










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 n.5 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i5.1
ARTICLES
Moringa oleofera leaf meal as a feed supplement for dairy calves
T.W. KekanaI, II, #; U. MarumeI; C.M. MuyaIII; F.V. Nherera-ChokudaIV
IDepartment of Animal Science, North-West University, Mmabatho, 2735, South Africa
IIElsenburg Agricultural Training College, Elsenburg, 7607, South Africa
IIIAgricultural Research Council-Animal Production Institute, Irene, 0062, South Africa
IVNational Emergent Red Meat Producers' Organisation, Pretoria, 0102, South Africa
ABSTRACT
The study evaluated the effects of supplements of Moringa oleífera leaf meal (MOLM) on blood biochemistry and growth of pre-weaned Holstein calves (n = 48). Four-day-old calves were blocked (based on order of birth and birth weight) and randomly allocated to three dietary treatments, namely M0: unsupplemented control, M30: supplemented with 8.33 g MOLM /100 kg bodyweight (BW), and M60: supplemented with 16.66 g MOLM /100 kg BW. The experiment ended when the calves were 42 days old. The MOLM was diluted in the milk of the morning feeding session. Feed intake, faecal score, and body temperature were recorded daily. Bodyweight and body measurements were taken weekly, and blood samples were collected at the same time. Starter dry matter intake (DMI) for the M60 group was higher than for M0 group with a significant week effect. Calves in M60 had higher final bodyweight and body measurements than M0 and M30. Similar trends were observed for superoxide dismutase (SOD), catalase (CAT), and serum antioxidant activity (SAOA). Immunoglobulin G (Ig) levels increased with the age of calves in both M30 and M60 groups, with the increase being progressively larger with the level of MOLM supplementation. Calves in the M60 group had fewer days of diarrhoea. Supplementation of MOLM supplied secondary metabolites and nutrients that improved feed utilization and later stimulated growth, increased enzymatic antioxidants, and reduced diarrhoeal incidence in pre-weaned calves. Dilution of MOLM by as little as 16.66 g/100 kg BW in the milk for dairy calves can be recommended.
Keywords: faecal score, immunoglobulin G, oxidative balance, weight gain
Introduction
Management of replacement heifers is a crucial component of dairy production (Hawkins et al., 2019). Neonatal calves do not produce enough gamma globulin, and the onset of breathing generates reactive oxygen species (ROS), which can result in oxidative stress (Wiedemann et al., 2003). Feeding high-quality colostrum is critical in conferring passive immunity to new-born calves (Besser & Gay, 1994; Godden, 2008; Stelwagen et al., 2009). However, colostrum has few antioxidants and is rich in pro-oxidants (Kankofer & Lipko-Przybylska, 2008; Sordillo, 2016) that induce oxidative stress (Albera & Kankofe, 2011). Failure to transfer passive immunity also contributes to oxidative stress and poor growth performance in early life (Ling et al., 2018). Severe oxidative stress is associated with diarrhoea and pneumonia in the first 21 days of life (Ranjan et al., 2006). Neonatal diarrhoea is as a major disease of calves and contributes to the risk of calf mortality (Windeyer et al., 2014). Interventions to counteract birth-associated oxidative stress by increasing their pool of antioxidants are critical for calf growth.
Several researchers (Khalel et al., 2014; Kholif et al., 2016; Falowo et al., 2018; Kekana et al., 2019, 2020) suggested synthetic anti-oxidants as alternatives to minimize the effects of ROS with different levels of success. From an organic perspective, Moringa oleifera has attracted attention because of the high concentrations of antioxidant, antimicrobial, and immunomodulatory properties inferred by flavonoids (Kholif et al., 2016; Dong et al., 2018; Falowo et al., 2018). Moreover, the leaves of MO are rich in minerals (Ca, K, Zn, Mg, Fe, and Cu) and vitamins (A, B, C, D, and E) (Mbikay, 2012) and are preferentially browsed by many species of birds, possibly because of these attributes (Nherera-Chokuda, 2021, personal observation). The potential of MO leaves as immunity boosters in mature cows and goats has been noted (Kekana et al., 2019; Kekana et al., 2020; Kholif et al., 2016). However, its effects on the maintenance of oxidative stress, energy balance, health, and growth performance in calves have not been fully investigated. Hence, this study evaluated the effects of pre-weaning supplementation of MOLM on immunity, diarrhoea control, and growth performance in dairy calves.
Material and methods
This experiment was conducted at the Agricultural Research Council - Animal Production Institute at Irene, South Africa (28013' 0" S; 25055' 0" E, altitude 1523 m) during the November 2017 to January 2018 calving season. Ethical clearance was granted by the Institute's ethical review committee (APIEC 17/008).
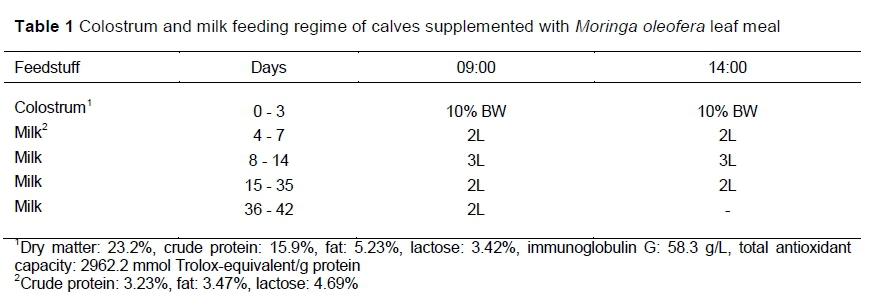
After calving, cows were monitored closely, and suckling was prevented. After cleaning, calves were separated from the dams and taken to the calf unit where birth weight was determined. Colostrum samples were collected and tested with a colostrometer (Biogenics, Napa, Calif, USA) for quality control and fed to calves within six hours of birth. Then the calves were fed colostrum-rich milk twice daily (09h00 and 14h00) at the rate of 10% of their birth BW until day 3. The calves were housed individually in 15 m2 pens consisting of concrete and dirt ground (concrete floor with a rubber mat and Eragrostis curvula hay for cold shock. The concrete floor (8 m2) had a roof, and the remaining area was dirt exercise. Pens were cleaned every other day with a pressured hose and disinfectant. Only 18 of 30 pens were used twice, so were cleaned and disinfected (Formalin, ANB Vet, Midrand) and left dry for seven days before housing the new calf. Throughout the trial, health was observed daily.
