










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.51 no.1 Pretoria 2021
http://dx.doi.org/10.4314/sajas.v51i1.8
ARTICLES
Effects of dietary fibre level on rumen pH, total microbial count and methanogenic archaea in Bonsmara and Nguni steers
S.T. JiyanaI, II, #; M.M. RatsakaIII; K-J. LeeuwI; K.R. MbathaII
IAgricultural Research Council, Animal Production Institute, Animal Nutrition Section, Irene, South Africa
IIUniversity of South Africa, Science Campus, Department of Agriculture and Animal Health, Florida, South Africa
IIITshwane University of Technology, Department of Animal Science, Pretoria, South Africa
ABSTRACT
A study was conducted to evaluate the effect of the level of dietary fibre on rumen pH, total microbial count and methanogenic archaeacount of Bonsmara (225 ± 10.0 kg live weight) and Nguni (215 ± 10.0 kg live weight) steers. Nine steers per breed were used, aged 8 to 9 months. A 3 x 2 factorial experiment was executed with treatments allocated in a completely randomized design. The treatments were diets which included 64.3% neutral detergent fibre (NDF) for diet 1, 40.4% NDF for diet 2, and 29.9% NDF for diet 3. Steers were fed for 90 days and housed in individual pens. Rumen fluid was collected from steers using a stomach tube. Rumen fluid samples were taken immediately to the laboratory for microbial assay. Data were subjected to analysis of variance (ANOVA). The various levels of dietary fibre did not affect rumen pH and methanogenic archaea count. However, a low-fibre diet yielded high rumen total microbial count for Bonsmara steers and low rumen total microbial count for Nguni steers. Therefore, a low-fibre diet can be applied as a viable strategy to enhance rumen total microbial count in exotic beef breeds and crossbred cattle such as Bonsmara. Nguni steers fed a high NDF diet had higher rumen total microbial count than Nguni steers fed a low-fibre diet. Therefore, a high NDF diet can be used efficiently by feeding it to indigenous breeds and purebred cattle such as Nguni.
Key words: Enteric fermentation, lactic acidosis, rumen function, methanogenic archaea
Introduction
The exponential growth of human population in the world compels farmers to improve livestock production to meet the increasing demand for animal protein (Delgado, 2003). Feedlot operators view cattle with a shorter production cycle, rapid growth and better feed efficiency as ideal to supply the quality protein required for the growing human population (Smil, 2002). In South Africa, most feedlot markets prefer exotic crossbred animals, such as Bonsmara, because of their outstanding feed efficiency under feedlot conditions, to indigenous and purebred animals such as Nguni (Strydom, 2008). Nguni cattle that were fed a high-fibre diet showed high feed efficiency compared with the Nguni fed a low-fibre diet (Linde et al., 2016). This shows the ability of Nguni cattle to utilize high-fibre diets efficiently. When an animal consumes a high-fibre diet, fibre-digesting microbes such as Fibrobacter spp proliferate in its digestive system, which results in balanced microbial activity with normal (6 - 7) rumen pH (Brown et al., 2006; Owens et al., 1998). However, if the same animal changes to a low-fibre diet, starch-digesting microbes such as amylolytic bacteria proliferate in its digestive system, converting starch into lactic acid and ultimately reducing the rumen pH (Reddy et al., 2008). Fibrous feedstuffs are commonly incorporated in diets to prevent the excessive dropping of rumen pH that may be encountered with low-fibre feedlot diets (Meissner et al., 2010).
The main global concern with high-fibre diets is the production of methane, which is one of the greenhouse gases (Morgavi et al., 2010; Wright et al., 2004). Methanogenic archaea produce methane when they synthesize energy for growth by using hydrogen to reduce carbon dioxide, formed as products of the microbial feed digestion process (Hook et al., 2010). Several rumen microbes are involved in enteric feed fermentation (Henderson et al., 2015). However, the rumen microbial population and its diversity depend on the nutrient composition of the diet consumed by the host animal (Kamra, 2005). Methanogenic archaea, like most other rumen microbes, are sensitive to low levels of ruminal pH (Popova et al., 2012). Feeding beef cattle with extremely low-fibre diets might result in ruminal pH level below 6.0 for prolonged periods (Hünerberg et al., 2015), which is not conducive to rumen microbial activities such as enteric fermentation (Russell & Rychlik, 2001).
