










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 n.2 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i2.6
ARTICLES
Factors affecting In vitro methane production from cecum contents of White Roman geese
Y.H. ChenI; J.C. HsuII; S.Y. WangIII; P.H. LinIV; J. PerezV; H.W. KuoV, VI, #
IDepartment of Animal Science and Biotechnology, Tunghai University. Taichung, Taiwan
IIDepartment of Animal Science, National Chung Hsing University, Taichung, Taiwan
IIIGraduate Institute of Biotechnology, Chinese Culture University, Taipei, Taiwan
IVDepartment of Animal Science, National Chiayi University, Chiayi, Taiwan
VSustainability Science and Engineering Program, Tunghai University, Taichung, Taiwan
VIDepartment of Environmental Science and Engineering, Tunghai University, Taichung, Taiwan
ABSTRACT
The goal of this research was to gain understanding of in vitro methane (CH4) production from the cecal contents of White Roman geese under various incubation conditions. Five experiments were conducted to ascertain the effects of i) incubation time, ii) pH, iii) the addition of formic acid to the culture media, iv) temperature, and v) the addition of salt to the nutritive liquid. Methane production increased significantly with the supplementation of formic acid in the culture fluid (Experiment III). Additionally, CH4 production Experiment V was higher than that without saline. In contrast, low CH4 production occurred under acidic conditions (pH< 5.4) and at temperatures higher or lower than typical bird body temperature (43 °C) without formic acid and saline solution in the culture media. Since bird body temperature cannot be controlled easily, approaches such as maintaining cecum fluid at low pH and preventing the formation of formic acid by adjusting the recipes of feeds could be considered for controlling in vivo CH4 production from the intestinal tract digesta of geese.
Keywords: body temperature, formic acid, geese farm, methane emission, saline solution
Introduction
Among the greenhouse gases (GHGs), CH4 is a major contributor to global warming effects (Naqvi & Sejian, 2011). It is expected to contribute about 18% of total annual GHG emissions (Milich, 1999; Forabosco et al., 2017). Major CH4 emission sources include landfills, sediment, natural wetlands, flooded paddy fields, sewage treatment works, animal enteric fermentation, and anaerobic fermentation of agricultural wastes (Yang et al., 2003). Among these, animal enteric fermentation has been considered the main source of CH4 production via the digestion processes of ruminants (e.g. cattle, goats and sheep) and non-ruminants (e.g. hogs, horses, chickens, ducks, and geese) (Du Toit et al., 2013; Rendón-Huerta et al., 2018). Additionally, CH4 production from manure produced in the livestock and poultry industries is a major GHG source (Yang et al., 2003; Zhou et al., 2007; Wang et al., 2017). Geese ceca are major places for in vivo CH4 production (Chen et al., 2009; Chen et al., 2014). Chen et al. (2003) showed CH4 production from caecectomized geese was only 8 - 10% of that of sham-operated geese.
In the gastrointestinal systems of ruminant and non-ruminant animals, the cecum provides a habitat for growing microbes that can transform some dietary fibres symbiotically into short-chain fatty acids to provide energy (Gasaway, 1976a, 1976b; Herd & Dawson, 1984). As a result of microbial metabolism, nonprotein nitrogen can be synthesized into amino acids and proteins that can be further digested and absorbed by poultry (Bjornhag & Sperber, 1977; Morternsen & Tindall, 1981). Nutrients of cecal contents in poultry can be fermented and converted into short-chain (2 - 5 carbons) fatty acids and biogas (e.g. ammonia, carbon dioxide and CH4) by microorganisms (Marounek et al., 1999; Chen et al., 2014). Besides, microorganisms cultured from poultry and cattle can generate CH4 (Van Kessel & Russell, 1996; Montagna et al., 2019), suggesting that microorganisms in poultry ceca and bovine rumen fluids have similar functions. Nonetheless, the CH4 production rates from these microbes could differ because of niche conditions (pH and temperature) for diverse animal enteric systems and in vitro tests. Better understanding of CH4 production from animal enteric systems would be helpful in attaining sustainable strategies to lower CH4 emission from poultry farms (Montagna et al., 2019).