Forty-eight new-born female Holstein calves (32.5 ± 4.5 kg BW) were used in this study. The calves were randomly allocated at birth to one of three treatment groups. Month of birth was considered a block, and treatments were allocated within the block. Treatments were based on levels of MOLM, namely M0: control, M30: 8.33 g MOLM/100 kg BW and M60: 16.66 g MOLM/100 kg BW. A portion of milk (100 ml) from the calf's daily allowance was warmed to 40 °C and mixed with the amounts of MOLM for each treatment. The mixture was returned to the milk feeding bucket and fed to each calf from day 4 to 42 days old. The composition of MOLM was as described in Kekana et al. (2020) and the levels were adjusted weekly to maintain 8.33/100 kg and 16.66/100 kg BW.
The milk feeding regime is shown in Table 1. Between day 4 and day 42, calves received starter feed (Table 2) and fresh clean water at libitum. Calf starter, milk and water intakes were recorded daily as the difference between amounts offered every morning and refusals, collected the following morning before the next feeding.
Composite daily milk samples from the dairy were collected weekly in the morning and evening and pooled to determine their CP, fat and lactose content using a System 4000 infrared analyser (Foss Electric, Hiller0d, Denmark). Calf starter was sampled, and oven dried at 90 °C for 24 hours to determine its DM content. Ground samples were analysed for CP according to method 968.06, and ether extract (EE) according to method 920.39 (AOAC, 2000). Calcium, potassium, and phosphorus were assayed according to method 965.17(AOAC, 2000). Neutral detergent fibre was determined following Van Soest et al. (1991).
The calves were weighed at birth and at seven-day intervals until weaned at 42 days old. Bodyweight was measured using a digital weighing scale (TAL-TEC®, Brits, South Africa) and bodyweight change was calculated. Heart girth was defined as the circumference of the thoracic cavity immediately behind the forelimbs, and body length was the distance from point of the shoulder to the point of tuber ischia. All measurements were taken in the morning before feeding.
Blood samples were collected before feeding and treatment allocation on day 3, and before feeding on days 12, 22, 32, and 42. Samples were collected from the jugular vein using a 10-ml vacutainer tube (ANB Vet, Centurion, South Africa). Then the samples were kept at room temperature for a minimum of 1 hour to allow clotting to occur before centrifugation at 1764 χ g for 15 minutes. Serum was decanted into three aliquots and stored at -20 °C until analysis.
Beta-hydroxybutyrate (ßHB) (Williamson et al., 1962), non-esterified fatty acids (NEFA) (De Villiers et al., 1977) and glucose (Bradford, 1976) were analysed. The activities of superoxide dismutase (SOD) (Beauchamp & Fridovich, 1971), glutathione peroxidase activity (GSH-Px) (Paglia & Valentine, 1967), catalase activity (CAT) and malondialdehyde (MDA) (Tug et al., 2005) were also assayed, and serum antioxidant activity (SAOA) was assayed according to Koracevic et al. (2001).
Immunoglobulin G (IgG) concentration was analysed with an ELISA kit for bovine IgG from Bio-X Diagnostics (Jemelle, Belgium). All kit components were brought to 21 °C before use. The wash buffer was diluted twentyfold with distilled water. A calibration curve was developed and samples were diluted at 1/1000 in PBS according to the manufacturer's instructions. Diluted samples were added to the test plate in duplicate and incubated at 21 °C for 1 hour. The test plate was then washed three times with the wash buffer before a chromogen solution (100 μΙ_) was added to each well and incubated away from light for approximately 10 minutes. Finally, stop solution (50 μΙ_) was added to each well and the optical densities were recorded with a microplate spectrophotometer with a 450-nm filter. An interassay CV of <0.15 was observed. The concentration of IgG in samples was calculated from the standard reference curve provided in the test kit.
Body temperature was measured daily between 8h00 and 9h00 with a rectal thermometer (Dt-K111B, ANB Vet, Midrand, South Africa). The procedure was repeated twice for accuracy. All occurrences and treatments of lung diseases and diarrhoea were noted individually. Lung infection was defined as coughing or sneezing for two days or more or heavy breathing with additional signs such as nasal discharge. Diarrhoea was defined as soft watery faeces lasting for two days or more, or as soft or watery faeces in combination with impaired condition and possible weight loss.
Faecal scoring was conducted daily (08h00) to determine faecal fluidity and consistency on a scale of 1-4, in which 1 = formed stools, 2 = soft or moderate consistency, 3 = start of the diarrhoea (mild) or runny, and 4 = watery faeces or profuse diarrhoea (Larson et al., 1977). Number of days of diarrhea (duration), diarrheal occurrence (frequency) and severity (faecal score) were recorded.
The MIXED procedure of SAS version 9.0 was used to analyse the data (SAS Institute Inc., Cary, North Carolina, USA). The linear model included fixed effects of block, treatment, time, and two- and three-way interactions, with the identity of the calf included as a random effect. No block, block χ treatment, block χ time, and block χ treatment χ time effects were identified. Time was used as a repeated measure with a first-order autoregressive covariance structure, which provided the best fit according to Schwartz's Bayesian information criterion. Degrees of freedom were calculated using the Kenward-Roger option, and the means were determined with the least-squares means statement. Effects were considered significant at P <0.05 and existence of a trend was accepted when 0.05 < P <0.10.
