










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 no.6 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i6.1
ARTICLES
Yeast culture (Saccharomyces cerevisiae) and its active metabolites affect the cecal microbiome of broilers
Z. SunI, III; Y. ZhenII, III; T. LiII; N.D. AschalewII; T. WangII, III; X. ChenII, III; W. ZhaoII, III; X. ZhangII, III, #; G. QinII, #
ICollege of Life Science, Jilin Agricultural University, Changchun 130118, China
IICollege of Animal Science and Technology, JLAU-Borui Dairy Science and Technology R&D Centre, Jilin Agricultural University, Changchun 130118, P.R. China
IIIPostdoctoral Scientific Research Workstation, Feed Engineering Technology Research Centre of Jilin Province, Changchun Borui Science & Technology Co., Ltd, Changchun 130118, P.R. China
ABSTRACT
Yeast cultures (YCs) are defined as promising feed additives that maintain the health of birds and improve growth performance by modulating gut microbiota. YCs contain effective metabolites such as glycine, fructose, inositol, galactose, and sucrose. This study investigated the effects of YCs and their effective metabolites on carcass traits and cecal microflora in broilers. A total of 280 one-day-old mixed-sex Arbor Acres broilers were randomly allocated to seven groups. The basal diet (control DZ) was supplemented with various proportions of glycine, fructose, inositol, galactose, and sucrose (Groups A, B, and C), 24-hour grown Saccharomyces cerevisiae cultures (Group D) (YC), and a commercial yeast culture product (SZ) at concentrations of 0.1% and 1% (Groups E and F). Bodyweight of broilers was correlated positively with proportions of Proteobacteria in Group C and Lactobacillus and Roseburia in Group B (P <0.05). Broilers fed diets supplemented with YC or its active metabolites had the highest proportions of bacteria involved in nucleotide metabolism, and amino acid and carbohydrate metabolism. These results suggested that the dietary addition of YC could alter the proliferation of beneficial bacteria in broilers.
Keywords: broiler, carcass traits, microflora, yeast culture
Introduction
Numerous yeast and yeast products have been fed to livestock for more than a century (Stone, 2006; Saied et al., 2011; Bilal et al., 2020). Recently the addition of antibiotics to livestock diets to promote production efficiency has been restricted and has declined. Therefore, research aimed at developing alternatives has increased significantly, with particular emphasis on potential uses of natural feed additives (Chand et al., 2019; Saeed et al., 2019; Tufail et al., 2019). A yeast culture is one such natural additive (Kondo et al., 2014). Yeast culture (YC) is a natural yeast fermentation product, containing a small amount of yeast biomass and a large number of fermentation metabolites. The yeast cells are used as a tool to produce cellular metabolites during production of the YC (Poppy et al., 1995). A number of studies have confirmed that YC can modulate the animal gut microbiota, promote a beneficial balance of the microbial population of the gastrointestinal tract, and inhibit the survival of pathogens, thereby enhancing the immune capacity of animals, and may ultimately play a role in maintaining animal health and improving performance. For instance, dietary YC not only modulated the gut immune response of nursery pigs (Shen et al., 2009), but it also affected intestinal mucosal morphology and ileal villus development in broilers (Zhang et al., 2005; Bai et al., 2013). It has been reported that YC could improve bodyweight gain and feed conversion efficiency (Hooge et al., 2004; Stanley et al., 2004; Shurson et al., 2018; Stanley et al., 2016).
Nutritional and health effects of YC are achieved through the fermentation of extracellular metabolites that have high thermal stability (Shurson et al., 2018; Liu et al., 2018). However, there are many types of metabolites in YC and most have unknown effects (Shurson et al., 2018; Kondo et al., 2014), except for a few with known nutritional value, such as organic acids, oligosaccharides, and amino acids Thus Sun et al. (2020) screened glycine, fructose, inositol, galactose, and sucrose as potential metabolites of YC with gas chromatography-mass spectrometry. Therefore, this study aimed to determine and compare the effects of YC and its active metabolite supplementation on carcass characteristics and the cecal microbiota of broilers to verify further whether dietary supplementation with the effective compounds of YC could alter intestinal health.
Materials and methods
The animal trial was approved by the Experimental Animal Committee of Jilin Agricultural University (JLAU-ACUC2017-007, Changchun, China). This study was conducted at a commercial broiler chicken farm (Changchun, China). A total of 280 one-day-old mixed-sex Arbor Acres broilers were collected and randomly assigned to seven groups, with five replicates for each group, and each replicate consisting of eight broilers (n = 40 broilers/group). The treatments consisted of control group (DZ), in which broilers were fed a basal diet. In Groups A, B, and C broilers were fed a basal diet supplemented with various proportions of glycine, fructose, inositol, galactose, and sucrose. In Group D (YC), broilers were fed a basal diet supplemented with 24-hour- grown Saccharomyces cerevisiae cultures at a concentration of 0.192%. Groups E and F were fed a basal diet supplemented with a commercial yeast culture product (SZ) at concentrations of 0.1% and 1% (Table 1). The components of the basal diet and the relative ratios of the five additives are given in Sun et al. (2020).
