










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.50 n.5 Pretoria 2020
http://dx.doi.org/10.4314/sajas.v50i5.10
ARTICLE
SHORT COMMUNICATION
Hatch traits of artificially incubated ostrich eggs as affected by setting position, angle of rotation and season
Z. BrandI, #; S.W.P. CloeteII, III; C.R. BrownIV
IDirectorate Animal Sciences: Oudtshoorn, Western Cape Department of Agriculture, PO Box 351, Oudtshoorn, 6220, South Africa
IIDirectorate Animal Sciences: Elsenburg, Western Cape Department of Agriculture, P Bag X1, Elsenburg, 7607, South Africa
IIIDepartment of Animal Sciences, Stellenbosch University, P Bag X1, Matieland, 7602, South Africa
IVInstitute of Science and the Environment, University of Worcester, Henwick Grove, Worcester, WR2 6AJ, England
ABSTRACT
High levels of hatching failure in artificially incubated ostrich eggs cause considerable loss in income for the industry. During the 2015 - 2016 breeding seasons, between 846 and 1 549 egg records were used to determine the effect of various setting positions during artificial incubation. Fresh eggs were placed in trolleys to be turned automatically through a 60-degree or a 90-degree angle hourly in the setter. Additional treatments in a factorial design consisted of eggs set in the horizontal position for the total period of five weeks in the setter; eggs set horizontally for three weeks and vertically for two weeks; and eggs set vertically for five weeks in the setter. These treatments were repeated over two production years to represent various seasons, namely winter (June to August), spring (September to November) and summer (December). Late embryonic mortalities were improved significantly in eggs set in trolleys to turn through an angle of 90 degrees (0.16 ± 0.02) compared with eggs set in trolleys to turn through 60 degrees (0.28 ± 0.02), regardless of season and setting position. The preferred way of setting ostrich eggs would thus be in the vertical position in a trolley that turns hourly through an angle of 90 degrees with the air cell upwards to utilize incubator space optimally.
Keywords: chick weight, embryonic mortalities, moisture loss, ostrich, pipping time
Artificial incubation of ostrich eggs has become a major part of the ostrich industry. The success rate of artificially incubated ostrich eggs is low compared with commercially reared chickens (90 - 95%), turkeys (75 - 77%), and ducks (65 - 82%) (Hodgetts, 1990; Deeming & Ar, 1999), with hatching rates of only 50 -60% of eggs set (Brown et al., 1996; Deeming & Ar, 1999; Van Schalkwyk, 2000; Brand et al., 2007). This low hatching success represents a considerable loss of production and is cause for concern in the industry. More than 70% of all embryonic mortalities occur during the second half of incubation (Brand et al., 2007; 2012) and it is evident that there is still considerable room for improvement in this area.
Numerous factors affect the successful hatching of artificially incubated eggs, for example evaporative water loss, female age, season, storage conditions prior to setting, and genotype (Blood et al., 1998; Van Schalkwyk et al., 1999; Brand et al., 2007; 2008). However, almost all avian eggs need to be turned throughout incubation for correct embryonic development to take place (Tullett & Deeming, 1985; Deeming, 1991). More than 55% of shell deaths in ostrich eggs are because of malpositioning of the embryo (Brown et al., 1996). Although most embryos tend to adopt the correct pipping position (Deeming, 1995), Brand et al. (2017a) found that a substantial proportion of dead-in-shell chicks that pipped internally were positioned with their heads oriented towards the middle of the egg instead of towards the air cell. Ley et al. (1986) and Brand et al. (2017a) reported that embryos with pipping positions away from the air cell were more likely to succumb due to asphyxia as they would be unable to penetrate the air cell in the final stages of incubation.
Although hatchability causes a significant increase in late embryonic mortalities in chicken eggs, Badley (1997) reported that it was improved in ostrich eggs when set vertically and turned through a 180 degree angle along the long axis compared with those turned through a 90 degree angle. These results suggest that ostrich eggs respond differently to hatchery practices used for chicken eggs. Vertical hatching is not the normal position of eggs incubated naturally in ostrich nests (Wilson, 2003). Takeshita and McDaniel (1982) reported that early embryonic development was improved in eggs that were incubated horizontally. In contrast, Van Schalkwyk et al. (2000) found that the hatchability of fertile ostrich eggs was relatively low, but was similar when set in either the vertical or the horizontal position for six weeks. These contradictory findings indicate scope for a more detailed study. This study seeks to establish the effects of setting position on the overall hatchability of ostrich eggs.