Results and Discussion
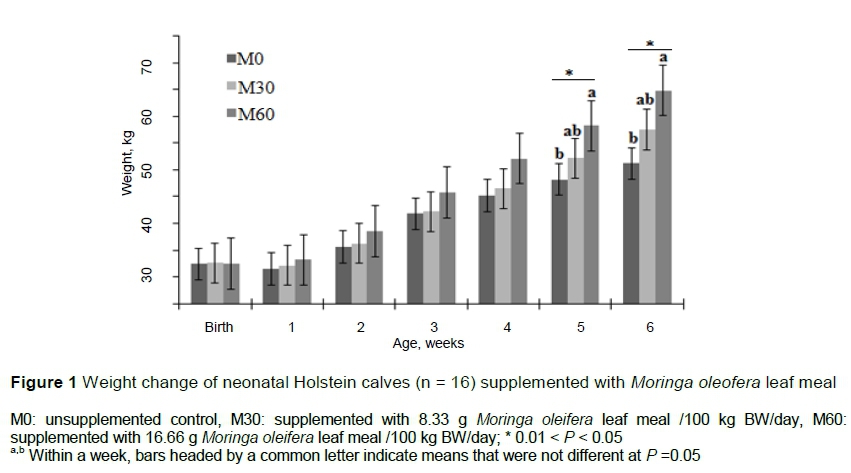
Intake of solid and liquid feeds increased with the level of MOLM. Both levels of MOLM increased starter DMI compared with the control group (P =0.01). Mean milk and water intakes for the M60 group were higher than M0 (p =0.01). Feed and water intake was 28% and 14% higher in M60 compared with M0 and M30, respectively. However, there were no differences between M60 and M30. A similar trend was noted on final bodyweight (P =0.04). Average daily gain tended to be higher in M60 (0.48 kg/day) (P =0.06) relative to control (0.39 kg/ day). When evaluated weekly, weight in the M60 group increased (P =0.02) and was higher in week 4 (Figure 1). The body measurements of heart girth and body length increased significantly (P <0.01) with MOLM levels. Means for feed intake and growth performance are presented in Table 3.
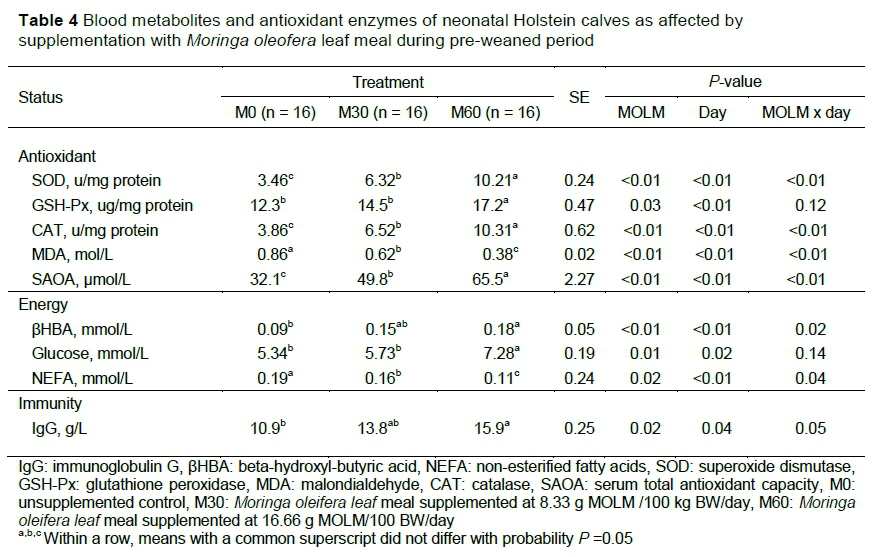
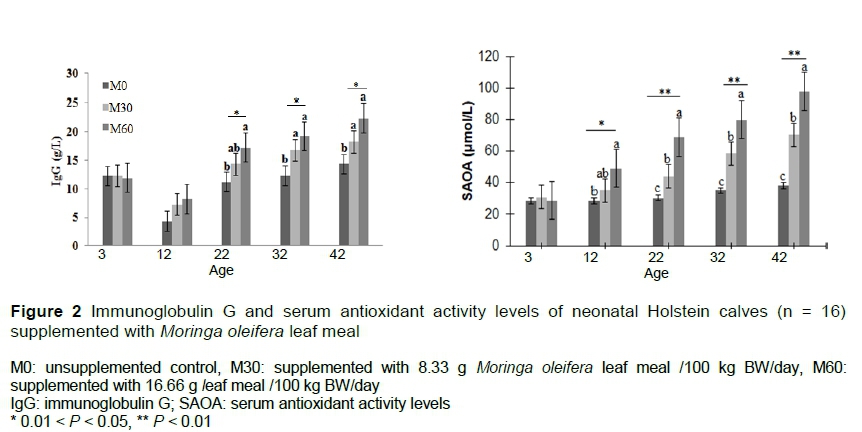
Mean antioxidant status (SOD, CAT, GSH-Px, and SAOA), energy metabolites (ßHBA, glucose and NEFA) and immunity (IgG) are presented in Table 4. With the exception of MDA, the activities of oxidative enzymes increased with the level of MOLM. The activity of SOD from M60 was the highest (P <0.01) relative to M0 and M30 calves, with M0 being the least. A similar trend was observed with M60 having the highest CAT (P <0.001) and SAOA (P <0.01). SAOA increased from 12 days old (P <0.01) until the end of the trial (Figure 2). The concentration of MDA was reduced by the inclusion of MOLM (P <0.001). The activity of GSH-Px also increased with the incremental levels of MOLM (P =0.03), with more significant effect over time (week).