Van Kessel and Russell (1996) reported that in vitro CH4 production rates from rumen fluid increased after a period of incubation, but the trend did not follow a linear relationship, probably because of changes of pH in the batches. Chen et al. (2009) showed that in vivo formic acid was the precursor of CH4 in geese, and the pH value in the goose ceca typically ranged between 6.21 and 6.51. However, the effects of pH values and formic acid concentrations on in vitro CH4 generation from goose cecal fluid have not been studied fully, making precise estimation of overall CH4 emission from geese difficult (Zhou et al., 2007). In addition, little is known about the potential influence of other environmental factors (e.g. temperature and salinity) on the methanogenesis of their cecal contents. An incubation temperature of 39 °C was used to investigate methanogenesis of cecal content fermentation of chickens (Tsukahara & Ushida, 2000) and cows (Van Kessel & Russell, 1996; Lalla et al., 1998), whereas 38 °C was used for geese (Chen et al., 2014). Waterfowl (e.g. swans, geese, and ducks) usually have higher body temperatures, ranging from 40 °C to 43 °C (Stanier et al., 1984; Whittow, 1986), but CH4 production from the cecal contents of geese under various in vitro incubation temperatures has not been fully addressed, nor has salinity in the culture media (Chen et al., 2014). Accordingly, this study aimed to investigate the effects of incubation conditions (times, pH, temperature, formic acid concentration, and the presence of saline) on CH4 production from the cecal fluid of White Roman geese to achieve strategies to lower CH4 emission and global warming.
Material and Methods
All the experimental birds were slaughtered at an official slaughterhouse using humane approaches, which are regulated under the Animal Industry Act of 1989 and the Animal Protection Act of 1998 of Taiwan (home slaughter has been forbidden since 1990s). The animal use protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of Tunghai University (THU), Taichung, Taiwan (THU IACUC Approval Number 109-58).
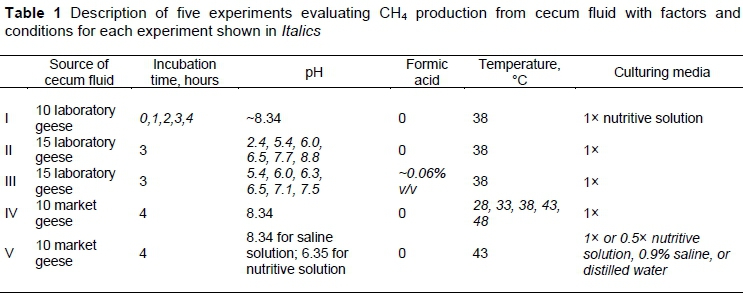
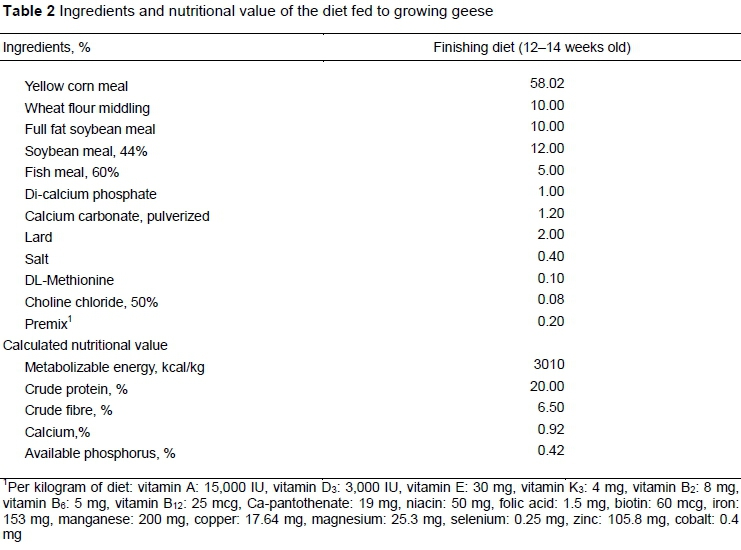
Five experiments were conducted (Table 1). A total of 40 (10, 15, and 15 geese for Experiments I, II, and III, respectively) 14-week-old White Roman geese were used as experimental birds. They were raised in poultry houses and fed a commercial pellet finisher diet (Table 2) with water supplied ad libitum. For Experiments IV and V, fluids were sampled from the ceca of another twenty (10 for each experiment) 14-week-old White Roman geese that were harvested at a local poultry slaughterhouse.