The Saccharomyces cerevisiae (JLAU-Borui No. 2012) was obtained from JLAU-Borui Dairy Science and Technology R&D Centre of Jilin Agricultural University (Changchun, China). Its active components (i.e., glycine, fructose, inositol, galactose, sucrose) in 24-hour-grown YC were screened through metabolomic and animal experiments (Sun et al., 2020). The proportions of glycine, fructose, inositol, galactose, and sucrose in the feed were believed to be most beneficial. In Group A the proportions of these components were the same as in 24-hour-fermented YC.
The broilers were raised in wire-floored cages in an environmentally controlled room with continuous light. They had access to feed and water ad libitum. During the first seven days, the temperature was kept at 32 - 35 °C, and then gradually reduced to 22 °C. To evaluate the carcass characteristics, two broilers of similar bodyweight from each replicate were killed at 42 days by cervical dislocation. The weights of the breast, thigh, pancreas and abdominal fat were recorded, and the dressing percentage was calculated relative to the live bodyweight.
Cecal samples were collected from five broilers from each group at 42 days and stored at -80 °C. DNA was isolated from each 220 mg sample with the Fast DNA SPIN Extraction Kit (MP Biomedicals, Santa Ana, CA, USA) and stored at -20 °C. The quantity of the DNA was determined with a Nano Drop ND-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) and the quality was evaluated with agarose gel electrophoresis.
Polymerase chain reaction (PCR) amplification of V3-V4 regions of the bacterial 16S rRNA gene was performed with the forward primer 515F (5'-GTGCCAGCMGCCGCGGTAA-3') and the reverse primer 907R (5'-CCGTCAATTCMTTTRAGTTT-3'). The PCR cycling conditions consisted of an initial denaturation at 98 °C for 2 min, followed by 25 cycles of denaturation at 98 °C for 15 seconds, annealing at 55 °C for 30 seconds, and extension at 72 °C for 30 seconds, with a final extension of 5 min at 72 °C. PCR amplicons were purified with Agencourt AMPure beads (Beckman Coulter, Indianapolis, IN, USA) and quantified with the PicoGreen dsDNA assay kit (Invitrogen, Carlsbad, CA, USA). Equal amounts of the amplicons were pooled, and pair-end 2 *300 bp sequencing was performed with the Illumina MiSeq platform with the MiSeq reagent kit at Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China). The broiler 16S rRNA gene sequence was deposited in the NCBI SRA database with accession number SRP238826.
Sequence reads were analysed with the quantitative insights into microbial ecology (QIIME, v1.8.0) tool. After chimera detection, the UCLUST algorithm was used to cluster the remaining high-quality sequences into operational taxonomic units (OTUs) at 97% sequence identity. The classification of OTUs was performed by BLAST searching the representative sequences against the Greengenes database (DeSantis et al., 2006) using the best hit. Alpha diversity indices, including Chaol, Shannon, Simpson, and abundance-based coverage estimator (ACE), were used to calculate expected OTUs based on observed OTUs. Principal component analysis (PCA) was performed with genus-level compositional profiles. The taxonomic compositions and proportions were visualized with MEGAN and GraPhlAn software packages. The functions of microbes were assigned by the Kyoto encyclopaedia of genes and genomes (KEGG) tool and predicted by phylogenetic investigation of communities by reconstruction of unobserved states (PICRUSt) software (Langille et al., 2013).
Data were analysed with SPSS 20.0 software (SPSS Inc., Chicago, Illinois, USA) for analysis of variance and results expressed as means ± SE. Groups were compared with Duncan's multiple comparison test, and significance was taken at P <0.05. Linear regression was used to examine the association between the bodyweight and microbial taxa. An absolute value of the correlation coefficient closer to 1 indicated a stronger relationship.
Results and Discussion
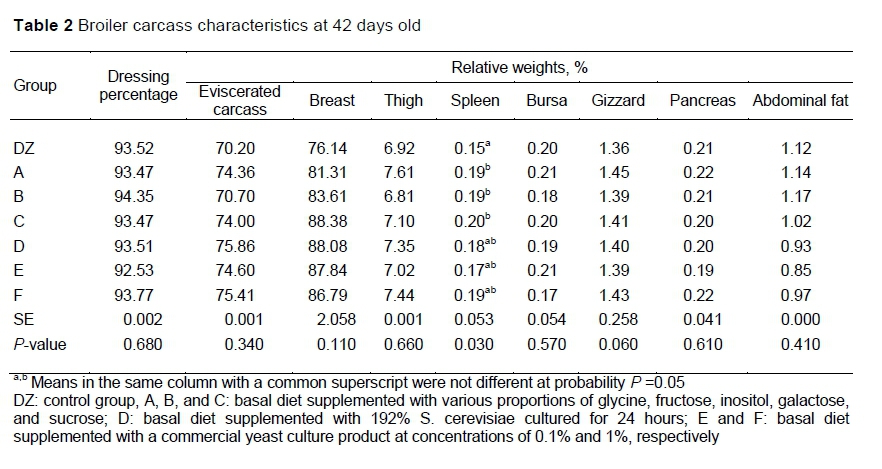
No differences were detected between the groups and the control in the relative weights of the breast, thigh, pancreas, and abdominal fat, and dressing percentage at 42 days old. Relative weight of the spleen was affected significantly by the treatments, with A, B and C having heavier spleens than DZ (P <0.05) (Table 2).