The literature recommends that NDF should be maintained at 25% of dietary dry matter with at least 75% from forage for efficient rumen functioning (Banakar et al., 2018; Vermaak, 2011). There is a need to determine the level of dietary fibre at which methanogenic archaea would be reduced without affecting the rumen pH and total microbial count (rumen functioning) negatively. Supplemental fats, starch, ionophores, concentrates, halogenated compounds, condensed tannins, probiotic yeast products and direct-fed microbials are some of nutritional interventions that have been tested and applied to reduce methane production in ruminants (Vohra et al., 2016; Jeyanathan et al., 2014; Beauchemin et al., 2008; Guan et al., 2006; Yan et al., 2000; Johnson & Johnson, 1995). However, most of these nutritional interventions are costly and thus not practical for poorly resourced beef cattle farmers, especially those under communal farming systems. The purpose of this study was to evaluate the effect of the level of dietary fibre on rumen pH, total microbial count and methanogenic archaea count of Bonsmara and Nguni steers.
Materials and Methods
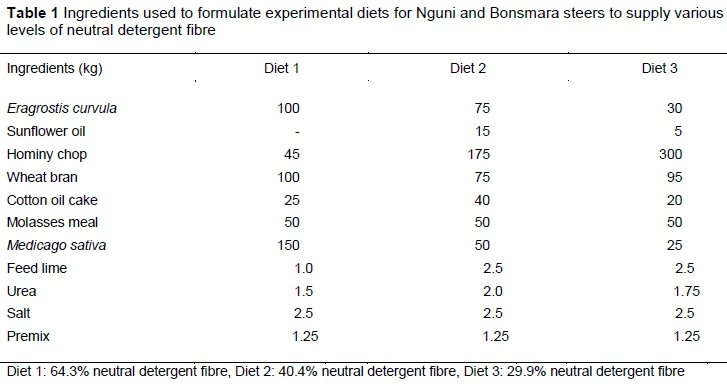
This study was conducted at Agricultural Research Council-Animal Production (ARC-AP) Institute in Irene, Pretoria. Ethical approvals were obtained from the ARC-AP Research Ethics Committee (Ref: APIEC15/047) and University of South Africa, College of Agriculture and Environmental Sciences (Ref: 2015/CAES/130). Nine Bonsmara steers and nine Nguni steers received three dietary treatments with various levels of neutral detergent fibre (NDF). These were 64.3% for diet 1, 40.4% for diet 2, and 29.9% for diet 3. A 3 x 2 factorial experiment (three dietary treatments and two cattle breeds) was used with treatments being allocated in a completely randomized design. The steers were adapted gradually to the experimental diets for 14 days to avoid ruminal acidosis and bloat (Klieve et al., 2003). They were fed 2.5% of their live bodyweight on a dry matter basis each day. The orts were removed from the feed bunks and weighed weekly. Feed consumed was taken as the difference between the amount fed and the orts. The ingredients used in the formulation of treatment diets are presented in Table 1.