The aim of Experiment I was to study the CH4 production (accumulative amount) from the cecal contents of White Roman geese under six incubation times (0, 0.3, 1, 2, 3, and 4 hours) (n = 5). The test for each incubation time had five replicates. Ten randomly selected 14-week-old birds were slaughtered to collect a mixture of cecal contents. In each test, 1 g cecal content was put into a 15-mL vial containing 3.25 mL of full-strength (1x) nutritive buffer solution (Salvador et al., 1993). The vials were filled with 100% CO2, sealed with butyl rubber stoppers and aluminium caps, and incubated in a 38 °C shaker (80 rpm) under anaerobic conditions. At the end of each incubation time, 0.2 mL of 10% chloride mercury (HgCl2) was added to the vials to terminate bacterial activity. Biogas from each test was sampled from the vial with a gas-tight syringe to measure CH4 concentration.
Experiment II was designed to study the in vitro effect of six pH values (2.4, 5.4, 6.0, 6.5, 7.7, and 8.8) on CH4 production from the cecal contents. A total of 18 tests (n = 3) (triplicate for each pH test) were conducted in 15-mL vials, each containing 1 g cecal content and 3.25 mL mixed nutritive buffer solution in which pH was adjusted to intended values with 10% sodium hydroxide (NaOH) or 6 N hydrochloric acid (Van Kessel & Russell, 1996). The pH values were measured with a pH sensor and meter system (Hanna HI model 8424; Hanna Instruments, Inc., Woonsocket, Rhode Island). The incubation time was three hours. Other preparation conditions were the same as Experiment I.
The aim of Experiment III was to study how supplementation of formic acid affected in vitro CH4 production from the cecal contents of the geese (15 randomly selected birds) at seven levels of pH (5.4, 6.0, 6.3, 6.5, 7.1, 7.5, and 8.3). Each condition was evaluated in triplicate. Experimental preparations were similar to those of Experiments I and II, except for the supplementation of 2 µL formic acid (98%) (Merck, NJ, USA) to the 3.25-mL mixed nutritive buffer solution.
The purpose of Experiment IV was to study the in vitro CH4 production from the cecal contents of the geese (collected from 10 randomly selected birds) under five incubation temperatures (28 °C, 33 °C, 38 °C, 43 °C and 48 °C). Each of the temperature conditions was evaluated in triplicate. Most experimental preparations were as described, whereas an incubation time of four hours and a pH of 8.34 were used.
In Experiment V, in vitro CH4 production was tested in triplicate with four culture fluids (3.25 mL of 1* nutritive buffer solution, half strength (0.5*) nutritive buffer solution, 0.9% physiology saline solution, and distilled water only). Most of experimental preparations were the same as described above, whereas a pH of 6.35 (for saline solution), a pH of 8.34 (for nutritive solution), a temperature of 43 °C, and four hours' incubation were used.
Methane was measured with gas chromatography (Shimadzu, model 14 B) with a FID (flame ionization detector) and a column packed with Porapak Q (Supelco, PA, USA). The oven temperature was 70 °C and the temperature for injection and detector was 130 °C. Nitrogen gas was used as the carrier gas with a flow rate of 10 mL/min. Standards of CH4 (0.5, 10, 50, 100, 500, and 1000 ppm) were prepared by diluting stock CH4 gas (95.5%) (China Petroleum Co.) with nitrogen gas (98.5%) to construct a calibration curve to determine CH4 concentration in each test. The linear calibration curve for CH4 had a R2 value > 0.998 and a coefficient of variation (CV) < 4.7%. Detection of CH4 from 100-ppm standard was used for quality control as the CV value was kept <10%. The amount of CH4 production for each batch was then expressed as microgram CH4 accumulated per gram of cecal content (µg/g), whereas CH4 production rate was shown as microgram CH4 per gram of cecal contents per hour (µg/g/h).