Previous work had shown that improved growth performance was attributable to YC active metabolites, which affected the proportions of amino acids, sugar, and organic acids (Sun et al., 2020). At cellular level, nutrition depends on the feed ingredients provided, the absorption rate of the nutrients, and the digestion of nutritionally active substances that are released and dissolved, and which are absorbed by cells to affect gene expression and cell function. It was reported that carbohydrates and amino acids in feeds could regulate gene expression, thereby facilitating animal health and growth (Adams, 2006). Chickens require dietary sugars, such as sucrose, fructose, and galactose, which could replace corn as the primary energy source (Hussein et al., 2018). Dietary sugars are highly metabolizable, thereby providing instant energy. They also have a pleasing aroma and make feed more palatable, which can increase feed intake in broilers. In animal systems, sucrose, and not starch, is considered the optimal energy source, and glucose can replace the fatty acids in the feed. Wang et al. (2014) reported that in ovo use of glucose and fructose may improve the immunity of chicks. On the other hand, as a nonessential amino acid in animals, glycine works with other amino acids in the synthesis of creatine, glutathione, bile acids, nucleic acids, and uric acids (Corzo et al., 2004). Pirgozliev et al. (2017) also demonstrated that dietary myo-inositol, which is critical to cell survival, can improve the intestinal health and growth of broilers. However, in another study, inositol supplementation did not affect the mineral availability and growth of broiler chickens (Pirgozliev et al., 2018). in addition, the spleen is an important immune organ in birds. Ahiwe et al. (2019) reported an increase in spleen weight after supplementation with yeast and its derivatives in broilers, consistent with the current results, which showed that supplementation with component C increased the spleen weight. These findings may have been caused by increased immune activity in chicken fed YC active metabolites.
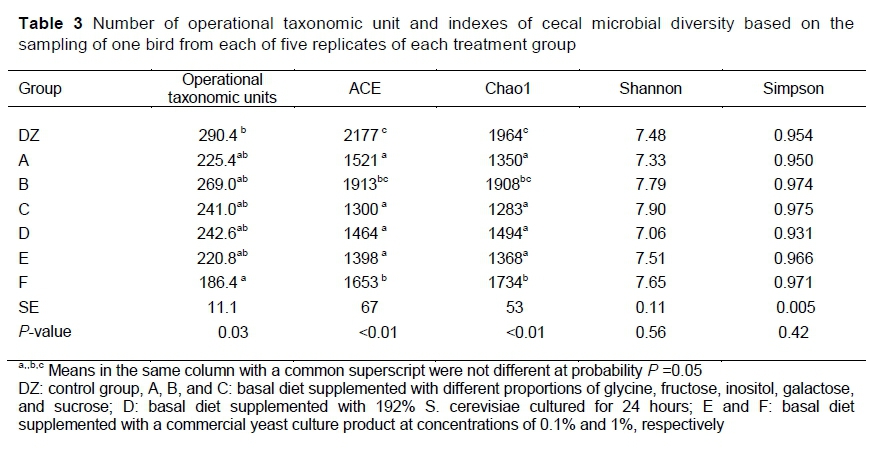
A total of 1,599,212 high quality sequences were generated from V3-V4 regions of the 16S rRNA gene with an average of 45,691 reads per subject (range 27,853 - 157,129). The assembled sequences had an average length of 425 bp. The number of operational taxonomic units (OTUs) and the sample richness and diversity are shown in Table 3. Chao1 and ACE diversity indices were lower in A, C, D, and E than in the control or F (Ρ <0.05). However, there were no significant differences in Shannon and Simpson indices among the groups (Ρ >0.05). Figure 1 shows the relationships by PCA among microbial communities after various treatments. The contribution rates of principal component factor (PC) 1 and PC2 were 32.28% and 24.39%, respectively. The two principal components explained 56.7% of the composition of the microbiota at the genus level. After supplementation with YC or active metabolites, the overall composition of the cecal microbial environment was different from that of the control. However, there was no apparent separation in microbial genera among Groups A, B, C, and E, indicating that active metabolites induced changes that were similar to those induced by the YC product.
Microbial diversity in the gut is important for digestion and nutrient uptake (Xu et al., 2016). The results indicated that active metabolites reduced the complexity of the cecal microbial environment, as indicated by decreased Chao1 and ACE indices. A previous study revealed a negative correlation between bodyweight gain and intestinal microbial diversity (Han, 2018). The results of PCA indicated a separate microbial cluster between metabolite-supplemented and unsupplemented diets of broilers. This indicated that the microbial environment was affected by the metabolites in the YC. In contrast, Ran et al. (2015) illustrated that YC supplementation of the diets of Nile tilapia affected the alpha-diversity of microbiota, but PCA revealed no significant difference.