Eggs were obtained from the commercial ostrich breeding flock at Oudtshoorn Research Farm in the Klein Karoo region of South Africa during the 2015 - 2016 breeding seasons. Details of the flock, management and feeding programme were described by Brand et al. (2007) and Cloete et al. (2008). The flock consisted of 155 breeding pairs, and the age of the breeders ranged between two and ten years. The flock consisted mostly of birds of the South African Black (SAB) genotype, but birds from the Zimbabwean Blue and Kenyan Redneck strains were introduced from 2003 to study crossbreeding of the SAB with these genotypes. The climate at the experimental site is arid with an average annual precipitation of 330 mm. The breeding season started mid-May and concluded in mid-December in each year.
After collection, the eggs were sterilized by exposure to ultraviolet light for 20 minutes, weighed, and the time, origin (paddock number) and date of collection were recorded. The eggs were divided at random into three groups with two stored in a horizontal position and the third stored vertically. All eggs were stored in their position at setting under controlled conditions (temperature of 17 °C; relative humidity (RH) of 75%) and rotated twice daily through 180 degrees until they were set. Eggs were set to turn automatically through a 60-degree or a 90-degree angle (i.e. 30 degrees and 45 degrees either side of the setting position, respectively) at hourly intervals. Turning was around the long axis when set horizontally and around the short axis when set vertically. The treatments per tray that represented each turning angle consisted of eggs set in the horizontal position for the incubation period of five weeks before being transferred to the hatcher; eggs set horizontally for three weeks and vertically for two weeks; and eggs set vertically for five weeks. These treatments were repeated over two years to include winter, spring and summer seasons.
Set eggs were candled and weighed on days 21 and 35 of incubation. Together with initial egg weight, these weights were used to derive moisture loss on day 21 of incubation (ML21) and day 35 of incubation (ML35). On day 35, the eggs were moved from the setters to a hatcher, which also operated at 36 °C and an RH of 24%. In the hatcher, the eggs set vertically in the setters were kept in a vertical position, while the eggs set horizontally were kept in a horizontal position. Eggs that did not show signs of continuing development at candling at 21 and 35 days (about 20% and 10% of eggs, respectively) were opened and assessed for early embryonic development or infertility (Brand et al., 2012). An egg with clear evidence of embryonic development that had subsequently ceased during the first 21 days of incubation was considered an early embryonic mortality (EEM). Subsequent embryonic mortality was classified as late embryonic mortality (LEM). Overall embryonic mortality (OEM) was the sum of EEM and LEM and included embryos that died during and after pipping (about 10% of eggs).
Embryonic mortality (defined as above) was calculated on a setting batch level as proportion of eggs set. Data for 846 to 1549 egg records at setting were used to derive averages for groups of eggs treated similarly. Setting batch data were subjected to a factorial analysis in a 2 (years) x 3 (seasons) x 3 (setting positions) x 2 (angles of rotation) design. Between 24 and 43 eggs represented each experimental unit in the analysis of variance (ANOVA). The LSD test was used to compare treatment means, provided that it was protected by a significant F-value in the ANOVA.
