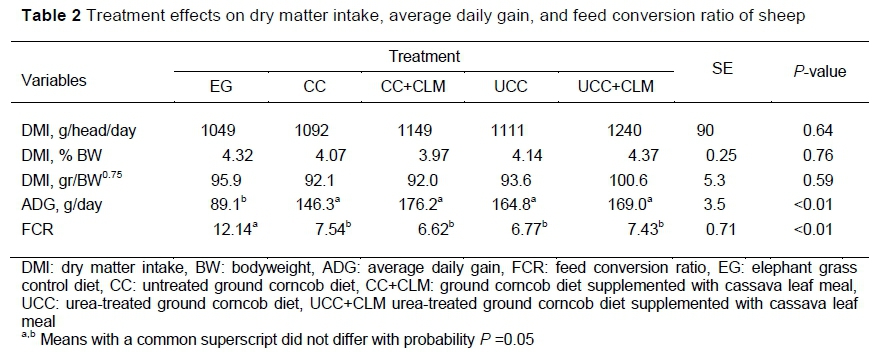
Feed consumption was expressed in g/head/day, as a percentage of bodyweight, and in g/kg metabolic weight. There were no significant treatment effects (P >0.05) on feed consumption. Thus, the use of ground corncob did not reduce palatability of these diets, which implied that it could be used as a fibre source to replace elephant grass in a basal diet. Ground corncob was also used in place of rice straw as a fibre source in diets for dairy cows without affecting milk production (Wachirapacorn et al., 2016). Supplementation with 5% CLM did not increase DMI. Phesatcha et al. (2016) observed that 6% cassava hay meal supplementation had no effect on DMI. Even levels of more than 20% cassava leaf did not increase DMI (Wanapat et al., 2000; Ampapon et al., 2016; Wanapat et al., 2018; Viennasay et al., 2018). However, Odusanya et al. (2017) found that inclusion of 20% CLM in the concentrate diet produced higher DM intake by West African dwarf rams compared with those fed 0%, 10%, and 30% CLM. Feed consumption, ADG, and feed conversion from the growth trial are presented in Table 2.
The ADG of sheep fed EG was lower (P <0.05) than those fed the CC, UCC, CC+CLM and UCC+CLM diets, which produced similar levels of performance (Table 2). Similarly, FCR was improved with these diets compared with EG, which indicated that corncob untreated or treated with urea could be used as a fibre source to increase bodyweight gain of sheep after weaning. Negewo et al. (2018) used urea-treated corncob as a fibre source to study the effects of supplemental Noug seed cake and wheat bran in Arsi-Bale sheep. Ososanya et al. (2013) observed that the highest weight gain and most efficient utilization were achieved by feeding a diet of 66.7% cowpea husk and 33.3% corncob to West African dwarf sheep. The ADG achieved in the current study was higher than that reported by Ososanya et al. (2013). This disparity in response may be because of differences in the level of cassava leaf supplementation, in diet composition and in the breed of the sheep. In contrast, Yulistiani and Puastuti (2012) reported that sheep fed a diet containing urea-treated corncob had similar ADG to those fed a diet containing grass. The higher ADG in the current study than in Yulistiani and Puastuti (2012) may be attributed to the ratios of corncob and concentrate. In the previous study, the ratio was 50% to 50%, whereas in the present study it was 40% to 60%. In addition, the corncob in the current study was of better quality, as was revealed by its higher CP content. The higher ADG of sheep fed the treatment diets and the similarity in feed consumption in the current study resulted in better (P <0.05), the responses in FCR corresponding to those in ADG (Table 2). Lunsin et al. (2012) also reported that cassava hay supplementation to a diet consisting of concentrate and urea-lime-treated rice straw basal diet did not increase milk production, but it affected milk composition in lactating dairy cows. However, Wanapat et al. (2018) reported that cassava top silage supplementation to total mixed ration that included rice straw did not affect DMI but increased milk production. On the other hand, Odusanya et al. (2017) reported that supplementation of CLM at 20% in the Panicum maximum basal diet increased ADG by 13.5%. In the current study, the lack of response in ADG to cassava leaf supplementation may have resulted from the unsupplemented rations being sufficient to meet the nutritional requirements of the animals.