The SAS software was used for statistical analysis (SAS Institute Inc., Cary, North Carolina, USA). Least square means were used to estimate the differences between treatments in each experiment.
Results and Discussion
Results from Experiment I showed amounts of CH4 accumulated for longer incubation times (181.4 and 188.4 µg/g for three- and four-hour tests, respectively) were significantly higher than those for shorter incubation times (9.9 and 65.8 µg/g for 0.3- and 1-hour tests) (Figure 1). Nonetheless, CH4 production seemed to reach saturated level after 3-4 hours incubation, as shown in the notable decrease of CH4 production during this 0 period (Figure 1, bar chart). Only a little CH4 was produced after three hours of incubation, probably because of the gradual exhaustion of substrate in the cecal contents supplied for methanogenesis. In this study, a maximum CH4 production rate of 4.85 µ mole/g/h (or 77.6 µg/g/h) (Figure 1, bar chart) occurred during incubation times of 0.3 - 1 hour. In previous studies, CH4 production rates were 3.5 - 10.3 µ mole/g/h for rumen contents (Hungate et al., 1970), 0.1 - 0.3 µ mole/g/h for ceca of rats, and 8.2 - 11.4 µ mole/g/h for chicken (Tsukahara & Ushida, 2000). These findings suggest that CH4 production from cecum and rumen contents for various animals could be affected by culturing conditions and even microbial community structures in testing mixtures.
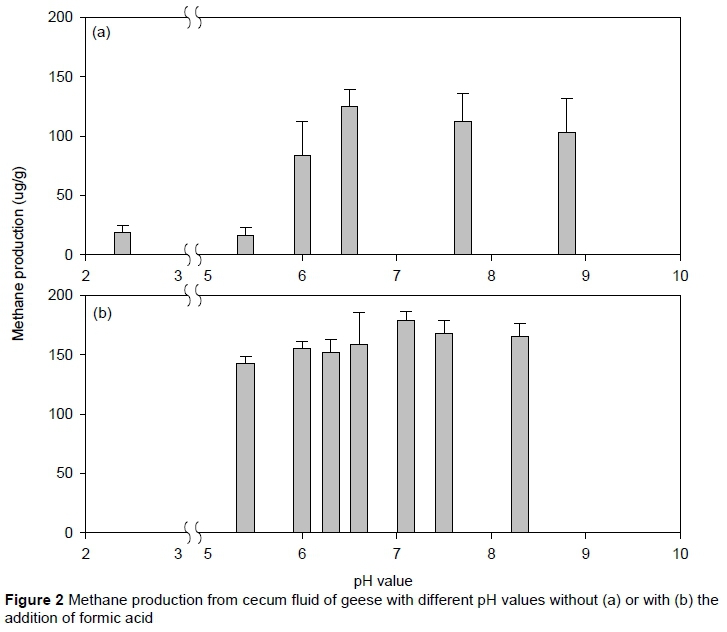
Methane production varied after three hours' incubation under different initial pH (Figure 2). Results showed that the CH4 production for low pH (2.4 and 5.4) batches was significantly lower than for pH 6.0 -8.8. A maximum CH4 production was recorded at a pH of 6.5, but did not show statistical difference (P >0.05) from those for pH 6.0, 7.7, and 8.8. In contrast, little CH4 was produced under acid conditions (pH <5.4). Amount of CH4 produced after three hours' incubation at pH 5.4 (16.4 µg/g) was about five times lower (P <0.05) than for pH 6.0 (83.4 µg/g), suggesting acid conditions would inhibit CH4 production from goose cecum contents, whereas the pH range from 6.0 to 8.8 was relatively more suitable for bacteria activity correlated with CH4 production.