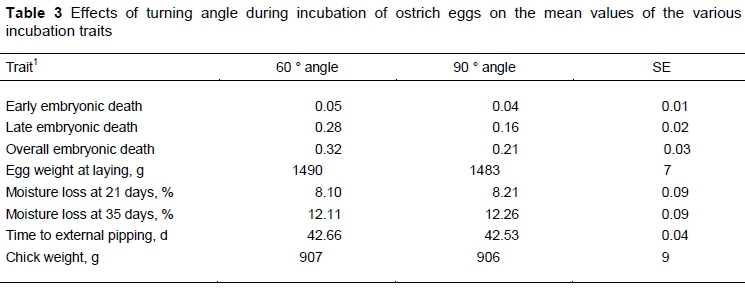
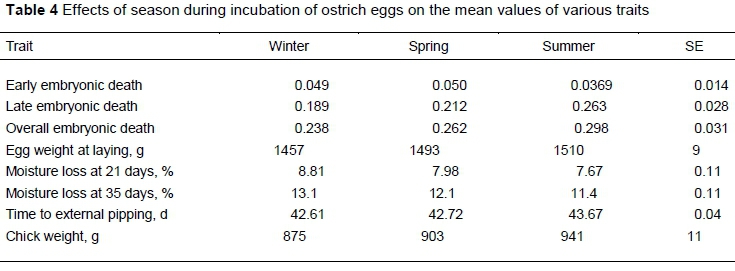
The significance of the fixed effects and their interactions are presented in Table 1. Late embryonic mortality and OEM were (P <0.01) affected by year, setting position and turning angle, whereas year and season affected moisture loss, pipping time and day-old-chick weight (P <0.01). Only main effects are presented in the absence of significant interactions between fixed effects for the various traits.
Moisture loss, pipping time and day-old-chick weight were largely independent of setting position and turning angle (Tables 2 & 3). The results from a previous study (Brand et al., 2012) are in broad agreement that the incubation position of eggs generally did not affect the measurements of the developing embryo throughout the 42-day incubation period. However, the findings in this study for the setting position are inconsistent with those of Van Schalkwyk et al. (2000), who reported improved hatchability of up to 36% in batches of eggs incubated for two or three weeks in the horizontal position and the rest of the incubation period in the vertical position. Conflicting results were also found in the literature for the best position of setting eggs of other species. Funk and Forward (1960) obtained better hatchability when chicken eggs were incubated vertically with the blunt end up as opposed to a horizontal setting, whereas Wilson (1991a; b) obtained significantly better hatchability for chickens and waterfowl when eggs were turned in a horizontal position rather than with the blunt end upwards. In this study, LEM was improved by ± 10% in eggs set vertically in comparison with eggs set horizontally for three weeks and vertically for two weeks. It was consistent with results of Brand et al. (2017b) that there was no difference in hatching results between eggs set horizontally and those set vertically.
There was an improvement of up to 50% in LEM for eggs set in trolleys that turned through an angle of 90 degree compared with eggs set in trolleys that turned through a 60 degree angle, regardless of season and setting position. The effect of turning angle on LEM was carried over to OEM. This was consistent with the findings of Van Schalkwyk et al. (2000) that the highest hatchability was at a turning angle of 90 degrees. A very low hatchability of 26.5% per eggs set was recorded in eggs that were not turned at all. The findings for other avian species suggested that failure to turn eggs results in retarded development of the area vasculosa and the extra-embryonic membranes, retarded embryonic growth, reduced oxygen uptake, and reduced albumen absorption (Tullett & Deeming, 1987; Deeming, 1989a; 1989b; 1989c). In contradiction, Brand et al. (2012; 2017b) found that the incubation position of eggs generally did not affect the measurements of the developing embryo throughout the 42-day incubation period, whereas the hatching of fertile eggs was independent of setting position.
Season did not affect EEM or LEM (Table 4). These results contradict the previous findings of Brand et al. (2007), who reported that chicks that hatched from eggs produced at the beginning of the breeding season, namely in winter, were more likely to succumb before hatching. This proportion decreased towards the end of the breeding season in summer. However, egg weight, moisture loss, pipping time and day-old-chick weight were affected by season, with moisture loss decreasing from winter to spring to summer. In contrast, pipping time became later, and day-old-chick weight became heavier from winter to autumn to summer. In the current study ML21 and ML35 decreased towards the summer, which could explain the later pipping time and increase in chick weight. Brand et al. (2008) reported that chick weight and pipping time in spring resembled winter values, but a subsequent decline took place towards summer. Moisture loss also increased from winter to summer. The anomaly in the results from the two studies could be attributed to the relatively small number of eggs used in the current trail, and to variations in seasonal climate. Ostrich nests are situated in open paddocks and the freshly laid eggs are subject to extreme weather fluctuations before they are collected and brought into the controlled environment at a hatchery.