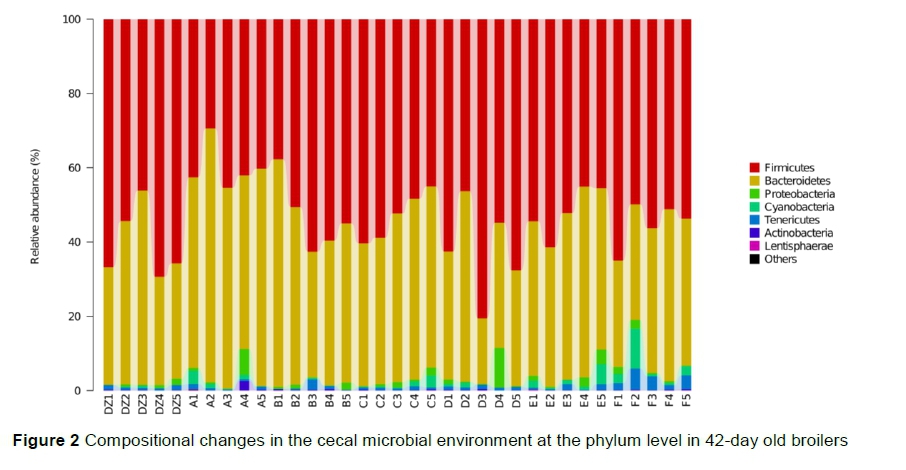
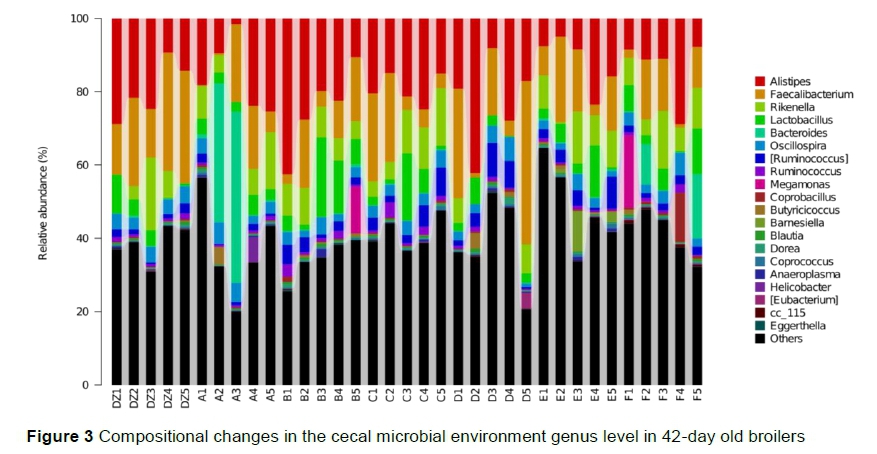
The dominant phyla across all groups were Firmicutes, Bacteroidetes, and Proteobacteria, which accounted for more than 90% of the total phyla in the cecum (Figure 2). The proportions of Cyanobacteria and Proteobacteria in Group F and Actinobacteria in Group A were higher than those in the other groups. The most abundant genera were Aíistipes, Faecaíibacterium, Rikeneíía, Lactobaciííus, Bacteroides, Oscillospira, Ruminococcus, and unclassified Ruminococcus (Figure 3). The proportions of Bacteroides in Group A and Megamonas and Coprobaciííus in Group F were significantly higher than in the other groups. In the present study, the most predominant phyla in the cecum of broilers were Bacteroidetes, Firmicutes, Actinobacteria, and Proteobacteria, which was consistent with (Oakley et al., 2014; Wei et al., 2013) and inconsistent with previous studies (Liu et al., 2018). It was found that the proportions of Actinobacteria and Bacteroides increased significantly after supplementation with component A. Actinobacteria includes probiotic strains Collinsella and Bifidobacterium (Tremaroli et al., 2012; Liu et al., 2018), whereas Bacteroidetes breaks down polysaccharides to improve nutrient utilization, promote immunity, and maintain the microecological balance in the intestine (Bäckhed et al., 2004).