The considerable decrease in CH4 production from geese cecum contents between pH 6.0 and pH 5.4 in this study was similar to a study on cow rumen contents, which showed CH4 production decreased rapidly at pH lower than 6.5 (Van Kessel & Russell, 1996). Another study suggested that methanogenic bacteria were sensitive to pH changes (Fahey & Berger, 1988), probably owing to changes in availability of the hydrogen ion (H+) involved in the microbial metabolism pathway. Besides, low pH conditions would possibly affect acidifying bacteria to convert carbonaceous substrates in cecum contents and culture fluid to short-chain fatty acids (SCFAs) (e.g. formic acid, acetic acid, and propionic acid) - intermediates for anaerobic fermentation and reactants for CH4 production. Consequently, in the culturing mixture, changes in acetate to propionate ratio would occur, which has been suggested to have a high correlation with pH of rumen fluid and capacity of bacteria in producing CH4 from hydrogen and carbon dioxide (Lalla et al., 1998).
When formic acid was added to the culturing mixture, more CH4 was produced than in the previous tests without formic acid addition (Experiment II) (Figure 2b). The amount of CH4 produced under pH 5.4 -8.3 after three hours' incubation ranged from 142.3 to 178.3 µg/g. Maximum CH4 production was found under a pH of 7.1, whereas CH4 production for other pH tests was rather lower, but not significant. Interestingly, under pH 5.4, CH4 production from the test with formic acid (142.3 µg/g) (Figure 2b) seemed not to have been inhibited, as shown in Experiment II without formic acid (16.4 µg/g) (Figure 2a). This suggested that formic acid in the culturing fluid could still be converted into CH4 under a pH of 5.4 by methanogenic bacteria in cecal contents. The finding that low pH (5.4) restrained the capability of the acidification microorganisms for converting carbonaceous substrates in culturing fluid to short-chain fatty acids (including formic acid and acetic acid), but did not have much effect on methanogenic bacteria, as shown in Experiment III, among which most of the formic acid seemed to have been converted into CH4.
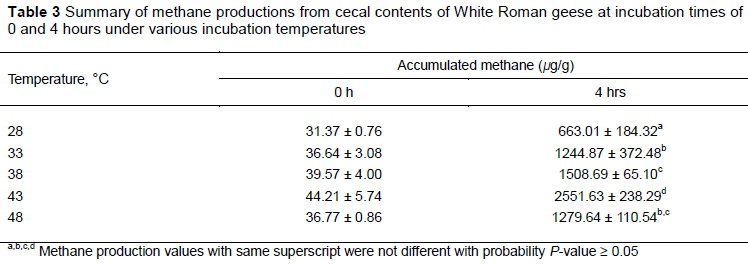
Methane production from the cecum contents was significantly different under various incubation temperatures (Table 3). The highest CH4 production (1508.69 ± 65.1 µg/g) occurred at 43 °C, whereas the lowest (65.01 ± 184.32 µg/g) happened at 28 °C. Low CH4 concentrations could be detected even right after preparation and at the beginning of incubation (time 0). Correlation between incubation temperatures and CH4 production followed linear and quadratic trends well (P <0.001). These results demonstrated that more CH4 would be produced at 43 °C, which was close to the typical temperature of the abdominal cavity of waterfowl (Whittow, 1986; Salvador et al., 1993). Interestingly, CH4 production for the cecum fluid from the local slaughterhouse was significantly different from that from laboratory poultry houses, although some testing conditions for Experiments IV and I were similar (pH ~8.34, temperature 38 °C, and four hours' incubation). Amounts of CH4 produced in Experiment IV were much higher (about tenfold at 38 °C) than those from Experiments I, II, and III. It was surmised that the cecum contents sampled from the birds purchased from the local poultry processing farm had higher substrate contents or more active methanogenic bacteria than those from laboratory-grown birds. To minimize CH4 emission from geese, useful approaches would be to control the availability of substrates and the activity of methanogenic microbes.