Calves fed M60 had higher (P <0.01) ßHBA than M0, but M30 did not differ from the M0 group. Similarly, glucose levels did not vary between M0 and M30, but both groups had lower concentrations (P =0.01) than the M60 calves. Non-esterified fatty acid concentrations were lowest (P <0.01) in M60 (0.11 mmol/L) compared with M30 (0.17 mmol/L) and M0 (0.19 mmol/L) with a significant week effect (P <0.01), and treatment-week interaction (P =0.04) effect. The IgG levels were higher in M60 calves (P =0.02) than M0, but M60 and M30 did not differ (P >0.05). When evaluated over time, IgG for the MOLM groups increased with age (P =0.04) with a significant increase being noted from day 22 for M60 (Figure 2).
The M60 calves had the fewest days of diarrhoea (P <0.01) and lowest diarrhoea frequency (P =0.04), with a noticeable week effect (P <0.01, P = 0.01) (Table 5). Moringa oleífera leaf meal tended (P =0.08) to reduce the faecal score. Calves in the M0 group received electrolytes for more days (4 days) compared with the M60 group (2 days), and only 3 days in M30. The effects of MOML were not observed on lung disease and body temperature.
Moringa oleifera and its by-products have been investigated extensively in ruminant nutrition because of its relatively high biomass yield, high nutritional quality, and low cost of production. Hence, in this study, antioxidant enzymes, energy metabolites, health, and growth performance were investigated when pre-weaned Holstein calves were fed increasing levels of MOLM.
The current data showed that milk intake rose with the increasing levels of MOLM, though an unexpected reduction in milk intake was noted from control calves, which could have been from loss of appetite as a result of sickness (diarrhoea) (Heath et al., 1987). The increased milk in MOLM-supplemented calves could be attributed to the changed taste and the aroma of the milk, which stimulated appetite. In this study, the dilution of MOLM in milk may have acted as a sialagogue and stimulated the secretion of saliva, which made swallowing easier. Herbal supplements enhance and add flavours to animal feed, and therefore influence eating patterns, secretion of digestive fluids and total feed intake (Bhatt, 2015).
Milk and water intakes were stimulated in MOLM groups, and also intake of starter feed. Stimulated feed intake in neonatal calves is associated with higher nutrient digestibility, promoted reticular rumen weight, papillae width and papillae density as a result of elevated ruminal butyric acid (Muya et al., 2017). In Kholif et al. (2016), a 25% increase in ruminal butyric acid concurrent with enhanced nutrient digestibility was observed in Nubian goats that were supplemented with MOLM. It could therefore be inferred that MOLM supplementation induced ruminal butyric acid production, which accelerated rumen development and stimulated nutrient digestibility in calves, although ruminal butyric acid was not measured in the current study. This resulted in increased feed intake providing a greater quantity of energy that was reflected in increased energy metabolites (glucose and ßHBA). Higher serum glucose level is essential in calves as it serves as the primary source of energy before rumen development. However, as consumption of solid feed increases (beyond 21 days) and the rumen begins to develop, the contribution of volatile fatty acids (VFAs) to the calves' energy increases, resulting in a higher level of ßHBA (Muya et al., 2015). This could explain the elevated ßHBA levels in calves treated with MOLM.
There is a dearth of literature about the effects of flavonoids and phenolic-rich herbs on the performance of pre-weaned dairy calves. However, Frutos et al. (2004) reported that rumen microorganisms in calves at 21 days old could tolerate low and moderate concentrations of secondary metabolites, including phenolic compounds and tannins, without negative effects on rumen fermentation. Phenolic compounds and tannins inhibit ruminal methanogenesis in the rumen environment of the matured ruminants, resulting in efficient repartition of the energy consumed in methane and milk production (Aerts et al., 1999). Supplementation of MOLM to mature goats resulted in increased in VFA and propionate concentrations in the rumen (Kholif et al., 2014), the key substrates for gluconeogenesis. Collectively, these characteristics explain the greater performance in this study (Figure 1). Elaidy et al. (2019) observed increased weight gain and feed/gain ratio from MOLM supplementation in buffalo calves. Ahmed and Shaarawy (2019) reported a significant growth performance in final weight gain, total weight gain, average daily gain and feed conversion efficiency in goat kids that received Moringa leaves.
The current data showed that MOLM could reduce the blood NEFAs and the activity of MDA. In calves, NEFA levels are established maternally through colostrum feeding (Ling at aí., 2018) and may directly induce cellular oxidative stress (Inoguchi et al., 2000). The high levels of NEFA alter gene expression that is related to lipid and carbohydrate metabolism in bovine embryos (Van Hoeck et al., 2015). Malondialdehyde is another degradation product of lipid peroxidation formed under oxidative stress conditions (Nielsen et al. 1997). Hence, the reduced levels of malondialdehyde reflect i) reduced risk of oxidative stress, ii) reduced lipid peroxidation and iii) pronounced energy balance in dairy cattle (Caré et al., 2018; Ling et al., 2018). In this study, NEFA levels were much lower than those reported by Ling et al. (2018). Similarly, reduced NEFA levels after MOLM supplementation were observed in mature cows (Kekana et al., 2019; Shankhpal et al., 2019; Kekana et al., 2020), mature goats (Kholif et al., 2016; Ahmed & Shaarawy, 2019), attributable to the increased feed intake that provided greater quantities of energy. Similarly, Kirisattayakul et al. (2013) reported significantly attenuated MDA levels with M. oíeifera leaf extract at a dose of 100 mg.kg-1 BW in rats. In Serafini et al. (2011), M. citrifoíia rich with flavonoids acted against lipid peroxidation and hydroxyl and nitric-oxide-induced radicals and improved antioxidant activity. Although low NEFA levels and reduced MDA activity are desirable, they also play a role in curbing production of cellular free radicals (ROS) indirectly by stimulating inflammatory pathways under severe lipid peroxidation (Gawel et al., 2004; Ohtsu et al., 2017).