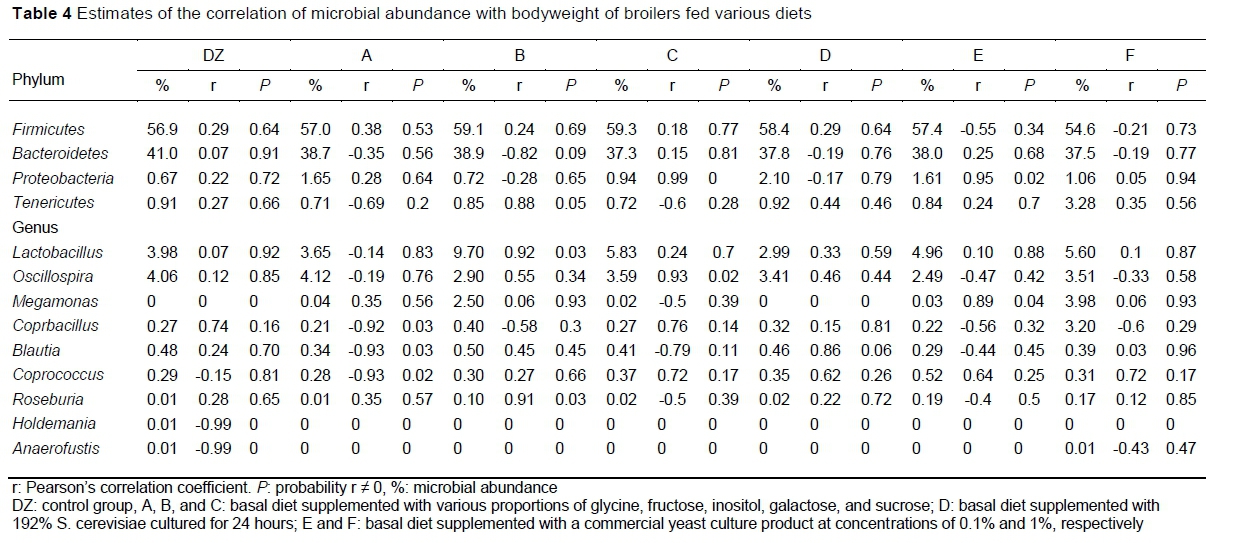
In this experiment, birds in Groups B, C and D were heavier at 42 days than those in DZ (Sun et al., 2020). The relationships of microbial abundance with bodyweight (r and P-values) are shown in Table 4. In Group B, Tenericutes was correlated positively with bodyweight. In Groups C and E, Proteobacteria was correlated positively with bodyweight. In B Group, Lactobacillus and Roseburia were correlated positively with bodyweight. In Group A, Coprobacillus, Blautia and Coprococcus were correlated negatively with bodyweight. In Group C, Oscillospira and in Group E, Megamonas were correlated positively with bodyweight. In the control group, Holdemania and Anaerofustis were correlated negatively with bodyweight.
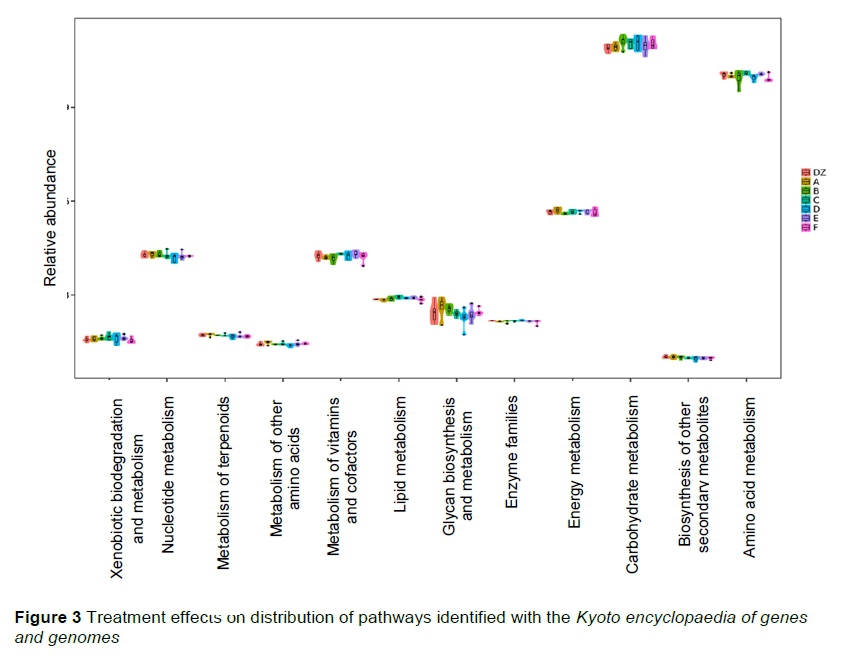
Kyoto encyclopaedia of genes and genomes (KEGG) analysis revealed that microbes in Group D were involved mainly in xenobiotic biodegradation, metabolism, and nucleotide metabolism (Figure 3). Microbes in Group B were involved mainly in nucleotide metabolism, cofactor and vitamin metabolism, lipid metabolism, and amino acid metabolism. Microbes in Groups A and C were involved mainly in energy metabolism and carbohydrate metabolism. In addition, the fermented YC with many active metabolites can affect the metabolic processes of intestinal microbes. Compared with the control, most metabolic processes, such as xenobiotic degradation, were higher in YC and active metabolite supplementation groups, consistent with the results of a previous study (Stanley et al., 2016). The authors observed differences in amino sugar and nucleotide sugar metabolism pathways in Groups A and C. Amino sugars break down proteins in feed into amino acids for energy (Miska et al., 2014), whereas nucleotide sugars are important for the synthesis of purines and pyrimidines for nucleotides (Rengaraj et al., 2013). As reported, the cecal microbial environment of birds with improved feed efficiency was enriched with microbes functioning in amino acid metabolism (Yan et al., 2017). Therefore, as active ingredients, amino acids and carbohydrates play important roles in the metabolism of nutrients in broilers.