Earlier studies showed different optimal temperatures for CH4 production from cow rumen fluid (39 °C) (Lalla et al., 1998), paddy field soil (34.5 °C) (Parashar et al., 1993), and subarctic peat soil (25 °C) (Dunfield et al., 1993), which were all lower than the 43 °C determined for geese cecum fluid in this study. It was surmised that microbial composition in the geese cecum fluid tested in this study would be different from samples used for previous studies. Microbial species or strains that were well-adapted to conditions in the ceca would become dominant in the cecum fluid samples and would show high CH4 production ability under a temperature close to typical bird body temperature. This may provide crucial ideas for suitable anaerobic microbes and optimal operating parameters for engineered processes for anaerobic biodegradation of particular waste. Such information could also be used to amend strategies for minimizing CH4 production from uncontrolled sources when CH4 gas was not considered for recovery as a biofuel source.
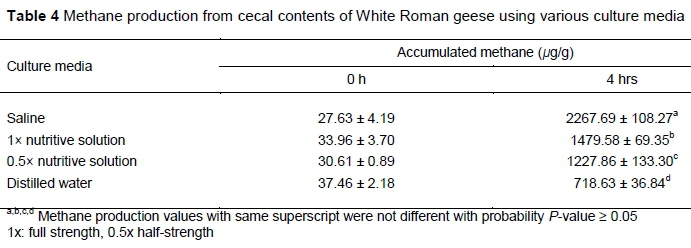
Methane production after four hours' incubation from the batch tests with saline solution was higher than from those using 1* and half-strength nutritive buffer or just distilled water (Table 4). These results suggested that the nutritive buffer solution and saline solution enhanced CH4 production from the cecum contents. Interestingly, the batch using distilled water without nutritive buffer showed significant CH4 production (from 37.46 µg/g at time 0 to 718.63 µg/g at four hours). This implied that geese cecum fluids already contain nutrients that are required to metabolize anaerobic methanogenic microbes. Additions of 1* nutritive solution or 0.9% saline solution seemed to create better niche conditions for in vitro microbial CH4 production. In contrast, less rich (or oligotrophic) culturing conditions would result in lower CH4 production, which would ease a little the global warming effect of in vivo CH4 production from animal intestinal systems.
Since several heavy metals (e.g. zinc, copper, and manganese) were included in the nutritive solution as trace elements and different microbes may prefer certain optimal concentrations, inappropriate concentrations could lead to adverse effects on microbial activity. Earlier studies showed the toxicity of heavy metals on microorganisms (Chaudri et al., 1992; Giller et al., 1998). In addition, salt (sodium chloride) seemed to affect CH4 production, even when a nutritive buffer solution containing minerals was used broadly to test CH4 production from ruminal fluids (Van Kessel & Russell, 1996), human faeces (Salvador et al., 1993), and chicken cecum fluids (Tsukahara & Ushida, 2000). Methane production from the samples with saline solution (0.9% w/v) was about 1.53 times higher than from the samples using 1* nutritive solution (with a sodium chloride concentration of 0.047% w/v). Besides, based on results from Experiment II, the saline solution with a pH of 6.35 would be more suitable for CH4 production than the nutritive solution with a pH of 8.34.
Conclusion
Because bird body temperature cannot be regulated easily, maintaining the flow of caecum fluid at low pH and preventing the formation of formic acid by adjusting the diet might be considered when attempting to control in vivo CH4 production in the intestinal tract of geese. These findings provide insight into in vitro CH4 production from geese cecum fluids under various conditions and can be used for its estimation and control.
Acknowledgements
This study was financially supported by the Tunghai Green Energy Development and Management Institute (TGEI) of Tunghai University. The authors would like to express their gratitude to Miss Chi-Yin Chiou and Mr. Wen-Wan Kuo at the Department of Animal Science, Chinese Culture University, for their help with animal management, sample collection, and methane gas detection.
Authors' Contributions
CY-H (ORCID 0000-0002-8870-4103) designed the experiments, collected and analysed the data, and prepared the manuscript; HJ-C (ORCID 0000-0001-6158-196X) designed the experiments and supervised the progress of the studies; WS-Y (ORCID 0000-0003-1566-5542) collected and analysed the data; LP-H (ORCID 0000-0001-5140-1195) collected and analysed the data; JP (ORCID 0000-0002-5746-9542) prepared the manuscript; KH-WD (ORCID 00000002-7612-6879) participated in data analysis and manuscript preparation.
Conflict of Interest Declaration
There were no conflicts of interest regarding this work.