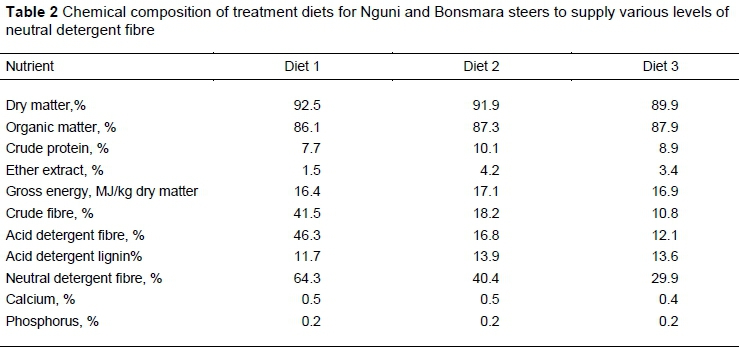
The dry matter content of the diets was determined by drying freshly mixed samples of each diet in a drying oven at 60 °C for 48 hours, following the procedure of AOAC (2010). Dried diet samples were then ground through a 1-mm screen (Wiley mill, Standard Model 3, Arthur H. Thomas Co., Philadelphia, PA) and stored in labelled bottles for chemical analyses. Crude protein (CP), organic matter (OM) and ether extract (EE) were analysed according to AOAC (2010). Acid detergent fibre (ADF), neutral detergent fibre (NDF), acid detergent lignin (ADL) and crude fibre (CF) were determined according to the technique prescribed by Van Soest et al. (1991). Gross energy (GE) was assessed with a bomb calorimeter (MC-1000 modular calorimeter, energy instrumentation, 135 Knoppieslaagte, Centurion, South Africa). For the analysis of phosphorus (P) and calcium (Ca), the feed samples were compressed in a ring vessel to produce a pellet, which was then dried to a residual moisture of <3% for 3 - 4 hours at 60 °C before X-ray fluorescence analysis (West et al., 2013). Table 2 shows the chemical composition of the diets.
A stomach tube was used to collect rumen fluid from steers fed the three diets (Geishauser, 1993). It was rinsed thoroughly with distilled water before and after every collection to avoid cross contamination of the samples from different animals. Three samples per treatment were collected. Rumen fluid was collected within two minutes per animal. The fluid was then poured into a pre-warmed collecting flask, sealed and taken to the laboratory within 20 minutes. Rumen fluid was filtered through two layers of cheesecloth and kept at 39 °C under constant flushing with CO2. Fresh rumen fluid samples were then transferred into the anaerobic cabinet to ensure that rumen fluid samples were not exposed to oxygen to retain the anaerobic condition. A pH meter was used to measure pH level (Duffield et al., 2004) from the rumen fluid samples. Monitoring rumen pH level is one of viable strategies for diagnosing acidosis (Enemark, 2009).
The method for determining microbial count from rumen fluid was adapted from Mahadevan et al. (1982). One ml of fresh rumen fluid was added to 9 ml volumes of anaerobic diluent. The process continued with tenfold dilution of the original sample. One ml of the dilution was placed in labelled sterile disposable petri dish. The same procedure was carried out for a control dish that contained no sample and 1 ml of un-inoculated diluent. Fifteen ml per plate of MRS agar solution was added as a growth media. The contents of the dishes were then swirled to achieve thorough mixing. The agar was allowed to set. The dishes were inverted and stacked in transparent plastic canisters to protect the contents against desiccation. Incubation at 39 °C in an oxygen-free atmosphere took place for 48 hours. The control dish was also incubated and examined for possible contamination. After incubation, the colonies on each inoculated petri dish containing between 30 and 300 colonies were counted. The figures were then multiplied by the dilution factor to obtain colony-forming units (cfu) per gram/ml of sample.
The method for determining rumen methanogenic archaea count was adapted from Carberry et al. (2014). The DNA extraction from rumen fluid was done with a DNA extraction kit (Inqaba BiotecTM, Africa's Genomics Company, Pretoria, RSA) according to the manufacturer's instructions. Universal primers were used to detect rumen archaea counts, using a real-time PCR molecular method. Quantification of methanogenic archaea counts was performed by quantitative real-time PCR (qRT-PCR) analysis of the 16S rRNA genes.
Shapiro-Wilk's test was performed to test for non-normality. The data were analysed using analysis of variance procedures of SAS (SAS Institute Inc., Cary, North Carolina, USA). The linear model included the main effects of breed and diet, and the interaction between them. Student's t-tests at the 5% level of probability were used to compare the treatment means.
Results and Discussion
There was no difference (P >0.05) in rumen pH between diets and breeds (Figure 1). However, steers fed the low-fibre diet had rumen pH of 6.3 for Bonsmara and 6.4 for Nguni, indicating that 29.9% NDF of diet 3 was enough to maintain adequate rumen buffering capacity. This may also result from the steers being gradually adapted to experimental diets to avoid a sudden change from high- to low-fibre diets (Klieve et al., 2003). An extreme drop in rumen pH usually occurs when ruminants are suddenly introduced to low-fibre diets without restriction (Reddy et al., 2008). This abrupt change leads to lactic acid accumulation in the rumen because of a shift in microbial populations from gram-negative predominance to gram-positive lactic acid producers (Meissner et al., 2014). These results were supported by the outcomes of Brown et al. (2006), who reported that a high-fibre diet resulted in balanced microbial activity and a normal (6 - 7) rumen pH level.