Because ostrich farming is more extensive than most other avian farming enterprises, a better understanding of environmental impacts on the success of artificial hatching is important. Data from this study suggested that season affected moisture loss, pipping time, and day-old-chick weight more than setting position or angle of rotation. The preferred way of setting ostrich eggs would be in the vertical position in a trolley that turns hourly through an angle of 90 degrees with the air cell upwards to utilize incubator space optimally.
Acknowledgements
The authors are grateful to the Western Cape Department of Agriculture and Oudtshoorn Research Farm for the use of the resource flock and facilities. They thank I. Janse and U. Izaks for opening all the dead-in-shell eggs to record the data on the positions of the chicks for this study.
Authors' Contributions
Concept, design, data collection, drafting of the paper and submitting the manuscript, ZB. Critical analyses, SWPC. All the authors made substantial contributions to the original conception and design, acquisition of data, analyses and interpretation of data. All the authors have approved the manuscript being submitted.
Conflict of Interest Declaration
There are no conflicts of interest.
References
Badley, A.R., 1997. Fertility, hatchability and incubation of ostrich (Struthio camelus) eggs. Poult. Avian Biol. Rev. 8, 5376. [ Links ]
Blood, J.R., Van Schalkwyk, S.J., Cloete, S.W.P. & Brand, Z. 1998. Embryonic deaths in relation to moisture loss of artificially incubated ostrich eggs. Proc. 2nd Int. Ratite Conf. Oudtshoorn, South Africa. Pp. 148-151. [ Links ]
Brand, Z., Cloete, S.W.P., Brown, C.R. & Malecki, I.A., 2007. Factors related to shell-deaths during artificial incubation of ostrich eggs. J. S. Afr. Vet. Assoc. 78, 195-200. doi.org/10.4102/jsava.v78i4.322 [ Links ]
Brand, Z., Cloete, S.W.P., Brown, C.R. & Malecki, I.A., 2008. Systematic factors that affect ostrich egg incubation traits. S. Afr. J. Anim. Sci. 38(4), 315-325. [ Links ]
Brand, Z., Cloete, S.W.P., Malecki, I. & Brown, C., 2012. Heritability of embryonic mortalities in ostrich eggs and factors affecting hatching failure of fertile eggs during artificial incubation. Anim. Prod. Sci. 42, 806-812. doi.org/10.1071/AN11225 [ Links ]
Brand, Z., Cloete, S.W.P., Malecki, I. & Brown, C., 2017a. Dead-in-shell positions of near-term ostrich embryos. S. Afr. J. Anim. Sci. 47 (1), 2-6. doi.org/10.4314/sajas.v47i1.2 [ Links ]
Brand, Z., Cloete, S.W.P., Malecki, I. & Brown, C., 2017b. Ostrich (Struthio camelus) embryonic development from 7 to 42 days of incubation. Br. Poult. Sci. 58 (2), 139-143. doi.org/10.1080/00071668.2016.1259529 [ Links ]
Brown, C.R., Peinke, D. & Loveridge, A., 1996. Mortality in near-term ostrich embryos during artificial incubation. Br. Poult. Sci. 37, 73-85. doi.org/10.1080/00071669608417838 [ Links ]
Bunter, K.L. & Cloete, S.W.P., 2004. Genetic parameters for egg-, chick- and live-weight traits recorded in farmed ostriches (Struthio camelus). Livest. Prod. Sci. 91, 9-22. doi.org/10.1016/j.livprodsci.2004.06.008 [ Links ]
Cloete, S.W.P., Engelbrecht, A., Olivier, J.J. & Bunter, K.L. 2008. Deriving a preliminary breeding objective for commercial ostriches: An overview. Aust. J. Exp. Agric. 48, 1247-1256. doi.org/10.1071/EA08135 [ Links ]
Deeming, D.C., 1989a. Characteristics of unturned eggs, critical period, retarded embryonic growth and poor albumen utilisation. Br. Poult. Sci. 30, 253-263. doi.org/10.1080/00071668908417144 [ Links ]
