










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.50 no.3 Pretoria 2020
http://dx.doi.org/10.4314/sajas.v50i3.15
ARTICLES
Defatted yellow mealworm larvae (Tenebrio molitor l.) meal as possible alternative to fish meal in quail diets
S. SaricaI, #; B. KanogluI; U. YildirimII
IDepartment of Animal Science, Agricultural Faculty, Tokat Gaziosmanpasa University, Tokat 60240, Turkey
IIPoultry Husbandry Research Institute, Ankara 06170, Turkey
ABSTRACT
The effects of replacing fish meal (FM) in the diet with defatted mealworm larvae meal (DMLM) on growth, carcass, and biochemical constituents of serum of Japanese quail chicks were investigated. A total of 600 unsexed one-day-old chicks were allotted to five dietary treatments with six replicates of 20 chicks each. The diets were formulated by replacing FM with DMLM at the levels of 25% (D25), 50% (D50), 75% (D75), and 100% (D100). The FM, D25 and D50 diets resulted in greater final bodyweight, higher bodyweight gain, and improved feed conversion ratio compared with the other diets. Diets that contained DMLM decreased the feed intake compared with the FM diet. The FM and D25 diets increased the hot and cold carcass yields compared with the other diets, whereas the D75 and D100 diets significantly enhanced the relative weight of the small intestine and abdominal fat compared with FM, D25, and D50. The replacement of FM with DMLM reduced the serum globulin, total cholesterol and triglyceride levels, and the albumin to globulin ratio significantly. The best protein and energy efficiency ratios and European efficiency factor were obtained with the D25 and D50 diets. In conclusion, the replacement of FM with DMLM at the levels of 25% and 50% produced positive results in growth performance, in serum albumin to globulin ratio, and in nutrient use efficiency in quail.
Keywords: performance, serum biochemistry, slaughtering
Introduction
As a result of rapid growth, the population of the world is expected to be nine billion by 2050 (Cullere et al., 2016; Onsongo et al., 2018). Thus, the demand for poultry meat and eggs for human consumption will increase (Bahadori et al., 2017), resulting in a 58% rise in global meat demand compared with the 2010 level (Cullere et al., 2016). Therefore, increased poultry production is inevitable and would exacerbate the competition for conventional feedstuffs (Onsongo et al., 2018). Feedstuffs currently make up 60% - 70% of the total cost of production for broilers, with protein accounting for over 15% of the total (Khatun et al., 2005; Khan et al., 2017). Soybean meal (SBM) and FM are currently the major protein sources for broilers (Marono et al., 2015; Onsongo et al., 2018). As meat production increases, land that is available for soybean cultivation will decrease, and the price of SBM will rise (Onsongo et al., 2018).
Imbalances between essential and non-essential amino acids, anti-nutritional factors, and non-starch polysaccharides in SBM limit its use in broiler diets, although SBM protein has high digestibility (Sanchez-Muros et al., 2014). These problems are solved with dietary supplementation of synthetic amino acids, and the use of FM, which has high digestibility and a better balance of amino acids as a source of protein in broiler diets (Sanchez-Muros et al., 2014). The protein in FM has high biological value in broiler diets, because it is rich in essential amino acids, particularly sulphur-containing amino acids and lysine (Zadeh et al., 2019). But the deterioration of fisheries has reduced the availability of FM and increased its price (Sanchez-Muros et al., 2014). The inclusion of SBM and FM in broiler diets has become dependent on imports to an ever-greater degree, resulting in even higher costs and less profitable broiler production (Selaledi et al., 2019). Therefore, many researchers investigated alternative cheaper new protein source feedstuffs for SBM and FM that do not have adverse effects on broiler performance (Onsongo et al., 2018; Zadeh et al., 2019).