Megamonas is a commensal organism present in the intestinal tract of mammals and birds (Hooper, 2004). A long-term study reported that the proportion of Lactobacillus was higher in healthy individuals than in unhealthy individuals (Isolauri et al., 1991). The proportion of Lactobacillus increased after YC supplementation, suggesting that it might promote overall health, which may have been because several Lactobacillus species can induce butyrate production. Roseburia and Oscillospira are also butyrate-producing bacteria, with Roseburia fermenting an array of carbohydrates. The proportions of both bacteria were shown to correlate positively with antioxidant activity and to correlate negatively with inflammation (Gophna et al., 2017; Wang et al., 2018). In this study, the proportions of Roseburia in Group B and Oscillospira in Group C were correlated positively with bodyweight. However, bacteria in the gut can produce harmful substances that affect nutrient absorption. For example, Holdemania can break down intestinal mucus and induce intestinal pain (Willems et al., 2015), whereas Anaerotruncus, an anaerobic bacterium, lacks metabolic enzymes (Lawson et al., 2015). Both bacteria were found to affect the production performance of broilers in the control group, but not in the YC group, indicating that YC can reduce the production of harmful bacteria and improve the overall health of the intestinal environment. Furthermore, Coprobacillus, which is widely distributed in mammalian intestines, was correlated negatively with bodyweight after supplementation with component A. Most Coprobacillus strains are non-pathogenic (Takayuki, 2015).
Conclusions
Dietary supplementation of YC could alter the proliferation of beneficial bacteria of broilers through its content of active metabolites. Further studies are needed to understand how these metabolites promote the growth performance of broilers.
Acknowledgements
This work was supported financially by the funding project 201805965017 granted by the China Scholarship Council and the Major Scientific & Technological Achievement Transformation Project of Jilin Province 20160301003NY.
Authors' Contributions
ZS, XF and GX were in charge of designing and conducting the experiment and analysing the data. All other authors participated in interpreting the results, and writing and editing the manuscript.
Conflict of Interest Declaration
The authors declare that there is no conflict of interest.
References
Adams, C.A., 2006. Nutrition-based health in animal production. J. Nutr. Res. Rev. 19, 79-89. DOI: 10.1079/NRR2005115 [ Links ]
Ahiwe, E., Abdallh, M., Chang'a, E., Al-Qahtani, M., Omede, A., Graham, H. & Iji, P., 2019. Influence of autolyzed whole yeast and yeast components on broiler chickens challenged with salmonella lipopolysaccharide. Poult. Sci. 98(12), 7129-7138. DOI: 10.3382/ps/pez452 [ Links ]
Bäckhed, F., Ding, H,, Wang, T., Hooper, L.V., Koh, G.Y., Nagy, A., Semenkovich, C.F. & Gordon, J.I., 2004. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. 101(44), 15718-15723. DOI: 10.1073/pnas.0407076101 [ Links ]
Bai, S.P., Wu, A.M., Ding, X.M., Lei Y., Bai, J., Zhang, K.Y. & Chio, J.S., 2013. Effects of probiotic-supplemented diets on growth performance and intestinal immune characteristics of broiler chickens. Poult. Sci. 92(3), 663-670. DOI: 10.3382/ps.2012-02813 [ Links ]
Bilal, R.M., Hassan, F., Saeed, M., Ayasan, T., Rashed, N., Akhtar, M.U. & Seidavi, A., 2020. Prospects of yeast based feed additives in poultry nutrition: Potential effects and applications. Indian J. Anim. Sci. 90(4), 495-505. http://www.journals4free.com/link.jsp?l=928357 [ Links ]
Chand, N., Shamsullah, Rafiullah, R.U. Khan, Mobashar, M., Naz, S., Rowghani, I. & Khan, M.A., 2019. Mannanoligosaccharide (MOS) in broiler ration during the starter phase: 1. Growth performance and intestinal histomorpholgy. Pak. J. Zool. 51, 173-176. [ Links ]
Corzo, A., Kidd, M.T., Burnham, D.J. & Kerr, B.J., 2004. Dietary glycine needs of broiler chicks. Poult. Sci. 83, 13821384. DOI: 10.1093/ps/83.8.1382 [ Links ]
DeSantis, T.Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E.L., Keller, K., Huber, T., Dalevi, D., Hu, P. & Andersen, G.L., 2006. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069-5072. DOI: 10.1128/AEM.03006-05 [ Links ]
Gophna, U., Konikoff, T. & Nielsen, H.B., 2017. Oscillospira and related bacteria - from metagenomic species to metabolic features. Environ. Microbiol. 19, 835-841. DOI: 10.1111/1462-2920.13658 [ Links ]