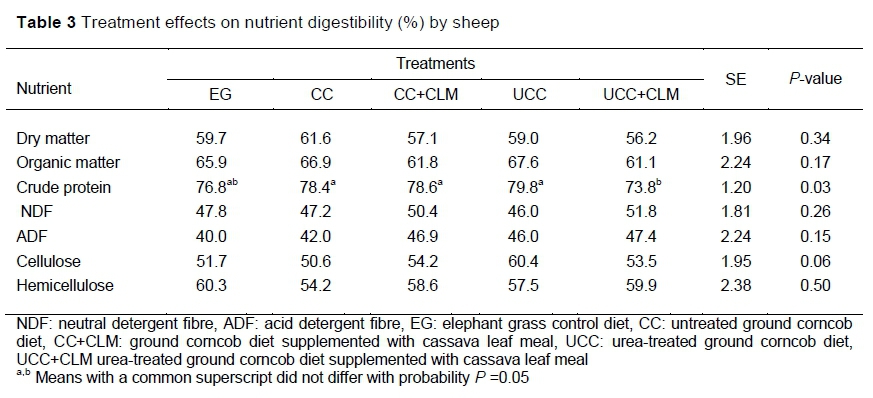
There was no significant difference (P >0.05) in nutrient digestibility among treatments except for CP. The CP digestibility of UCC+CLM was the lowest (P <0.05) among the treatments, but did not differ from EG (P >0.05). In contrast, the effect of urea treatments on corncob (Oji et al., 2007), rice straw (Gunun et al., 2013; Zhang et al., 2019) and wheat straw (Yadete, 2014) increased nutrient digestibility. The difference between this study and those just referenced could be because of the diets. In the current study, the ration was formulated to meet the nutrient requirements (Kearl, 1982) of growing sheep weighing 15 kg, which provided sufficient nitrogen and fermentable carbohydrates to create optimal rumen function for microbial protein synthesis and for fibre degradation in all treatments. The effects of the treatments on nutrient digestibility are presented in Table 3.
The addition of cassava leaves did not increase nutrient digestibility (Table 3). Currently, there appears to be no information about the supplementation of diets that incorporate corncob as a fibre source with cassava leaves. However, wilted cassava leaf supplementation of low-quality gamba grass (Andropogon gayanus) increased CP, NDF, and ADF digestibility (Phengvichith & Ledin, 2007). Lunsin et al. (2012) also reported that the supplementation of cassava hay to urea-lime-treated rice straw increased NDF digestibility. The lack of effects on digestibility in the present study might be because of the lower level of cassava that was fed.
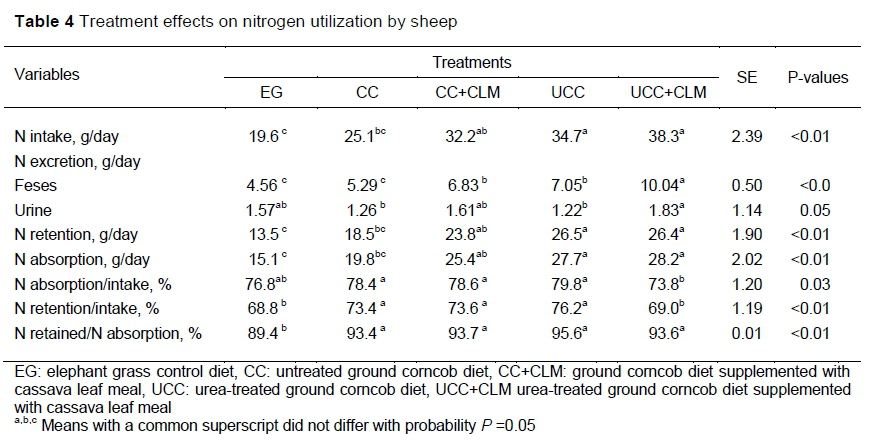
The N intake in CC+CLM, UCC, and UCC+CLM did not differ (P >0.05). Sheep fed the UCC and UCC+CLM diets consumed more N (P <0.05) than EG and CC. Sheep that were fed the UCC, CC+CLM and UCC+CLM diets excreted more N in faeces than EG and CC. Higher faecal N because of urea treatment of feedstuffs was reported for rice straw (Yulistiani et al., 2003; Gunun et al., 2013; Zhang et al., 2019) and wheat straw (Nurfeta et al., 2009; Yadete, 2013). Ammonia bound to the cell wall of urea-treated roughages is more readily absorbed in the hindgut and could account for the higher level of faecal nitrogen with an ammoniated diet (Rath et al., 2001). The higher bacterial counts in sheep fed ammoniated straw compared with those fed untreated rice straw may contribute to higher faecal N (Cann et al., 1991). Faecal nitrogen is produced from bacterial nitrogen and from nitrogen bound to the undegraded cell wall. Hassen and Chenost (1992) reported more nitrogen bound to the cell wall of ammoniated straw than with untreated straw. The cell wall that was not degraded in the rumen would be fermented in the large intestine and that partly undegraded cell wall would pass the large intestine, resulting in the higher faecal N excretion in the UCC diet. Treatment effects on nitrogen utilization of sheep are presented in Table 4.