Insect meals have recently been regarded as an alternative source of protein for poultry because of the similarity of their crude protein and amino acid contents to FM (Bovera et al., 2015; Zadeh et al., 2019). Currently, raising insects as an alternative protein source in poultry diet originates from their use of organic waste as feedstuffs, thereby reducing environmental contamination and transforming low-quality plant waste into high quality protein. This insect-derived protein could replace increasingly expensive protein supplements such as FM (Veldkamp et al., 2012; Bovera et al., 2015; De Marco et al., 2015; Khan et al., 2017; Kovitvadhi et al., 2019). Several studies investigated the efficacy of silkworms (Bombyx mori L.) (Khatun et al., 2003; Ijaiya & Eko, 2009; Dutta et al., 2012), houseflies (Musca domestica L.) (Hwangbo et al., 2009), black soldier flies (Cullere et al., 2016; Onsongo et al., 2018), and mealworms (Tenebrio molitor L.) (Ballitoc & Sun, 2013; Biasato et al., 2016, 2017, 2018; Bovera et al., 2015, 2016; Hussain et al., 2017; Dabbou et al., 2019; Zadeh et al., 2019) as alternatives to SBM and FM in poultry diets.
Yellow mealworm, the larva of the mealworm beetle, is among the insect species that have widespread production in the world (Ramos-Elorduy et al., 2000). Mealworm has potential as a feedstuff for poultry because it contains high levels of protein, lipids, carbohydrates, and vitamins (Hussain et al., 2017). Larvae and pupa stages of yellow mealworm (T. molitor L.) are rich in protein, and the breeding and feeding of them is straightforward (Ghaly & Alkoaik, 2009). They grow well on organic waste, and are usually fed live to broilers, but may also be sold canned, dried or processed into a powder (Bovera et al., 2015). Yellow mealworm larvae meal (YMLM) is rich in crude protein (44% - 69%) and crude fat (23% - 47%) (Veldkamp et al., 2012). Extraction of oil from yellow mealworm larvae produces a meal that has only 3% - 7% crude fat on a dry matter (DM) basis (Veldkamp & Bosch, 2015). Important performance limiting essential amino acids in YMLM for growing poultry include methionine, methionine + cystine and arginine (Veldkamp et al., 2012).
Insects also contain chitin, a polysaccharide of the arthropod's exoskeleton (Henry et al., 2015). The chitin level affects the use of YMLM in poultry diet by producing a graded negative response in the digestibility of nutrients (Khempaka et al., 2011). Diets for broilers with 1.36% to 2.42% chitin in broiler diets had beneficial effects on growth without detrimental effects on nutrient digestibility, but a diet containing 3.23% chitin resulted in decreased digestibility of nutrients and performance. The chitin content of YMLM is a approximately 2.8% on a DM basis (Finke, 2013).
Chitin is not degraded and absorbed in the small intestine. Even so, chitin is fermented by the microbiota of the large intestine and acts as a prebiotic (Bovera et al., 2015; Loponte et al., 2017). Thus, chitin in YMLM is able to improve the sanitary status of broilers (Bovera et al., 2015) and laying hens (Marono et al., 2017). Chitin also has an antibacterial activity on gram negative pathogen bacteria (Bovera et al., 2015). Furthermore, feeding insects to chickens may reduce the need for dietary antibiotics (Bovera et al., 2015).
In the past, YMLM has replaced 25% (Schiavone et al., 2014) and 100% (Bovera et al., 2015, 2016; Khan et al., 2017) of SBM in broiler diets without negative effects on performance and serum biochemistry parameters. In addition, Loponte et al. (2017) reported that the inclusion of dietary YMLM meal as a replacement for SBM at the level of 25% in a Barbary partridge diet improved their growth performance and reduced their serum albumin to globulin ratio. Black soldier fly larvae meal has also been used to replace the conventional part of SBM and soybean oil in diets for growing broiler quail without detrimental effects (Cullere et al., 2016). However, no research has investigated the effects of replacing FM with DMLM in Japanese quail diets. Therefore, the aim of the present study was to investigate the effects of using various levels of DMLM to replace FM as a source of animal protein on growth performance, carcass traits and serum biochemistry parameters of Japanese quail.
Materials and Methods
A total of 600 one-day-old Japanese quail (Coturnix coturnix japonica) chicks were purchased from a commercial hatchery (19 Mayis University Poultry Production and Marketing Plant, Samsun, Turkey) for use in this experiment. The experimental procedures were approved by the Ethical Animal Care and Use Committee of Tokat Gaziosmanpasa University (Prot. no. 2017 HADYEK-03). The chicks were wing-banded, weighed and randomly assigned to five treatment groups, each with six replicates of 20 chicks (10 female, 10 male). From hatching until five weeks old, the chicks were kept under uniform environmental conditions in wire cages equipped with nipple drinkers. The temperature was kept at 32 °C for the first week, 28 °C for the second week, and then at 21 °C.