References
Bjornhag, G. & Sperber, I., 1977. Transport of various food components through the digestive tract of turkey, geese, and guinea fowl. Swedish J. Agric. Res. 7, 57-66. [ Links ]
Chaudri, A.M., McGrath, S.P. & Gille, K.E., 1992. Survival of the indigenous population of Rhizobium leguminosarum biovar trifolii in soil spiked with Cd, Zn, Cu and Ni salt. Soil Biol. Biochem. 24, 625-632. https://doi.org/10.1016/0038-0717(92)90040-5 [ Links ]
Chen, Y.H., Wang, S.Y. & Hsu, J.C., 2003. Effects of caecectomy on body weight gain, intestinal characteristics and enteric gas production in goslings. Asian-Aust. J. Anim. Sci. 16, 1030-1034. DOI: 10.5713/ajas.2003.1030 [ Links ]
Chen, Y.H., Wang, S.Y. & Hsu, J.C., 2009. In vivo methane production from formic and acetic acids in the gastrointestinal tract of White Roman geese. Asian-Aust. J. Anim. Sci. 2009; 22(7), 1043-1047. DOI: 10.5713/ajas.2009.80319 [ Links ]
Chen, Y.H., Lee, S.M., Hsu, J.C., Chang, Y.C. & Wang, S.Y., 2014. Methane generation from the intestine of Muscovy ducks, mule ducks and White Roman geese. Aerosol Air Qual. Res. 14, 323-329. DOI: 10.4209/AAQR.2013.05.0180 [ Links ]
Du Toit, C.J.L., Van Niekerk, W.A. & Meissner, H.H., 2013. Direct greenhouse gas emissions of the South African small stock sectors. S. Afr. J. Anim. Sci. 43(3), 340-361. DOI: 10.4314/sajas.v43i3.8 [ Links ]
Dunfield, P., Knowles, R., Dumont, R. & Moore, T.R., 1993. Methane production and consumption in temperate and subArctic peat soils: Response to temperature and pH. Soil Biol. Biochem. 25, 321-326. https://doi.org/10.1016/0038-0717(93)90130-4 [ Links ]
Fahey, G.C. Jr. & Berger L.L., 1988. Carbohydrate nutrition of ruminants. In: D.C. Church (ed). The ruminant animal digestive physiology and nutrition. Prentice Hall, New Jersey, USA. [ Links ]
Forabosco, F., Chitchyan, Z.H. & Mantovani, R., 2017. Methane, nitrous oxide emissions and mitigation strategies for livestock in developing countries: A review. S. Afr. J. Anim. Sci. 47(3), 268-280. http://dx.doi.org/10.4314/sajas.v47i3.3 [ Links ]
Gasaway, W.C., 1976a. Seasonal variation in diet, volatile fatty acid production and size of the cecum of rock ptarmigan. Comp. Biochem. Physiol. 54A, 109-114. https://doi.org/10.1016/S0300-9629(76)80021-7 [ Links ]
Gasaway, W.C., 1976b. Volatile fatty acids and metabolizable energy derived from cecal fermentation in the willow ptarmigan. Comp. Biochem. Physiol. 53A, 115- 121. DOI: 10.1016/s0300-9629(76)80022-9 [ Links ]
Giller, K.E., Witter, E. & McGrath, S.P., 1998. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 30, 1389-1414. https://doi.org/10.1016/S0038-0717(97)00270-8 [ Links ]
Herd, R. & Dawson, T., 1984. Fiber digestion in the emu, Dromaius novaehollandiae, a large bird with a simple gut and high rates of passage. Physiol. Zool. 57, 70-84. https://doi.org/10.1086/physzool.57.L30155970 [ Links ]
Hungate, R.E., Smith, W., Bauchop, T., Yu, I. & Rabinowitz, J.C., 1970. Formate as an intermediate in the bovine rumen fermentation. J. Bacteriol. 102, 389- 397. DOI: 10.1128/JB.102.2.389-397.1970 [ Links ]
Lalla, R.P., Russell, J.B. & Van Aniburgh, M.E., 1998. The role of pH in regulating ruminal methane and ammonia production. J. Anim. Sci. 76, 2190-2196 DOI: 10.2527/1998.7682190x [ Links ]