Han, G.G., 2018. Studies on the relationship of host physiological traits and metabolites with GutMicro biota in livestock. PhD (Agric) thesis, Seoul National University, South Korea. [ Links ]
Hooge, D.M., 2004. Turkey pen trials with dietary mannan oligosaccharide: Meta-analysis, 1993-2003. Int. J. Poult. Sci., 3, 179-188. DOI: 10.3923/ijps.2004.179.188 [ Links ]
Hooper, L.V., 2004. Bacterial contributions to mammalian gut development. Trends Microbiol. 12(3), 129-134. DOI: 10.1016/j.tim.2004.01.001 [ Links ]
Hussein, E. & Selim, S., 2018. Efficacy of yeast and multi-strain probiotic alone or in combination on growth performance, carcass traits, blood biochemical constituents, and meat quality of broiler chickens. Livest. Sci. 216, 153-159. DOI: 10.1016/j.livsci.2018.08.008 [ Links ]
Isolauri, E., Juntunen, M., Rautanen T., Sillanaukee, P. & Koivula, T.A., 1991. A human Lactobacillus strain (Lactobacillus casei sp strain GG) promotes recovery from acute diarrhea in children. Pediatrics 88(1), 90-97. DOI: 10.1203/00006450-199005000-00026 [ Links ]
Kondo, E., Marriott, P.J., Parker, R.M., Kouremenos, K.A., Morrison, P. & Adams, M., 2014. Metabolic profiling of yeast culture using gas chromatography coupled with orthogonal acceleration accurate mass time-of-flight mass spectrometry: Application to biomarker discovery. Anal. Chim. Acta. 807, 135-142. DOI: 10.1016/j.aca.2013.11.004 [ Links ]
Langille, M.G.I., Zaneveld, J., Caporaso, J.G., McDonald, D., Knights, D., Reyes, J.A., Clemente, J.C., Burkepile, D.E., Thurber, R.L.V. & Knight, R., 2013. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nature Biotechnology 31(9), 814-821. DOI: 10.1038/nbt.2676 [ Links ]
Lawson, P.A., 2015. Anaerofustis. In: Bergey's manual of systematics of archaea and bacteria. John Wiley & Sons, Ltd. DOI:10.1002/9781118960608.gbm00628 [ Links ]
Liu, H., Li, J.T., Guo, X.W., Liang Y.X. & Wang, W.M., 2018. Yeast culture dietary supplementation modulates gut microbiota, growth and biochemical parameters of grass carp. Microb. Biotechnol. 11(3), 551 -565. DOI: 10.1111/1751-7915.13261 [ Links ]
Miska, K.B., Fetterer, R.H. & Wong, E.A., 2014. The mRNA expression of amino acid transporters, aminopeptidase N, and the di- and tri-peptide transporter PepT1 in the embryo of the domesticated chicken (Gallus gallus) shows developmental regulation. Poult. Sci. 93, 2262-2270. DOI: 10.3382/ps.2014-03983 [ Links ]
Oakley, B.B., Lillehoj, H.S., Kogut, M.H., Kim, W.K., Maurer, J.J., Pedroso, A. & Cox, N.A., 2014. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 360, 100-112. DOI: 10.1111/1574-6968.12608 [ Links ]
Pirgozliev, V., Brearley, C.A., Rose, S.P. & Mansbridge, S.C., 2018. Manipulation of plasma myo-inositol in broiler chickens: effect on growth performance, dietary energy, nutrient availability, and hepatic function. Poul. Sci. 98(1), 260-268. DOI: 10.3382/ps/pey341 [ Links ]
Pirgozliev, V.R, Bedford, M.R., Rose, S.P., Whiting, I.M., Oluwatosin, O.O., Oso, A.O., Oke, F.O., Ivanova, S.G. & Staykova, G.P., 2017. Phosphorus utilisation and growth performance of broiler chicken fed diets containing graded levels of supplementary myo-inositol with and without exogenous phytase. J. World. Poul. Res. 7 (1), 1-7. https://repository.guildhe.ac.uk/id/eprint/15629/1/Vasil%20Pirgozliev%20phosphorou %20utilisation%2024%20Feb%2017%20upload.pdf [ Links ]
Poppy, G.D., Rabiee, A.R., Lean, I.J., Sanchez, W.K., Dorton, K.L. & Morley, P.S., 2012. A meta-analysis of the effects of feeding yeast culture produced by anaerobic fermentation of saccharomyces cerevisiae on milk production of lactating dairy cows. J. Dairy. Sci. 95(10), 6027-6041. DOI: 10.3168/jds.2012-5577 [ Links ]
Ran, C., Lu, H., Zhi, L., Xu, L., Yang, Y.L., Tacon, P., Auclair, E. & Zhou, Z.G., 2015. A comparison of the beneficial effects of live and heat-inactivated baker's yeast on Nile tilapia: Suggestions on the role and function of the secretory metabolites released from the yeast. PLoS ONE 10. DOI: 10.1371/journal.pone.0145448 [ Links ]
Rengaraj, D., Lee, B.R., Jang, H.J., Kim, Y.M. & Han, J.Y., 2013. Comparative metabolic pathway analysis with special reference to nucleotide metabolism-related genes in chicken primordial germ cells. Theriogenol. 79, 28-39. DOI: 10.1016/j.theriogenology.2012.09.004 [ Links ]