Cassava leaf meal supplementation of CC and UCC increased faecal N excretion (P <0.05) by 29.1 and 42.4 %. The UCC+CLM diet produced the highest (P <0.05) N excretion among the treatments, which was consistent with it having the highest CP content and the lower observed digestibility of CP. in the past, the effects of feeding cassava leaves on N excretion in faeces were inconsistent. Ukanwoko et al. (2009) and Thang et al. (2010) reported that cassava leaf supplementation increased N excretion in the faeces when feeding cassava peel and rice straw, respectively. On the other hand, cassava leaf supplementation of urea-treated rice straw that was fed with molasses (Hue et al., 2008) and increasing the level of cassava leaf supplementation from 10% to 30% on a Panicum maximum diet Odusonya et al., 2016) did not affect faecal N excretion. Therefore, these inconsistent effects might be attributed to the difference in rate and form of feeding cassava leaf and composition of the diets.
Urinary N from sheep fed UCC+CLM, EG and CC+CLM diets did not differ (P >0.05). However, only the urinary N of sheep fed UCC+CLM was significantly higher than those fed the CC and UCC diets. Nitrogen excretion in the urine of sheep fed the CC and UCC diets was similar. Cassava leaf meal supplementation increased N excretion in the urine produced by the sheep that were fed CC+CLM and UCC+CLM by 27.7% and 50%, respectively (Table 4). According to Hristov et al. (2004), protein in the rumen was degraded into amino acid, peptide and ruminal NH3-N. The N excreted in urine resulted when ruminal NH3-N concentration was in excess of the needs of the rumen microbes and was not incorporated in microbial protein. This excess ruminal NH3-N can be converted to urea in the liver and excreted in the urine (Hristov et al., 2004).
The N retained by sheep fed the UCC and UCC+CLM diets was similar (P >0.05) and higher (P <0.05) than EG and CC. Nitrogen retention by sheep fed CC+CLM differed only (P <0.05) from that the EG diet. The proportion of N retained relative to N intake of sheep fed the UCC+CLM diet was the lowest (P <0.05) but did not differ from EG (P >0.05). In the present study, the efficiency of N utilization relative to intake was high (69 - 79 %) for all diets (Table 4) and more than 90% of digested N was retained in the body. These results suggested that CLM supplementation is not needed in diets that contain about 40% corncob.
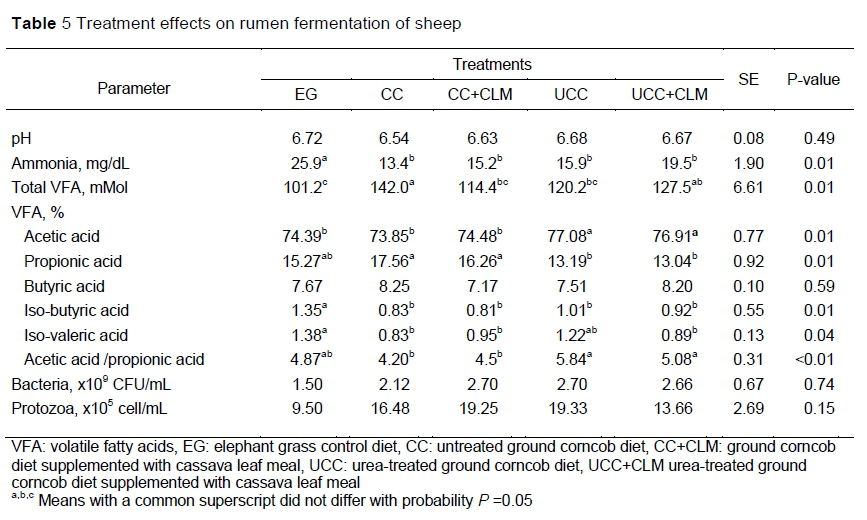
The optimal rumen function because of the treatments was indicated by the similarity in ruminal ammonia concentration and bacterial population (Table 5). Rumen pH was between 6.54 and 6.72, which reflected ideal conditions for rumen microbial activity and the digestion of these diets. Wanapat and Cherdong (2009) reported that optimum ruminal pH for digestion of fibre was 6.5 - 7.0 when feeding high roughage diets. Huhtanen et al. (2006) indicated that increasing the proportion of concentrate in the diet from 25% to 50% lowered rumen pH from 6.43 to 6.21 and resulted in a reduction NDF digestibility. Wachirapakorn et al. (2016) reported that a diet with 40% ground corncob and 60% concentrate fed to crossbred dairy cows produced a ruminal pH of 6.8 - 6.9. In the current study, CLM supplementation did not affect ruminal pH. For lactating dairy cows, similar results were reported by Lunsin et al. (2012) and Wanapat et al. (2018) using cassava hay and cassava top silage as supplements, respectively.