Marounek, M., Suchorska, O. & Savka, O., 1999. Effect of substrate and feed antibiotics on in vitro production of volatile fatty acids and methane in caecal contents of chickens. Anim. Feed Sci. Technol. 80, 223-230. https://doi.org/10.1016/S0377-8401(99)00065-6 [ Links ]
Milich, L., 1999. The role of methane in global warming: Where might mitigation strategies be focused? Global Environ. Change 9, 179-201. https://doi.org/10.1016/S0959-3780(98)00037-5 [ Links ]
Montagna, F., Scripnic, E. & Forabosco, F., 2019. Intensification of production, low emission pathways and sustainable strategies for backyard, layer and broiler chickens. S. Afr. J. Anim. Sci. 49(3), 534-545. http://dx.doi.org/10.4314/sajas.v49i3.14 [ Links ]
Mortensen, A. & Tindall, A. 1981. Caecal decomposition of uric acid in captive and free ranging willow ptarmigan (Lagopus lagopus lagopus). Acta Physiol. Scand. 111, 129- 133. DOI: 10.1111/j.1748-1716.1981 .tb06715.x [ Links ]
Naqvi, S.M.K. & Sejian, V., 2011. Global climate change: Role of livestock. Asian J. Agric. Sci. 3(1), 19-25. https://www.researchgate.net/publication/49605076 [ Links ]
Parashar, D.C., Gupta, P.K., Rai, J., Sharma, R.C. & Singh, N., 1993. Effect of soil temperature on methane emission from paddy fields. Chemosphere 26, 247-250. https://doi.org/10.1016/0045-6535(93)90425-5 [ Links ]
Rendón-Huerta, J.A., Pinos-Rodriguez, J.M., Kebreab, E., Garcia-López, J.C. & Vicente, J.G., 2018. Comparison of greenhouse gas emissions from Mexican intensive dairy farms. S. Afr. J. Anim. Sci. 48(1), 48-55. http://dx.doi.org/10.4314/sajas.v48iL6 [ Links ]
Salvador, V., Cherbut, C., Barry, J.L., Bertrand, D., Bonnet, C. & Delort-Laval, J., 1993. Sugar composition of dietary fibre and short-chain fatty acid production during in vitro fermentation by human bacteria. Br. J. Nutr. 70, 189-197. DOI: 10.1079/bjn19930116 [ Links ]
Stanier, M.W., Mount, L.E. & Bligh, J., 1984. Energy balance and temperature regulation. Cambridge University Press, Cambridge. [ Links ]
Van Kessel, J.S. & Russell, J.B., 1996. The effect of pH on ruminal methanogenesis. FEMS Microbiol. Ecol. 20, 205210. https://doi.org/10.1111/j.1574-6941.1996.tb00319.x [ Links ]
Wang, L.-Z., Xue, B. & Yan, T., 2017. Greenhouse gas emissions from pig and poultry production sectors in China from 1960 to 2010. J. Integr. Agric. 16(1), 221-228. DOI: 10.1016/S2095-3119(16)61372-2 [ Links ]
Whittow, G.C., 1986. Regulation of body temperature. In: P.D. Strukie (ed). Avian physiology. Springer-Verlag, New York. [ Links ]
Yang, S.S., Liu, C.M. & Liu, Y.L., 2003. Estimation of methane and nitrous oxide emission from animal production sector in Taiwan during 1999-2000. Chemosphere 52, 1381-1388. DOI: 10.1016/S0045-6535(03)00473-9 [ Links ]
Zhou, J.B., Jiang, M.M. & Chen, G.Q., 2007. Estimation of methane and nitrous oxide emission from livestock and poultry in China during 1949-2003. Energy Policy 35, 3759-3767. https://doi.org/10.1016/j.enpol.2007.01.013 [ Links ]
Submitted 16 March 2020
Accepted 7 January 2021
Published 8 March 2021
# Corresponding author: hwkuo@thu.edu.tw


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}