The authors evaluated the activities of SOD, CAT, and GSH-Px as proxy measures for oxidative damage to further determine oxidative stress in calves. During the enzymatic regulation of ROS, SOD is the first enzyme that is involved in the conversion of oxygen radicals to hydrogen peroxide (H2O2), whereas CAT is involved in the second step of removing H2O2 and converting it to O2 (Yu, 1994). Also, GSH-Px acts directly on H2O2 reduction during free radical mutation (Dröge, 2002). Currently, the activities of SOD, CAT and GSH-Px in serum increased with the level of MOLM and could be attributed to the synergistic antioxidant activities from antioxidant minerals and vitamins (Mbikay, 2012), flavonoids and phenolic (Falowo et al., 2018) in MOLM. Flavonoids (quercetin, kaempferol) were reported to possess abilities to reduce the activity of peroxidase and inhibit the release of free radicals by stimulating the proliferation of neutrophils and the activation of these cells through elevated a1-antitrypsin in vitro (Nijveldt et al., 2001). Moreover, the interaction of MOLM polyphenols with nitric oxide synthases may have inhibited xanthine oxidase (XO), a key source of free radicals. For example, quercetin, silybin and luteolin inhibit the activity of XO, thereby inducing serum activities of antioxidant enzymes (SOD, CAT, and GSH-Px) (Nijveldt et al., 2001). This resulted in balanced redox homeostasis as reflected by the increase in SAOA (Figure 2). Given that SAOA is the determinant of ROS scavenging capability and changes in the redox balance of the living cell (Maciejczyk et al., 2018), the current results highlighted the potentiality of MOLM in curbing the detrimental effects of maternal oxidative stress in neonates during the pre-weaned period. In addition, higher anti-oxidative capacity augments the foundation for sturdy immunity and prevention of subsequent diseases in calves (Nijveldt et al., 2001; Forman, 2016; Sordillo, 2016).
In calves, a solid immune defence that can rapidly inactivate pathogenic invasions is established by adequate serum immunoglobulins (Ig). Therefore, calves with adequate serum IgG will continue to grow normally and not be deterred, as would calves with insufficient IgG (Rodewald, 1976). The current data showed that serum IgG levels increased (Figure 2) with age in all calves. Similarly, Mandal et al. (2015) reported that IgG levels increased with sun-dried MOLM supplementation in cattle calves that were exposed to the industrial fluorotic area. In contrast, Elaidy et al. (2017) noted low IgG levels, but a significant rise in other immune parameters (leucocytes, plasma fluoride and haematology) of buffalo calves following oven-dried MOLM supplementation. The discrepancy could be because of the differing levels of supplementation, stage of production and breed. In addition, processing methods (extract, oven, sun, and shade dried) affected the levels of the available antioxidants such as ascorbic acids, polyphenols, flavonoids, and organosulfur compounds in MOLM (Aja et al., 2014; Kholif et al., 2016; Falowo et al., 2018), which could have contributed to different physiological responses after supplementation.
The stool fluid content reflects a balance between luminal input (ingestion and secretion of water and electrolytes) and output (absorption) along the gastrointestinal tract (Mukherjee et al., 2011). In the current study, MOLM calves tended to have a lower faecal score, resulting in a significantly reduced number of days that calves were fed electrolytes. Moreover, the number of diarrhoea days and diarrhoea frequency were significantly reduced in MOLM-fed groups, whereas all calves had the same prevalence of lung disease. This could not be explained so the secondary metabolites responsible for anti-diarrheal activity in MOLM, particularly in ruminants, have yet to be identified. However, the flavonoids in MOLM were reported to exhibit anti-diarrheal activity through inhibition of the intestinal motility and hydro electrolytic secretion in rats (Mukherjee et al., 2011). Moringa oleífera flavonoids were also responsible for inhibited hypersecretion and gastrointestinal motility in human beings (Lakshminarayana et al., 2011). Furthermore, the antimicrobial properties in the gut of goats and dairy cows were attributed to the moderate concentrations of phenolic and tannins in Moringa oleífera leaf (Kholif et al., 2016; Dong et al., 2018). Reports cited in this study provide evidence of the antidiarrheal activities of flavonoids and phenolic in MOLM and support the authors' observations that Moringa oíeifera reduced diarrhoeal incidence in supplemented calves.
Conclusion
Moringa oíeifera leaf meal supplementation as low as 0.02% BW supplied secondary metabolites and nutrients that improved feed utilization, enzymatic antioxidant status and calf health, reduced oxidative stress, and subsequently stimulated growth. Further studies are needed to determine the optimal intake of MOLM and study its effects on rumen development.
Acknowledgements
This study was made possible by a grant from Professional Development Programme (PDP) at ARC-AP and Department of Rural Development and Land Reform (DRDLR) of South Africa. The authors also thank North-West University and University of Pretoria, Onderstepoort Campus, for their input.
Authors' Contributions
TWK, FVN-C and UM carried out conceptualization, methodology, formal and statistical analysis, investigation, data curation, writing-original draft, editing and visualization, validation, resources, writing review and editing. CMM was involved in project administration, funding acquisition and validation.
Conflict of Interest Declaration
The authors declare there is no conflict of interest and confirm that the manuscript has been read and approved by all authors.