Ruminai NH3-N concentration in EG was significantly higher (P <0.05) than in any other diet. Ruminal NH3-N supplied from degradation of rumen-degradable protein or NPN is an important nutrient in the rumen for protein microbial synthesis and for stimulating ruminal fermentation efficiency (Satter & Slyter, 1974; Cherdthong et al., 2011). Satter and Slyter (1974) suggested for optimum microbial fermentation, NH3-N concentration should be at least 5 mg/dl. Whereas, Erdman et al. (1986) suggested a higher level of NH3-N would be needed for maximum rate of fermentation and Preston and Leng (1987) found that a rumen NH3-N level of 15 - 20 mg/dl was needed to increase intake and digestibility of cattle fed low-quality roughage such as straw. The higher rumen NH3-N in EG may have been because fresh forage was fed, and mastication solubilized its protein by up to 56 - 65% (Ulyatt et al., 1975). Yulistiani (2016) reported that ruminal NH3-N of sheep fed fresh chopped elephant grass was higher than a corncob silage-based diet. In this study, ruminal NH3-N concentration was 13.4 - 19.5 mg/dL which was more than adequate to promote increased feed intake and fibre digestibility.
Total volatile fatty acid concentration in CC was higher (P <0.05) than for the other diets, but did not differ (P >0.05) from UCC+CLM. In contrast, the proportion of propionic acid was significantly lower in the UCC and UCC+CLM diets compared with the CC and CC+CLM, but did not differ from EG. Butyric acid was similar (P >0.05) among treatments. The observation that cassava leaf supplementation did not increase VFA production was not reported in sheep. However, it was reported in dairy cows when cassava top silage replaced rice straw (Viennassay et al., 2018), supplemented rice straw (Wanapat et al., 2018), and supplemented urea-lime-treated rice straw (Lunsin et al., 2012). The higher VFA production and more efficient use of N with the CC diet composed of about 40% corncob compared with EG could be viewed as a positive outcome for sheep feeding and further the finding that it was not necessary to use urea to increase corncob quality was favourable because this method is more practical and cheaper.
Ruminal cellulolytic bacteria play a crucial role in fermenting fibrous forage diets. In the current study, CLM supplementation was expected to provide additional non-ammonia N to improve fibre digestibility in particular and to increase microbial synthesis and animal performance. However, digestibility of NDF, ADF, cellulose and hemicellulose was not increased with CLM supplementation. Thus, the current findings contradict previous studies, which could be associated with different N sources, deriving from ruminal NH3-N and non-ammonia nitrogen. Russell et al. (1992) stated that bacterial fermentation of structural carbohydrates used only ruminal NH3-N as the source of N. However, Carro and Miller (1999) used non-ammonia N (protein, peptide or amino acid) isolated from soybean meal with sufficient energy availability in in vitro and in vivo studies and demonstrated increased fibre degradability, total VFA production, and microbial protein synthesis. Zhang et al. (2012) also used non-ammonia N in an in vitro study in the form of valine, leucine and iso-leucine to increase VFA and NDF degradability from a wheat straw substrate. Zain et al. (2008) used ammoniated palm press fibre supplemented with amino acids (valine, leucine, and iso-leucine) and methionine hydroxyl analogue in a total mixed ration and reported increased feed digestibility, total VFA production, bacterial counts and live weight gain in sheep.
In the current study, microbial protein synthesis of sheep fed CC+CLM and UCC+CLM was not different from other diets, as indicated by similar bacterial counts. Ruminal NH3-N derived from cassava leaves was not used for rumen microbial synthesis, as was revealed by the higher N excretion in the urine, which could be related to availability of fermentable energy in the rumen providing energy and carbon skeletons in support of the synthesis of ruminant microbial protein (Hoover & Stokes, 1991). Thus, it was rationalized that NH3-N derived from the N in cassava leaves was not balanced by available energy. Second, fermentation of cassava leaves did not improve fibre digestibility and increase VFA production because of the lack of effect on the microbial population. Similarly, Phesatcha et al. (2016) found that cassava leaf hay, which made up 6% of the diet, did not enhance rumen fermentation. In contrast, studies using non-ammonia nitrogen sources increased fibre digestibility and rumen fermentation because they used N supplementation from pure branch-chain amino acids and isolated branch-chain amino acids (BCAA) from soybean (Carro & Miller, 1999; Zain et al., 2008; Zhang et al., 2012).
Volatile fatty acids are products of fermentation of feedstuffs by rumen microbes. The lack of effects on the populations of bacteria and protozoa in the sheep fed all treatments, except for EG, may have resulted in the similarity of their VFA production. In contrast, Wanapat et al. (2009) reported that VFA production of a urea-treated rice straw basal diet was higher than untreated rice straw because of increased nutrient digestibility. The higher VFA production in CC in the current study was because of the lower ADF content of the corncob, with most of the cell wall component being rumen-fermentable hemicellulose (Yulistiani et al., 2012). Gunun et al. (2013) observed that lactating cows had similar VFA production when fed diets containing untreated and urea-treated rice straw. Supplementation with branch-chain amino acids (BCAA) increased VFA production because of accelerated substrate degradation and BCAA metabolism (Zhang et al., 2012). However, the lack of effect on VFA production in the current study might be because BCAA could not stimulate microbial growth. The acetic acid proportion was higher in sheep fed the urea enhanced diets than in EG, CC and CC+CLM because of the tendency (P <0.06) towards higher digestibility of cellulose. Fermentation of nonstructural carbohydrates produces acetic acid (Firkin et al., 2006). Iso-butyric and iso-valeric acids are the products of fermentation of BCAA that are essential growth factors for most fibre-degrading microorganisms in the rumen (Yang, 2002). The results of the current study indicated that CLM supplementation was insufficient to increase supply of BCAA for the cellulolytic bacteria.