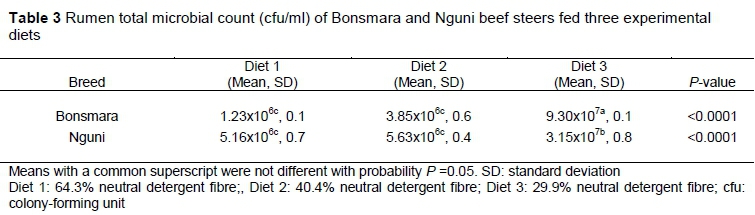
Bonsmara steers fed the low-fibre diet had a higher (P <0.05) rumen total microbial count than Nguni steers on the same diet (Table 3). This might be because Bonsmara cattle are bred to perform better under feedlot systems (Muchenje et al., 2008). These results concur with those of Guan et al. (2008), who found that the presence of particular rumen microbes relied on the interaction between animal genetic make-up and diet composition. High rumen total microbial count is also an indication of an ideal rumen environment and good rumen function (Russell & Rychlik, 2001), which depend on the feed efficiency of the host animal and the fermentation characteristics of the feed (Hutjens, 1991). Nguni fed a high NDF diet had higher (P <0.05) total microbial count than Nguni fed a low NDF diet. These results are supported by the findings that Nguni fed a high NDF diet had better feed efficiency than the Nguni fed a low NDF diet (Linde et al., 2016). When fed the low-fibre diet, rumen total microbial count for the two breeds differed (P <0.05) from the time that they were provided with more fibre (Table 3).
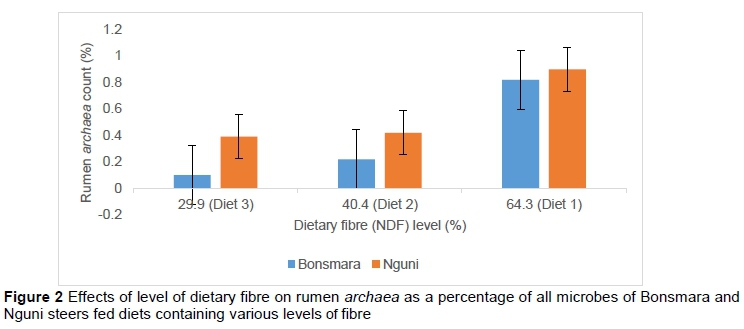
There was no difference (P >0.05) in rumen methanogenic archaea count between diets or between breeds. However, rumen methanogenic archaea count in steers fed diet 3 was 0.1% for Bonsmara and 0.4% for Nguni (Figure 2). This was consequently associated with rumen pH level of 6.3 for Bonsmara and 6.4 for Nguni, which were lowest among the diets. These results are supported by the observations of Hünerberg et al. (2015), who found that methanogens are sensitive to low rumen pH levels. On the other hand, rumen methanogenic archaea count in steers fed diet 1 was 0.8% for Bonsmara and 0.9% for Nguni (Figure 2). Rumen methanogenic archaea count for Bonsmara tended to be lower than for Nguni across all treatment diets (Figure 2). Whitford et al. (2001) reported that rumen archaea count was associated with the feed efficiency of a host animal, and Strydom et al. (2001) did not report substantial differences between Bonsmara and Nguni in feed conversion.