References
Aerts, R.J., Barry, T.N., McNabb, W.C. Polyphenols and agriculture: Beneficial effects of proanthocyanidins in forages. Agric. Ecosyst. Environ. 1999, 75, 1-12. https://org/10.1016/S0167-8809(99)00062-6 [ Links ]
Ahmed, M.E. & Shaarawy, A.M., 2019. Effect of feeding Moringa oleífera forage on productive performance of growing goat kids. Egypt. J. Sheep Goats Sci. 14, 25-37. https://easg-org.com/wp-content/uploads/2019/05/11-318-141-9-Motazz-new2-column.pdf [ Links ]
Aja, P.M., Nwachukwu, N., Ibiam, A.U., Igwenyi, I.O. & Onu, P.N., 2014. Comparative evaluation of transaminases and alkaline phosphatase activities in Albino rats administered aqueous, ethanolic and methanolic extracts of Moringa oleífera seeds locally grown in Abakaliki, Nigeria. J. Biol. Chem. Res., 31,164-181. [ Links ]
Albera, E. & Kankofer, M., 2011. The comparison of antioxidative/oxidative profile in blood, colostrum and milk of early post-partum cows and their newborns. Reprod. Domest. Anim. 46, 763-769. DOI: 10.1111/j.1439-0531.2010.01737x [ Links ]
AOAC, 2000. Official method of analysis. 17th edition. Volume I. Association of Official Analytical Chemists, Inc., Maryland, USA. [ Links ]
AOAC, 2005. Official method of analysis. 18th edition. Association of Officiating Analytical Chemists, Washington DC. [ Links ]
Beauchamp, C. & Fridovich, I., 1971. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 44, 276-287. https://doi.org/10.1016/0003-2697(71)90370-8 [ Links ]
Besser, T.E. & Gay, C.C., 1994. The importance of colostrum to the health of the neonatal calf. Vet. Clin. N. Am. Food Anim. Pract. 10: 107-117. DOI: 10.1016/S0749-0720(15)30591-0 [ Links ]
Bradford, M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein using the principle of protein dye binding. Anal. Biochem. 72, 248-254. http://doi.org/10.1016/0003-2697(76)90527-3 [ Links ]
Bueno, M.P., 1997. Collaborative study: determination of retinol and carotene by high-performance liquid chromatography. Food Chem. 59, 165-170. https://doi.org/10.1016/S0308-8146(95)00227-8 [ Links ]
S. Caré, S., Trevisi, E., Minuti, A., Ferrari, A., Loor, J.J. & Calamari, L., 2018. Plasma fructosamine during the transition period and its relationship with energy metabolism and inflammation biomarkers in dairy cows. Livestock Sci. 216, 138-147. https://doi.org/10.1016/j.livsci.2018.08.003 [ Links ]
De Villiers, S., Van Der Walt, J.G. & Procos, J., 1977. An accurate, sensitive and reproducible method for the colorimetric estimation of free fatty acids in plasma. Onderstepoort J. Vet. Res. 44, 169-172. [ Links ]
Dong, L., Zhang, T. & Diao, Q., 2019. Effect of dietary supplementation of Moringa oleífera on the production performance and faecal methanogenic community of lactating dairy cows. Anim. 9, 262. DOI: 103390/ani9050262 [ Links ]
Dröge, W., 2002. Free radicals in the physiological control of cell function. Physiol. Rev. 82. 47-95. DOI: 10.1152/physrev.00018.2001 [ Links ]
Dunn, A., Ashfield, A., Earley, B., Welsh, M., Gordon, A., McGee, M. & Morrison, S.J., 2017. Effect of concentrate supplementation during the dry period on colostrum quality and effect of colostrum feeding regimen on passive transfer of immunity, calf health, and performance. J. Dairy Sci. 100, 357-370. https://doi.org/10.3168/jds.2016-11334 [ Links ]
El Shanawany, E.E., Nassar, W.E., & Ata, E.B., 2019. Detection of humoral and cellular immune responses in buffaloes naturally infected with sarcocystosis with risk factor assessment. Acta Veterinaria-Beograd 69, 275-289. https://doi.org/10.2478/acve-2019-0023 [ Links ]
Elaidy, A.A., Selim, I.A.A., Abou-Elenin, E.I., Abbas, M.S. & Sobhy, H.M., 2017. Effect of feeding dry Moringa oleífera leaves on the performance of suckling buffalo calves. Asian J. Anim. Sci. 11, 32-39. DOI:10.3923/ajas.2017.32.39 [ Links ]
Falowo, A.B., Mukumbo, F.E., Idamokoro, E.M., Lorenzo, J.M., Afolayan, A.J. & Muchenje, V., 2018. Multi-functional application of Moringa oleífera Lam. in nutrition and animal food products. Food Res. Int. 106, 317-334. DOI:10.1016/j.foodres.2017.12.079 [ Links ]
Forman, H.J., 2016. Redox signalling: An evolution from free radicals to aging. Free Radical Bio. Med. 97, 398-407. DOI: 10.1016/j.freeradbiomed.2016.07.003 [ Links ]
Frutos, P., Hervás, G., Giráldez, F.J. & Mantecón, Á.R., 2004. Tannins and ruminant nutrition. Span. J. Agric. Res. 2, 191-202. http://hdl.handle.net/10261/17401 [ Links ]
GaweJ, S., Wardas, M., Niedworok, E., Wardas, P., 2004. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiadomoscilekarskie 57, 453-455. [ Links ]
Godden, S., 2008. Colostrum management for dairy calves. Vet. Clin. North Am. Food Anim. Pract. 24, 19-39. DOI:10.1016/j.cvfa.2007.10.005 [ Links ]