Conclusion
It was not necessary to apply urea to increase the quality of a diet consisting of 40% corncob. Use of CLM to supplement these diets also did not increase sheep performance. Sheep fed corncob as a fibre source had better growth rate and feed efficiency than those fed a diet that incorporated elephant grass, which suggested the viability of corncob as an alternative fibre source.
Author's Contributions
DY and WP were in charge of project design and project implementation. DY, WP and EW analysed data, interpreted the data, and wrote the manuscript. This manuscript was read and approved by all of the authors
Conflict of Interest Declaration
There is no conflict of interest related to this manuscript
References
Ampapon, T., Wanapat, M. & Kang, S., 2016. Rumen metabolism of swamp buffaloes fed rice straw supplemented with cassava hay and urea. Trop. Anim. Health Prod. 48, 779-784. DOI: 10.1007/s11250-016-1026-5 [ Links ]
AOAC, 2012. Official methods of analysis. 19th ed. Association of Official Analytical Chemists, Gaithersburg, MD. [ Links ]
Arroquy, J.I., Cochran, R.C., Wickersham, T.A., Llewellyn, D.A., Titgemeyer, E.C., Nagaraja, T.G. & Johnson, D.E., 2004. Effects of type of supplemental carbohydrate and source of supplemental rumen degradable protein on low quality forage utilization by beef steers. Anim. Feed Sci. Technol. 115, 247-263. DOI:10.1016/j.anifeedsci.2004.01.007 [ Links ]
Cann, I.K.O., Kobayashi, Y., Wakita, M. & Hoshino, S., 1991. Digestion properties of ammoniated rice straw in the rumen and lower tract of sheep. Anim. Feed Sci. Technol. 35, 55-68. DOI: 10.1016/0377-8401 (91)90099-E [ Links ]
Carro, M.D. & Miller, E.L., 1999. Effect of supplementing a fiber basal diet with different nitrogen forms on ruminal fermentation and microbial growth in an in vitro semicontinuous culture system (RUSITEC). Br. J. Nutr. 82(2), 149-157. DOI: 10.1017/S0007114599001300 [ Links ]
Chenost, M. & Kayouli, C. 1997. Roughage utilization in warm climates. FAO Animal Production and Health Paper 135. Rome. [ Links ]
Cherdthong, A., Wanapat, M. & Wachirapakorn, C., 2011. Influence of urea calcium mixture supplementation on ruminal fermentation characteristics of beef cattle fed on concentrates containing high levels of cassava chips and rice straw. Anim. Feed Sci. Technol. 163, 43-51. DOI: 10.1016/j.anifeedsci.2010.10.003 [ Links ]
Conway, E.J., 1962. Microdiffusion analysis and volumetric error. 5th edition. Crosby Lockwood, London [ Links ]
Erdman, R.A., Proctor, G.H. & Vandersall, J.H., 1986. Effect of rumen ammonia concentration on in situ rate and extent of digestion of feedstuffs. J. Dairy Sci. 69(9), 2312-2320. DOI: 10.3168/jds.S0022-0302(86)80670-1 [ Links ]
Firkin, J.L., Hristov, A.N., Hall, M.B., Varga, G.A. & St-Pierre, N.R., 2006. Integration of ruminal metabolism in dairy cattle. J. Dairy Sci. 89 (suppl.), E31-E51. DOI: 10.3168/jds.S0022-0302(06)72362-1 [ Links ]
Gunun, P., Wanapat, M. & Anantasook, N., 2013. Effects of physical form and urea treatment of rice straw on rumen fermentation, microbial protein synthesis and nutrient digestibility in dairy steers. Asian Australas. J. Anim. Sci. 26(12), 1689-1697. DOI: 10.573/ajas.2013.13190 [ Links ]
Hassen, L. & Chenost, M., 1992. Tentative explanation of the abnormally high faecal nitrogen-excretion with poor-quality roughages treated with ammonia. Anim. Feed Sci. Technol. 38, 25-34. DOI: 10.1016/0377-8401(92) 90073-F [ Links ]
Hoover, W.H. & Stokes, S.R., 1991. Balancing carbohydrates and proteins for optimum rumen microbial yield. J. Dairy Sci. 74(10), 3630-3644. DOI: 10.3168/jds.S0022-0302(91)78553-6 [ Links ]
Hristov, A.N., Etter, R.P., Ropp, J.K. & Grandeen, K.L., 2004. Effect of dietary crude protein level and degradability on ruminal fermentation and nitrogen utilization in lactating dairy cows. J. Anim. Sci. 82(11), 3219-3229. DOI: 10.2527/2004.82113219x [ Links ]