Conclusions
Dietary fibre level did not affect the rumen pH and methanogenic archaea count. However, feeding a low NDF diet resulted in high rumen total microbial count for Bonsmara steers and low rumen total microbial count for Nguni steers. Consequently, NDF of 29.9% was sufficient to ensure adequate rumen buffering capacity, which is crucial to the survival of essential rumen microbes and efficient enteric fermentation. On the other hand, Nguni steers fed a high NDF diet had higher total microbial count than Nguni steers fed a low NDF diet. A low-fibre diet can be applied as a viable strategy to enhance rumen total microbial count in crossbred cattle such as Bonsmara. A high-fibre diet can be used efficiently by feeding it to indigenous breeds and purebred cattle such as Nguni, since they are able to yield high rumen total microbial count when they are fed high-fibre diets. Further research on the diversity of rumen microbial populations should be carried out to contrast the numbers of beneficial versus detrimental microbes and additional insight into growth performance and methane emission of these two beef breeds.
Acknowledgements
The authors would like to offer their sincere gratitude to these institutions for supporting this study: Gauteng Department of Agriculture and Rural Development is highly appreciated for funding; Agricultural Research Council -Animal Production Institute (Irene) for research infrastructure; National Research Foundation for funding through Scarce Skills Master's Scholarship; University of South Africa for capacity building and funding through the postgraduate research bursary. Reviewers and editors of South African Journal of Animal Science are highly appreciated for constructive comments that improved the quality of this manuscript.
Authors' Contributions
STJ, MMR, K-JL & KRM were in charge of project design and writing the manuscript. STJ, MMR & K-JL were in charge of project implementation. All co-authors participated in analysis of the data, and tabulation and interpretation of the results. This manuscript has been read and approved by all of the authors.
Conflict of Interest Declaration
The authors wish to affirm that there are no conflicts of interest associated with this work.
References
AOAC, 2010. Official methods of analysis. 18th edition. Association of Official Analytical Chemists, Washington, DC. DOI: http://hdl.handle.net/10637/3158 [ Links ]
Banakar, P.S., Anand Kumar, N., Shashank, G.C. & Neeti, L., 2018. Physical effective fibre in ruminant nutrition: A review. J. Pharmacogn. Phytochem. 7, 303-308. http://www.phytojournal.com/archives/2018/vol7issue4/PartF/7-3-580-454.pdf [ Links ]
Beauchemin, K.A., Kreuzer, M., Mara, F.O. & McAllister, T.A., 2008. Nutritional management for enteric methane abatement: a review. Aust. J. Exp. Agric. 48, 21-27. https://doi.org/10.1071/ea07199 [ Links ]
Brown, M.S., Ponce, C.H. & Pulikanti, R., 2006. Adaptation of beef cattle to high concentrate diets: Performance and ruminal metabolism. J. Anim. Sci. 81, 25-33. https://doi.org/10.2527/2006.8413_supplE25x [ Links ]
Carberry, C.A, Kenny, D.A, Kelly, A.K. & Waters, S.M., 2014. Quantitative analysis of ruminal methanogenic microbial populations in beef cattle divergent in phenotypic residual feed intake (RFI) offered contrasting diets. J. Anim. Sci. Biotechnol. 5, 41. http://www.jasbsci.com/content/5/1/41 [ Links ]
Delgado, C.L., 2003. Rising consumption of meat and milk in developing countries has created a new food revolution. J. Nutr. 133, 3907S-3910S. https://doi.org/10.1093/jn/133.11.3907S [ Links ]