Saeed, M., Ayasan, T., Alagawany, M., Abdel-hack, M.E., Abdel-latif, M., Arain, M.A. & Chao, S., 2019. The role of ß- mannanase (Hemicell) in improving poultry productivity, health and environment. Braz. J. Poultry Sci. 21(3), 1-8. DOI: 10.1590/1806-9061-2019-1001 [ Links ]
Saied, J.M., Al-Jabary, Q.H. & Thalij, K.M., 2011. Effect of dietary supplement yeast culture on production and hematological parameters in broiler chicks. Int. J. Poult. Sci. 10, 376-380. DOI: 10.3923/ijps.2011.376.380 [ Links ]
Shen, Y.B., Piao, X.S., Kim, S.W., Wang, L., Liu, P., Yoon, I. & Zhen, Y.G., 2009. Effects of yeast culture supplementation on growth performance, intestinal health, and immune response of nursery pigs. J. Anim. Sci. 87, 2614-2624. DOI: 10.2527/jas.2008-1512 [ Links ]
Shurson, C.G., 2018. Yeast and yeast derivatives in feed additives and ingredients: Sources, characteristics, animal responses, and quantification methods. Anim. Feed. Sci. Tech. 235, 60-76. DOI: 10.1016/j.anifeedsci.2017.11.010 [ Links ]
Stanley, D., Hughes, R.J., Geier, M.S., & Moore, R.J., 2016. Bacteria within the gastrointestinal tract microbiota correlated with improved growth and feed conversion: Challenges presented for the identification of enhancing probiotic bacteria. Front. Microbiol. 7, 1-13. DOI: 10.1016/j.anifeedsci. 2017.11.010 [DOI for Shurson above] https://www.frontiersin.org/articles/10.3389/fmicb.2016.00187/full [ Links ]
Stanley, V.G., Gray, C., Daley, M., Krueger, W.F. & Sefton, A.E., 2004. An alternative to antibiotic-based drugs in feed for enhancing performance of broilers grown on Eimeria sp.-infected litter. Poult. Sci. 83, 39-44. DOI: 10.1093/ps/83.1.39 [ Links ]
Stone, C.W., 2006. Yeast products in the feed industry: A practical guide for feed professionals. https://en.engormix.com/feed-machinery/articles/yeast-products-infeed-industry-t33489.htm [ Links ]
Sun, Z., Wang, T., Demelash, N., Zheng, S., Zhao, W., Chen, X., Zhen, Y.G. & Qin, G.X., 2020. Effect of yeast culture (Saccharomyces cerevisiae) on broilers: A preliminary study on the effective components of yeast culture. Animals 10, 68, DOI: 10.3390/ani10010068 [ Links ]
Tremaroli, V. & Backhed, F., 2012. Functional interactions between the gut microbiota and host metabolism. Nature 489(7415), 242-249. DOI: 10.1038/nature11552 [ Links ]
Tufail, M., Chand, N., Rafiullah, Ahmad, S., R.U. Khan, Mobashar, M. & Naz. S., 2019. Mannanoligosaccharide (MOS) in broiler diet during the finisher phase. 2. Growth traits and intestinal histomorpholgy. Pak. J. Zool. 51, 597-602. [ Links ]
Wang, A., 2014. The effects of different feeding program and inclusion of glycerol, glucose or sucrose in broiler starter diets on growth performance and intestinal development. Master's thesis, Dalhousie University, Canada. https://central.bac-lac.gc.ca/.item?id=TC-NSHD-49110&op=pdf&app=Library&oclc_number=1032956679 [ Links ]
Wang, Y.B., Xie, Q.H., Sun, S., Huang, B.J., Zhang, Y., Xu, Y., Zhang, S.M. & Xiang, H.Y., 2018. Probiotics-fermented Massa Medicata Fermentata ameliorates weaning stress in piglets related to improving intestinal homeostasis. Appl. Microbiol. Biotechnol. 102, 10713-10727. DOI: 10.1007/s00253-018-9438-y [ Links ]
Wei, S., Morrison, M., & Yu, Z., 2013. Bacterial census of poultry intestinal microbiome. Poul. Sci. 92, 671-683. DOI: 10.3382/ps.2012-02822 [ Links ]
Whitman, W.B. (ed), 2015. Bergey's manual of systematics of archaea and bacteria. John Wiley & Sons, Hoboken, NJ, USA. DOI: 10.1002/9781118960608 [ Links ]
Willems, A., 2015. Holdemania. Bergey's manual of systematics of archaea and bacteria. John Wiley & Sons, Ltd. DOI: 10.1002/ 9781118960608.gbm00764 [ Links ]
Xu, Y., Yang, H., Zhang, L., Su, Y., Shi, D., Xiao, H. & Tian Y., 2016. High-throughput sequencing technology to reveal the composition and function of cecal microbiota in Dagu chicken. BMC Microbiol. 16(1), 259. DOI: 10.1186/s 12866-016-0877-2 [ Links ]
Yan, W., Sun, C., Yuan, J. & Yang, N., 2017. Gut metagenomic analysis reveals prominet roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 7, 45308. DOI: 10.1038/srep45308 [ Links ]
Zhang, A.W., Lee, B.D., Lee, S.K., Lee, K.W., An, G.H., Song, K.B. & Lee C.H., 2005. Effects of yeast (Saccharomyces cerevisiae) cell components on growth performance, meat quality, and ileal mucosa development of broiler chicks. Poult. Sci. 84, 1015-1021. DOI: 10.1093/ps/84.7.1015 [ Links ]
Submitted 19 June 2020
Accepted 10 March 2021
Published 12 November 2021
# Corresponding authors: zhangxuefengjlau@163.com; qgx@jlau.edu.cn


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}