Hue, K.T., Thanh Van, D.T. & Ledin, I., 2008. Effect of supplementing urea treated rice straw and molasses with different forage species on the performance of lambs. Small Rum. Res. 78, 134-143. DOI: 10.1016/j.smallrumres.2008.05.010 [ Links ]
Huhtanen, P., Ahvenjarvi, S., Weisbjerg, M.R. & Norgard P., 2006. Digestion and passage of fibre in ruminants. In: Ruminant physiology, digestion, metabolism and impact of nutrition on gene expression, immunology and stress. Pp. 87-135. Wageningen Academic Publishers. [ Links ]
Jaswandi, H. & Jhon, F., 2019. An ammoniated rice straw cow diet supplemented with cassava leaves and synchronized release of N-protein and energy in the rumen. Pak. J. Nutr. 18(6), 501-507. DOI: 10.3923/pjn.2019.501.507 [ Links ]
Kayastha, T.B., Dutta, S., Kayastha, R.B. & Deka, R.S. 2012. Growth performance and nutrient utilization of growing calves with urea treated wheat straw based ration. Indian J. Dairy Sci. 65(5), 435-438. http://epubs.icar.org.in/ejournal/index.php/IJDS/article/view/25439 [ Links ]
Kearl, L.C., 1982. Nutrient requirements of ruminants in developing countries. International Feedstuff Institute, Utah Agricultural Experiment Station, Utah State University. USA. [ Links ]
Khan, M.A., Sarwarand, M. & Khan. M.S., 2004. Feeding value of urea treated corncobs ensiled with or without enzose (corn dextrose) for lactating crossbred cows. Asian-Aust. J. Anim. Sci. l 17 (8), 1093-1097. https://doi.org/10.5713/ajas.2006.705 [ Links ]
Khang, D.N., Wiktorsson, H. & Preston, T.R., 2005. Yield and chemical composition of cassava foliage and tuber yield as influenced by harvesting height and cutting interval. Asian-Aust. J. Anim. Sci. 18 (7): 1029-1035 DOI: https://doi.org/10.5713/ajas.2005.1029 [ Links ]
Lunsin, R., Wanapat, M. & Rowlinson, P., 2012. Effect of cassava hay and rice bran oil supplementation on rumen fermentation, milk yield and milk composition in lactating dairy cows. Asian-Aust. J. Anim. Sci. 25(10), 1364-1373. DOI: 10.5713/ajas.2012.12051 [ Links ]
McGuire, D.L., Bohnert, D.W., Schauer, C.S. Falck, S.J. & Cooke, R.F., 2013. Daily and alternate day supplementation of urea or soybean meal to ruminants consuming low-quality cool-season forage: I. Effects on efficiency of nitrogen use and nutrient digestion. Livest. Sci. 155(2-3), 205-213. DOI: 10.1016/j.livsci.2013.05.015 [ Links ]
National Research Council (NRC). 2011. Guide for the care and use laboratory animals. Eighth edition. The National Academic Press, Washington DC. [ Links ]
Naves, J.R., de Jesus, E.F., Martins, C.M.M.R., Tomazi, T., de Freitas, J.E. Junior, Rennó, F.P. & Santos, M.V., 2015. Efficiency of dietary nitrogen utilization and digestive metabolism of dairy cows fed different nitrogen sources and sugarcane. J. Anim. Plant Sci. 25(4), 941-949. https://www.researchgate.net/publication/281969954_Efficiency_of_dietary_nitrogen_utilization_and_ digestive_metabolism_of_dairy_cows_fed_different_nitrogen_sources_and_sugarcane [ Links ]
Negewo, T., Melaku, S., Asmare, B. & Tolera, A., 2018. Performance of Arsi-Bale sheep fed urea treated maize cob as basal diet and supplemented with graded levels of concentrate mixture. Trop. Anim. Health and Prod. DOI: 10.1007/s11250-018-1544-4 [ Links ]
Nurfeta, A., Tolera, A., Eik, L.O. & Sundst0l, F., 2009. Feeding value of enset (Ensete ventricosum) leaf supplementation on feed intake, digestibility, nitrogen utilization and bodyweight gain of sheep fed hay and untreated or urea and calcium oxide treated wheat straw for sheep. J. Anim. Phys. Anim. Nut. 93(1), 94-104. DOI: 10.1016/j.livsci.2008.08.005 [ Links ]
Odusanya, L.Q., Fasae, O.A., Adewumi, O.O. & James, I.J., 2017. Effect of cassava leaf meal concentrate diets on the performance, haematology and carcass characteristics of West African dwarf lambs. Arch. de Zootec. 66 (256), 603-609. DOI: 10.21071/az.v66i256.2779 [ Links ]