Deeming, D.C., 1989b. Importance of sub-embryonic fluid and albumen in the embryo's response to turning of the egg during incubation. Br. Poult. Sci. 30, 591-606. doi.org/10.1080/00071668908417182 [ Links ]
Deeming, D.C., 1989c. Failure to turn eggs during incubation: Development of the area vasculosa and embryonic growth. J. Morph. 201, 179-186. doi.org/10.1002/jmor.1052010207 [ Links ]
Deeming, D.C., 1991. Reason for the dichotomy in egg turning in birds and reptiles. In: D.C. Deeming & M.W.J. Ferguson (eds). Egg incubation: Its effects on embryonic development in birds and reptiles. Cambridge University Press, Cambridge, UK. Pp 307-323. doi.org/10.1017/CBO9780511585739.020 [ Links ]
Deeming, D.C., 1995. Factors affecting hatchability during commercial incubation of ostrich (Struthio camelus) eggs. Br. Poult. Sci. 36, 51-65. doi.org/10.1080/00071669508417752 [ Links ]
Deeming, D.C. & Ar, A., 1999. In: D.C. Deeming (ed). The ostrich: Biology, production and health. CABI, Wallingford, Oxon, UK. Pp. 275-292. [ Links ]
Funk, E.M. & Forward, J.F., 1960. The relation of angle of turning and position of the egg to hatchability of chicken eggs. Poult. Sci. 39, 784-785. doi.org/10.3382/ps.0390784 [ Links ]
Hodgetts, B., 1990. Current hatchabilities in species of domestic importance and the scope for improvement. In S.G. Tullett (ed). Avian incubation. Butterworth-Heinemann, London. Pp. 139-144. [ Links ]
Ley, D.H., Morris, R.E., Smallwood, J.E. & Loomis, M.R. 1986. Mortality of chicks and decreased fertility and hatchability of eggs from a captive breeding pair of ostriches. J. Am. Vet. Med. Ass. 189, 1124-1126. [ Links ]
Pereira, D.F., Vitorasso, G., Oliveira, S.C., Kakimoto, S.K., Togashi, C.K.V. & Soares, N.M., 2008. Correlations between thermal environment and egg quality of two layer commercial strains. Brazilian J. Poultr. Sci. 81-88. doi.org/10.1590/S1516-635X2008000200002 [ Links ]
Snedecor, G.W. & Cochran, W.G. 1967. Statistical methods. 6th ed. Iowa State University, USA. [ Links ]
Takeshita, K. & McDaniel, G.R., 1982. Relationship of egg position during incubation on early embryonic growth and hatching of broiler breeder eggs. Poult. Sci. 61,667-672. [ Links ]
Tullett, S.G. & Deeming, D.C., 1987. Failure to turn eggs during incubation: Effects on embryo weight, development of the chorio-allantois and absorption of albumen. Br. Poult. Sci. 28, 239-249. doi.org/10.1080/00071668708416958 [ Links ]
Van Schalkwyk, S.J., Cloete, S.W.P., Brown, C. & Brand, Z., 1999. The effect of temperature on the hatching performance of ostrich eggs, and its implications for artificial incubation in forced draught wooden incubators. S. Afr. J. Anim. Sci. 29, 92-99. doi.org/10.4314/sajas.v29i2.44215 [ Links ]
Van Schalkwyk, S.J., Cloete, S.W.P., Brown, C. & Brand, Z., 2000. Hatching success of ostrich eggs in relation to setting, turning and angle of rotation. Br. Poult. Sci. 41, 46-52. doi.org/10.1080/00071660086394 [ Links ]
Wilson, H.R., 1991a. Physiological requirements of the developing embryo: Temperature and turning. In: S.G. Tullet (ed). Avian incubation. Butterworth-Heinemann, London. Pp. 145-156. [ Links ]
Wilson, H.R., 1991b. Interrelationship of egg size, chick size, posthatching growth and hatchability. World's Poult. Sci. J. 47, 5-20. doi.org/10.1079/WPS19910002 [ Links ]
Submitted 10 July 2018
Accepted 24 August 2020
Published 5 November 2020
# Corresponding author: Zanellb@elsenburg.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}