Hawkins, A., Burdine, K., Amaral-Phillips, D. & Costa, J.H.C., 2019. An economic analysis of the costs associated with pre-weaning management strategies for dairy heifers. Anim. 9. 471. DOI: 10.3390/ani9070471 [ Links ]
Heath, S.E., Naylor, J.M., Guedo, B.L., Petrie, L., Rousseaux, C.G. & Radostits, O.M., 1989. The effects of feeding milk to diarrheic calves supplemented with oral electrolytes. Can. J. Vet. Res. 53, 477-485. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1255580/ [ Links ]
Inoguchi, T., Li, P., Umeda, F., Yu, H.Y., Kakimoto, M., Imamura, M., Aoki, T., Etoh, T., Hashimoto, T., Naruse, M. & Sano, H., 2000. High glucose level and free fatty acid stimulate reactive oxygen species production throughprotein kinase C-dependent activation of NAD(P)H oxidase in cultured vascular cells. Diabetes 49, 1939-1945.https://doi.org/10.2337/diabetes.49.11.1939 [ Links ]
Kankofer M., Lipko-Przybylska J., 2008. Physiological antioxidative/oxidative status in bovine colostrum and mature milk. Acta Vet. 58, 231-239.DOI: 10.2298/AVB0803231K [ Links ]
Kekana, T.W., Marume, U., Muya, C.M. & Nherera-Chokuda, F.V., 2019. Lactation performance and blood metabolites in lactating dairy cows micro-supplemented with Moringa oleifera leaf meal. S. Afr. J. Anim. Sci. 49, 709-716. http://dx.doi.org/10.4314/sajas.v49i4.12 [ Links ]
Kekana, T.W., Marume, U., Muya, M.C. & Nherera-Chokuda, F.V., 2020. Periparturient antioxidant enzymes, haematological profile and milk production of dairy cows supplemented with Moringa oleifera leaf meal. Anim. Feed Sci. Technol. 268, 114606. https://doi.org/10.1016/j.anifeedsci.2020.114606 [ Links ]
Kholif, A.E., Morsy, T.A., Gouda, G.A., Anele, U.Y. & Galyean, M.L., 2016. Effect of feeding diets with processed Moringa oleifera meal as protein source in lactating Anglo-Nubian goats. Anim. Feed Sci. Technol. 217, 45-55. http://dx.doi.org/10.1016/j.anifeedsci.2016.04.012 [ Links ]
Kirisattayakul, W., Wattanathorn, J., Tong-Un, T., Muchimapura, S., Wannanon, P. & Jittiwat, J., 2013. Cerebroprotective effect of Moringa oleifera against focal ischemic stroke induced by middle cerebral artery occlusion. Oxi. Med. Cell. Long. DOI: 10.1155/2013/95145 [ Links ]
Koracevic, D., Koracevic, G., Djordjevic, V., Andrejevic, S. & Cosic, V., 2001. Method for the measurement of antioxidant activity in human fluids. J. Clin. Path. 54, 356-361. DOI: 10.1136/jcp.54.5.356 [ Links ]
Lakshminarayana, M., Shivkumar, H., Rimaben, P. & Bhargava, V.K., 2011. Antidiarrhoeal activity of leaf extract of Moringa oleifera in experimentally induced diarrhoea in rats. Int. J. Phytomedic. 3, 68. https://www.arjournals.org/index.php/ijpm/article/view/274 [ Links ]
Larson, L.L., Owen, E.G., Albright, J.L., Appleman, R.D., Lamb, R.C. & Muller, L.D., 1977. Guidelines towards more uniformity in measuring and reporting calf experimental data. J. Dairy Sci. 60, 989-991. https://doi.org/10.3168/jds.S0022-0302(77)83975-1 [ Links ]
Ling, T., Hernandez-Jover, M., Sordillo, L.M. & Abuelo, A., 2014. Maternal late-gestation metabolic stress is associated with changes in immune and metabolic responses of dairy calves. J. Dairy Sci. 101, 6568-6580. https://doi.org/10.3168/jds.2017-14038 [ Links ]
Maciejczyk, M., Zebrowska, E., Zalewska, A. & Chabowski, A., 2019. Redox balance, antioxidant defense, and oxidative damage in the hypothalamus and cerebral cortex of rats with high fat diet-induced insulin resistance. Oxid. Med. Cell. Longev. 6940515. https://doi.org/10.1155/2018/6940515 [ Links ]
Mandal, K.D., Das, M.R., Pati, M., Pati, P.D., Gupta, A.R., Patra, R.C. & Senapati, S.K., 2015. Effect of Moringa oleífera on haematological parameters of calves reared in industrial fluorotic area. Vet. World 8, 1364. DOI: 10.14202/vetworld.2015.1364-1369 [ Links ]
Mbikay, M., 2012. Therapeutic potential of Moringa oleífera leaves in chronic hyperglycemia and dyslipidemia. Front. Pharmacol. 3, 24. https://doi.org/10.3389/fphar.2012.00024 [ Links ]
Mukherjee, P.K., Saha, K., Murugesan, T., Manda,l S.C., Pal, M. & Saha, B.P., 1998. Screening of anti-diarrhoeal profile of some plant extracts of a specific region of West Bengal, India. J. Ethnopharmacol. 199860, 85-9. DOI: 10.1016/s0378-8741(97)00130-x. [ Links ]
Muya, M.C., Erasmus, L.J., Miller, K., Aperce, C., Nherera, F.V. & Moshidi, P.M., 2017. Performance of Holstein calves having free access to milk and dosed with Megasphaeraelsdenii. Sci. Agr. 74, 189-194. https://doi.org/10.1590/1678-992x-2015-0318 https://www.scielo.br/j/sa/a/6QbvSDLFP65WBjqdWPtGcmJ/?lang=en&format=html [ Links ]