OECD/FAO, 2019. OECD-FAO Agricultural Outlook. OECD Agriculture statistics. Chapter 3: Cereals. DOI: dx.doi.org/10.1787/agr-outl-data-en. http://www.agri-outlook.org/commodities/Cereals.pdf [ Links ]
Ogimoto, K. & Imai, S., 1981. Atlas of rumen microbiology. Japan Scientific Societies Press, Tokyo. [ Links ]
Oji, U.I., Etim, H.E. & Okoye, F.C., 2007. Effects of urea and aqueous ammonia treatment on the composition and nutritive value of maize residues. Small Rum. Res. 69, 232-236. DOI: 10.1016/j.smaNrumres.2006.01.015 [ Links ]
Oni, A.O., Onwuka, C.F.I., Arigbede, O.M., Oni, O.O., Anele, U.Y., Yusuf, K.O., Oduguwa, B.O. & Onifade, O.S., 2010. Chemical composition and in sacco degradability of four varieties of cassava leaves grown in Southwestern Nigeria in the rumen of sheep. Trop. Anim. Health Prod. 42,1385-1393. DOI: 10.1007/s11250-010-9596-0 [ Links ]
Ososanya, T.O., Alabi, B.O. & Sorunke, A.O., 2013. Performance and digestibility of corncob and cowpea husk diets by West African dwarf sheep. Pak. J. Nut. 12 (1), 85-88. DOI: 10.3923/pjn.2013.85.88 [ Links ]
Phesatcha, B., Wanapat, M., Phesatcha, K., Ampapon, T. & Kang, S. 2016. Supplementation of Flemingia macrophylla and cassava foliage as a rumen enhancer on fermentation efficiency and estimated methane production in dairy steers. Trop Anim. Health Prod. 48:1449-1454. DOI 10.1007/s11250-016-1115-5 [ Links ]
Phengvichith, V. & Ledin, I., 2007. Effect of feeding different levels of wilted cassava foliage (Manihot esculenta Crantz) on the performance of growing goats. Small Rum. Res. 71, 109-116. DOI: 10.1016/j.smallrumres.2006.05.009 [ Links ]
Preston, T.R. & Leng, R.A., 1987. Matching ruminant production systems with available resources in the tropics and sub-tropics. Renambul, Armidale, New South Wales, Australia. [ Links ]
Rath, S., Verma, A.K., Singh, P., Dass, R.S. & Mehra, U.R., 2001. Performance of growing lambs fed urea ammoniated and urea supplemented wheat straw-based diets. Asian-Aust. J. Anim. Sci. 14(8), 1078-1084. DOI: 10.5713/ajas.2001.1078 [ Links ]
Ravindran, V., 1993. Cassava leaves as animal feed: Potential and limitations. J. Sci. Food Agric. 61(2), 145-150. DOI: 10.1002/jsfa.2740610202 [ Links ]
Russell, J.B., O'Connor, J.D., Fox, D.G., Van Soest, P.J. & Sniffen, C.J., 1992. A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. J. Anim. Sci. 70 (11): 3551-3561. DOI: 10.2527/1992.70113551x [ Links ]
Satter, L.D. & Slyter, L.L., 1974. Effect of ammonia concentration on rumen microbial protein production in vitro. Br. J. Nut. 32(2), 199-208. DOI: 10.1079/BJN19740073 [ Links ]
Thang, C.M., Ledin, I. & Bertilsson, J. 2010. Effect of feeding cassava and/or Stylosanthes foliage on the performance of crossbred growing cattle. Trop. Anim. Health Prod. 42:1-11. DOI: 10.1007/s11250-009-9378-8 [ Links ]
Ukanwoko, A.I., Ahamefule, F.O. & Ukachukwu, S.N., 2009. Nutrient intake and digestibility of West African dwarf bucks fed cassava peel-cassava leaf meal based diets in South Eastern Nigeria. Pak. J. Nut. 8(7), 983-987. DOI: 10.3923/pjn.2009.983.987 [ Links ]
Ulyatt, M.J., McRae, J.C., Clarke, T.J. & Pearce, P.D., 1975. Quantitative digestion of fresh forages by sheep. IV. Protein synthesis in the stomach. J. Agric. Sci. 84(3), 453-458. DOI: 10.1017/S0021859600052655 [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber neutral detergent fiber and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74(10), 3583-3597. DOI: 10.3168/jds.S0022-0302(91)78551-2 [ Links ]
Viennasay B., Wanapat, M., Phesatcha, C.K., Phesatcha, B. & Ampapon,T., 2018. Replacement of rice straw with cassava-top silage on rumen ecology, fermentation and nutrient digestibilities in dairy steers. Anim. Prod. Sci. https://doi.org/10.1071/AN17477. [ Links ]