Duffield, T., Plaizier, J.C., Fairfield, A., Bagg, R., Vessie, G., Dick, P., Wilson, J., Aramini, J. & McBride, B., 2004. Comparison of techniques for measurement of rumen pH in lactating dairy cows. J. Dairy Sci. 87, 59-66. https://doi.org/10.3168/jds.S0022-0302(04)73142-2 [ Links ]
Enemark, J.D.M., 2009. The monitoring, prevention and treatment of sub-acute ruminal acidosis (SARA): A review. Vet. J. 176, 32-43. DOI: 10.1016/j.tvjl.2007.12.021 [ Links ]
Geishauser, T., 1993. An instrument for collection and transfer of ruminal fluid and for administration of water-soluble drugs in adult cattle. Bovine Practit. 27, 37-42. https://journals.tdl.org/bovine/index.php/bovine/article/view/372 [ Links ]
Guan, H., Wittenberg, K.M., Ominski, K.H. & Krause, D.O. 2006. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. 84(7), 1896-1906. https://doi.org/10.2527/jas.2005-652 [ Links ]
Guan, L.L., Nkrumah, J.D., Basarab, J.A. & Moore. S.S., 2008. Linkage of microbial ecology to phenotype: correlation of rumen microbial ecology to cattle's feed efficiency. FEMS Microbiol. Lett. 288, 85-91. https://doi.org/10.1111/j.1574-6968.2008.01343.x [ Links ]
Henderson, G., Cox, F., Ganesh, A., Young, W. & Janssen, P.H., 2015. Rumen microbial community varies with diet and host, but core microbiome is found across a wide geographical range. Sci. Rep. 5, 14567. DOI: 10.1038/srep19175 [ Links ]
Hook, S.E., Wright, A.G. & McBride, B.W., 2010. Methanogens: Methane producers of the rumen and mitigation strategies. Hindawi, Archaea, Canada. DOI: 10.1155/2010/945785 [ Links ]
Hünerberg, M., McGinn, S.M., Beauchemin, K.A., Entz, T., Okine, E.K., Harstad, O.M. & McAllister, T.A., 2015. Impact of ruminal pH on enteric methane emissions. J. Anim. Sci. 93(4), 1760-1766. DOI: 10.2527/jas2014-8469 [ Links ]
Hutjens, M.F., 1991. Feed additives. Veterinary Clinics of North America. Food Anim. Pract. 7, 525-540. DOI: 10.1016/s0749-0720(15)30789-1 [ Links ]
Jeyanathan, J., Martin, C. & Morgavi, D.P. 2014. The use of direct-fed microbials for mitigation of ruminant methane emissions: A review. Animal 8, 250-261. https://doi.org/10.1017/S1751731113002085 [ Links ]
Johnson, K.A. & Johnson, D.E. 1995. Methane emissions from cattle. J. Anim. Sci. 73(8), 2483-2492. https://doi.org/10.2527/1995.7382483x [ Links ]
Kamra, D.N., 2005. Rumen microbial ecosystem. Curr. Sci. 89, 124-135. https://pdfs.semanticscholar.org/8317/cba7556f2e17b3b9c69cd903ead90c63eab2.pdf [ Links ]
Klieve, A.V., Hennessy, D., Ouwerkerk, D., Foster, R.J. & Mackie, R.I. & Attwood, G.T., 2003. Establishing population of Megasphaera elsdenii YE 34 and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets. J. Appl. Microbiol. 95, 621-630. https://pubmed.ncbi.nlm.nih.gov/12911711/ [ Links ]
Linde, D.A., Scholtz, M.M., Theunissen, A. & Marle-Köster, E., 2016. Feedlot performance of South African Nguni cattle fed low and high energy rations. Proc. 1st Int. Conf. Trop. Anim. Sci. Prod. (TASP). 26-29 July 2016, Bangkok, Thailand. pg. 287-290. [ Links ]
Mahadevan, S., Sauer, F.D., Erfle, J.D., Teather, R.M. & Morse, P.M., 1982. Changes in ammonia concentrations, bacterial counts, pH and volatile fatty acid concentration in rumen of cows fed alfalfa hay or concentrate: Urea corn silage. Can. J. Anim. Sci. 62, 249-258. https://doi.org/10.4141/cjas82-025 [ Links ]
Meissner, H.H., Henning, P.H., Leeuw, K-J., Hagg, F.M., Horn, C.H., Kettunen, A. & Apajalahti, J.H.A., 2014. Efficacy and mode of action of selected non-ionophore antibiotics and direct-fed microbials in relation to Megasphaera elsdenii NCIMB 41125 during in vitro fermentation of an acidosis-causing substrate. J. Livest. Sci. 162, 115-125. http://dx.doi.org/10.1016/Llivsci.2014.01.026 [ Links ]