Muya, M.C., Nherera, F.V., Miller, K.A., Aperce, C.C., Moshidi, P.M. & Erasmus, L.J., 2015. Effect of Megasphaeraelsdenii NCIMB 41125 dosing on rumen development, volatile fatty acid production and blood β-hydroxybutyrate in neonatal dairy calves. J. Anim. Physiol. Anim. Nutr. 99, 913-918. https://doi.org/10.1111/jpn.12306 [ Links ]
Nielsen, F., Mikkelsen, B., Nielsen, J., Andersen, H. & Grandjean, P., 1997. Plasma malondialdehyde as biomarker for oxidative stress: Reference interval and effects of life-style factors. Clin. Chem. 43, 1209-1214. https://doi.org/10.1093/cNnchem/437.1209 [ Links ]
Nijveldt, R.J., Van Noo, E., Van Hoorn, D.E.C., Boelens, P.G., Van Norren K. & Van Leeuwen, P.A.M. 2001. Flavonoids: a review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 74(4), 418-425. https://doi.org/10.1093/ajcn/74.4.418 [ Links ]
Ohtsu, A., Tanaka, H., Seno, K., Iwata, H., Kuwayama, T. & Shirasuna, K., 2017. Palmitic acid stimulates interleukin-8 via the TLR4/NF-kappaB/ROS pathway and induces mitochondrial dysfunction in bovine oviduct epithelial cells. Am. J. Reprod. Immunol. 77. DOI: e1264228185389 [ Links ]
Paglia, D. & Valentine, W., 1967. Studies on the quantitative and qualitative characterization of erythtrocyte glutathione peroxidase. J. Lab. Clin. Med., 70, 158-169. DOI: 10.5555/uri:pii:0022214367900765 [ Links ]
Ranjan, R., Naresh R., Patra R.C. & Swarup, D., 2006. Erythrocyte lipid peroxides and blood zinc and copper concentrations in acute undifferentiated diarrhoea in calves. Vet. Res. Commun. 30, 249-254. DOI: 10.1007/s11259-006-3185-8 [ Links ]
Rodewald, R., 1976. pH-dependent binding of immunoglobulins to intestinal cells of the neonatal rat. J. Cell. Biol. 7, 1666-16669. DOI: 10.1083/jcb.71.2.666 [ Links ]
Serafini, M.R., Santos, R.C., Guimaraes, A.G., dos Santos, J.P.A., da Conceicao Santos, A.D., Alves, I.A., Gelain, D.P., de Lima Nogueira, P.C., Quintans-Júnior, L.J., Bonjardim, L.R. & de Souza Araújo, A.A., 2011. Morinda citrifolia Linn leaf extract possesses antioxidant activities and reduces nociceptive behaviour and leukocyte migration. J. Med. Food. 14, 1159-1166. https://pubmed.ncbi.nlm.nih.gov/21548805/ [ Links ]
Shankhpal, S.S., Waghela, C.R., Sherasia, P.L., Sridhar, V., Srivastava, A.K. & Singh, D., 2019. Effect of feeding Moringa oleifera as green fodder on feed intake, milk yield, microbial protein synthesis and blood profile in crossbred cows. Ind. J. Anim. Nutr. 36, 228-234. DOI: 10.5958/2231-6744.2019.00038.0 [ Links ]
Sordillo, L.M., 2016. Nutritional strategies to optimize dairy cattle immunity. J. Dairy Sci. 99, 4967-4982. DOI: 10.3168/jds.2015-10354 [ Links ]
Stelwagen, K., Carpenter, E., Haigh, B., Hodgkinson, A. & Wheeler, T.T., 2008. Immune components of bovine colostrum and milk. J. Anim. Sci., 87:3-9. DOI: 10.2527/jas.2008-1377 [ Links ]
Tug, T., Karatas, F., Terzi, S.M. & Ozdemir, N., 2005. Comparison of serum malondialdehyde levels determined by two different methods in patients with COPD: HPLC or TBARS methods. Pathol. 2, 41-44. DOI:10.1309/WTEET9TJ2LUMB3C3 [ Links ]
Van Hoeck, V., Rizos, D., Gutierrez-Adan, A., Pintelon, I., Jorssen, E., Dufort, I., Sirard, M.A., Verlaet, A., Hermans, N., Bols, P.E.J. & Leroy, J.L.M.R., 2015. Interaction between differential gene expression profile and phenotype in bovine blastocysts originating from oocytes exposed to elevated non-esterified fatty acid concentrations. Reprod. Fertil. Dev. 27, 372-384. https://pubmed.ncbi.nlm.nih.gov/24360349/ [ Links ]
Van Soest, P.J, Roberstson, J.B. & Lewis., B.A., 1991. Methods for dietary fibre, neutral detergent fibre and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. DOI: 10.3168/jds.S0022- 0302(91)78551-2 [ Links ]
Wiedemann, M., Kontush, A., Finckh, B., Hellwege, H.H. & Kohlschutter, A., 2003. Neonatal blood plasma is less susceptible to oxidation than adult plasma owing to its higher content of bilirubin and lower content of oxidizable fatty acids. Pediatr. Res. 53, 843-849. DOI: 10.1203/01.PDR.0000057983.95219.0B [ Links ]
Williamson, D., Mellanby, J. & Krebs, H., 1962. Enzymic determination of β-hydroxybutyric acid and acetoacetic acid in blood. Biochem. J. 82, 90-96. DOI: 10.1042/bj0820090 https://pubmed.ncbi.nlm.nih.gov/14007241/ [ Links ]
Windeyer, M.C., Leslie, K.E., Godden, S.M., Hodgins, D.C., Lissemore, K.D. & LeBlanc, S.J., 2014. Factors associated with morbidity, mortality, and growth of dairy heifer calves up to 3 months of age. Prev. Vet. Med., 113, 231-240. https://doi.org/10.1016/j.prevetmed.2013.10.019 [ Links ]
Yu, B.P., 1994. Cellular defenses against damage from reactive oxygen species. Physiol. Rev. 74, 139-162. https://doi.org/10.1152/physrev.1994.74.1.139 [ Links ]
Submitted 9 April 2020
Accepted 5 February 2021
Published 16 September 2021
# Corresponding author: madimetjatk@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}