Wachirapakorn, C., Pilachai, K., Wanapat, M., Pakdee, P. & Cherdong, A., 2016. Effect of ground corncobs as a fiber source in total mixed ration on feed intake, milk yield and milk composition in tropical lactating crossbred Holstein cows. Anim. Nut. 2(4), 334-338. DOI: 10.1016/j.aninu.2016.08.007 [ Links ]
Wanapat, M. & Cherdthong, A., 2009. Use of real-time PCR technique in studying rumen cellulolytic bacteria population as affected by level of roughage in Swamp buffalo. Curr. Microbio. 58(4), 294-299. DOI: 10.1007/s00284-008-9322-6 [ Links ]
Wanapat, M., Puramongkon, T. & Siphuak, W., 2000. Feeding of cassava hay for lactating dairy cows. Asian-Aust. J. Anim. Sci. 13(4), 478-482. DOI: 10.5713/ajas.2000.478 [ Links ]
Wanapat, M., Polyorach, S., Boonnop, K., Mapato, C. & Cherdthong. A., 2009. Effects of treating rice straw with urea or urea and calcium hydroxide upon intake, digestibility, rumen fermentation and milk yield of dairy cows. Livest. Sci. 125(2-3), 238-243. DOI: 10.1016/j.livsci.2009.05.001 [ Links ]
Wanapat, M., Phesatcha,K., Viennasay, B., Phesatcha, B., Ampapon, T. & Kang, S., 2018. Strategic supplementation of cassava top silage to enhance rumen fermentation and milk production in lactating dairy cows in the tropics Trop. Ani. Health Prod. DOI: 10.1007/s11250-018-1593-8 [ Links ]
Yadete, G.K., 2014. Effect of wheat straw urea treatment and Leucaena leucocephala foliage hay supplementation on intake, digestibility, nitrogen balance and growth of lambs. International J. Livest. Prod. 6 (4), 88-96. DOI:10.5897/IJLP12.040 [ Links ]
Yang, C.M., 2002. Response of forage fiber degradation by ruminal microorganisms to branched-chain volatile fatty acids, amino acids, and dipeptides. J. Dairy Sci. 85(5), 1183-1190. DOI: 10.3168/jds.S0022-0302(02)74181-7 [ Links ]
Yulistiani, D., 2016. Response of sheep fed on corncob silage or elephant grass basal diet with or without Calliandra leaf meal supplementation. Indonesian J. Anim. Vet. Sci. 21(3): 165-173. DOI: 10.14334/jitv.v21i3.1574 [ Links ]
Yulistiani, D. & Puastuti, W., 2012. Feed consumption and growth response of lambs fed on different basal diet. Proc. of the 15th AAAP Anim. Sci. Cong., 23-30 November 2012, Thammasat University, Thailand. Pp. 733-736. [ Links ]
Yulistiani, D., Gallagher, J.R. & Van Barneveld, R.J., 2003. Intake and digestibility of untreated and urea treated rice straw based diet fed to sheep. Indonesian J. Anim. Vet. Sci. 8 (1), 8-16. http://oaji.net/articles/2015/1610-1424061569.pdf [ Links ]
Yulistiani, D., Puastuti, W., Wina, E. & Supriati, 2012. Effect of processing on nutritive value of corncob: Chemical composition and in vitro digestibility. Pengaruh berbagai pengolahan terhadap nilai nutrisi tongkol jagung: Komposisi kimia dan kecernaan in vitro. (In Bahasa). Indonesian J. Anim. Vet. Sci. 17(1), 59-66. DOI: 10.14334/jitv.v17i1.712 [ Links ]
Zain, M., Sutardi, T., Suryahadi & Ramli, N., 2008. Effect of defaunation and supplementation methionine hydroxy analogue and branched chain amino acid in growing sheep diet based on palm press fiber ammoniated. Pakistan J. of Nut. 7 (6), 813-816. DOI: 10.3923/pjn.2008.813.816 [ Links ]
Zhang, H.L., Chen, Y., Xu, X.L. & Yang. Y.X., 2012. Effects of branched-chain amino acids on in vitro ruminal fermentation of wheat straw. Asian-Austr. J. Anim. Sci.26(4), 523-528. DOI: 10.5713/ajas.2012.12539 [ Links ]
Zhang X., Medrano R.F., Wang M., Beauchemin K.A., Ma Z., Wang R., Wen J., Bernard L.A. & Tan Z., 2019. Effects of urea plus nitrate pretreated rice straw and corn oil supplementation on fiber digestibility, nitrogen balance, rumen fermentation, microbiota and methane emissions in goats. J. Anim. Sci. Biotech. DOI: 10.1186/s40104-019-0312-2 [ Links ]
Submitted 2 May 2021
Accepted 6 September 2021
Published 20 November 2021
# Corresponding author's email: dwiyulistiani@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}