Meissner, H.H., Henning, P.H., Horn, C.H., Leeuw, K-J., Hagg, F.M. & Fouché, G., 2010. Ruminal acidosis: A review with detailed reference to the controlling agent Megasphaera elsdenii NCIMB 41125. S. Afr. J. Anim. Sci. 40, 79-100. http://dx.doi.org/10.4314/sajas.v40i2.57275 [ Links ]
Morgavi, D.P., Forano, E., Martin, C. & Newbold, C.J., 2010. Microbial ecosystem and methanogenesis in ruminants. Animal 4, 1024-1036. DOI: 10.1017/S1751731110000546 [ Links ]
Muchenje, V., 2008. Tick susceptibility and its effects on growth performance and carcass characteristics of Nguni, Bonsmara and Angus steers raised on natural pasture. Animal 2, 298-304. DOI: 10.1017/S1751731107001036 [ Links ]
Owens, F.N., Secrist, D.S., Hill, W.J. & Gill, D.R., 1998. Acidosis in cattle: A review. J. Anim. Sci. 76, 275-286, https://doi.org/10.2527/1998.761275x [ Links ]
Popova, M., Morgavi, D.P. & Martin, C., 2012. Methanogens and methanogenesis in the rumens and ceca of lambs fed two different high grain content diets. Appl. Environ. Microbiol. 79, 1777-1786. DOI: 10.1128/AEM.03115-12 [ Links ]
Reddy, G., Altaf, M., Naveena, B.J., Venkateshwar, M. & Kumar, E.V., 2008. Amylolytic bacterial lactic acid fermentation - a review. Biotechnol. Adv. 26, 22-34. DOI: 10.1016/j.biotechadv.2007.07.004 [ Links ]
Russell, J.B. & Rychlik, J.L., 2001. Factors that alter rumen microbial ecology. Science 292, 1119-1122. http://sciencemag.org/content/292/5519/1119 [ Links ]
Smil, V., 2002. Worldwide transformation of diets, burdens of meat production and opportunities for novel food proteins. Enzyme Microb. Technol. 30, 305-311. https://doi.org/10.1016/S0141-0229(01)00504-X [ Links ]
Strydom, P.E. 2008. Do indigenous southern African cattle breeds have the right genetics for commercial production of quality meat? Meat Sci. 80, 86-93. DOI: 10.1016/j.meatsci.2008.04.017 [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fibre, neutral detergent fibre, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. DOI: 10.3168/jds.S0022-0302(91)78551-2 [ Links ]
Vermaak, H.J., 2011. Effect of dietary energy and fibre source on rumen function in feedlot steers. MSc dissertation, University of Pretoria, South Africa. https://repository.up.ac.za/bitstream/handle/2263/29555/dissertation.pdf?sequence=1 [ Links ]
Vohra, A., Syal, P. & Madan, A., 2016. Probiotic yeasts in livestock sector. Anim. Feed Sci. Technol. 219, 31-47. https://doi.org/10.1016/j.anifeedsci.2016.05.019 [ Links ]
West, M., Ellis, A.T., Potts, P.J., Streli, C., Vanhoof, C., Wegrznek, D. & Wobrausschek, P., 2013. Atomic spectrometry update - A review of advances in X-ray fluorescence spectrometry. J. Anal. At. Spectrom. 28, 1544-1590. https://pubs.rsc.org/en/content/articlelanding/2013/ja/c3ja90046k [ Links ]
Whitford, M.F., Teather, R.M. & Forster, R.J., 2001. Phylogenetic analysis of methanogens from the bovine rumen. BMC Microbiol. 1, 1-5. http://www.biomedcentral.com/1471-2180/1/5 [ Links ]
Wright, A.D.G., Williams, A.J., Winder, B., Christophersen, C.T., Rodgers, S.L. & Smith, K.D., 2004. Molecular diversity of rumen methanogens from sheep in Western Australia. Appl. Environ. Microbiol. 70(3), 1263-1270. https://aem.asm.org/content/70/3/1263 [ Links ]
Yan, T., Agnew, R.E., Gordon, F.J. & Porter, M.G., 2000. Prediction of methane energy output in dairy and beef cattle offered grass silage-based diets. Livestock Prod. Sci. 64, 253-263. DOI: 10.1016/S0301-6226(99)00145-1 [ Links ]
Submitted 19 July 2019
Accepted 11 June 2020
Published 13 January 2021
# Corresponding author: JiyanaS@arc.agric.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}