Yellow mealworm (T. molitor L.) larvae were purchased from a commercial supplier in Ankara, Turkey. These larvae had been grown on organic feed, which consisted mainly of wheat, wheat bran and carrots. The larvae were not starved before being killed and were freeze-dried overnight to remove moisture. Approximately 1 kg of freeze-dried mealworm larvae was ground into the meal. The fat was removed from the freeze-dried the freeze-dried larvae meal with a Soxhlet device under optimized extraction conditions with a petroleum ether to mealworm larvae ratio of 3 to 1 L/kg, at 60 °C for 4 hours. After removing the fat, the YMLM was dried at 40 °C for three hours. As a result of this process, the fat content of YMLM was reduced from 23% to 6.6% and its crude protein content was increased from 44% to 76.2%. The amount of protein linked to acid detergent fibre (ADF) was determined (AOAC, 2007) and used to estimate the amount of chitin in YMLM (Finke, 2007) as follows:

The chitin content of the YMLM was 4.2%.
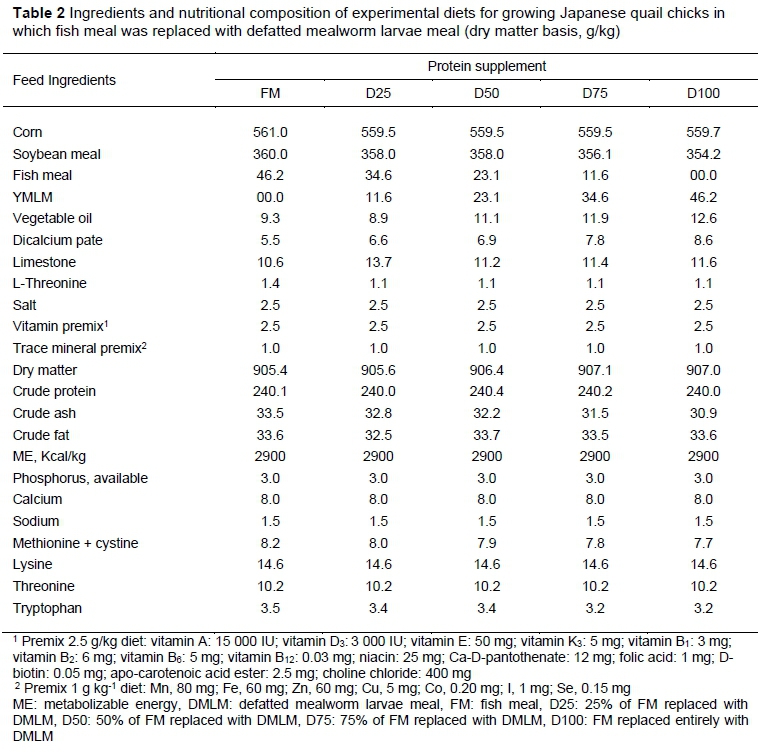
The dietary ingredients were ground through a 1-mm screen in preparation for chemical analysis. They were then analysed for dry matter, crude protein (CP), crude fat, crude ash, starch, total sugar, calcium, phosphorus and sodium contents according to the methods of the AOAC (2007) at Ankara Food Control Laboratory (Ankara, Turkey). Their essential amino acid contents were analysed by Turkish Scientific and Technical Research Institute (Gebze, Turkey) using high performance liquid chromatography. Their metabolizable energy (ME) content of each dietary ingredient was calculated based on its analysed nutrient composition (WPSA, 1989). The nutrient compositions of feed ingredients and the defatted YMLM (referred to hereafter as DMLM) are given in Table 1.
The experimental diets were formulated by replacing FM with DMLM at the levels of 0%, 25%, 50%, 75% and 100%. All diets were formulated to be isonitrogenous and isocaloric and meet the minimum nutrient requirements of the birds as established by the National Research Council (1994). The feed ingredients and calculated nutrient compositions of these diets are presented in Table 2. The diets were supplied in mash form and drinking water was provided ad libitum.
During the 35-day period, the growth performance of the quail was evaluated by recording the bodyweight gain (BWG), feed intake (FI) and feed conversion ratio (FCR). Bodyweight was recorded at the beginning of the experiment and then weekly. Feed supplied and leftovers were recorded weekly. The FI data were adjusted for mortalities. Mortality was recorded as it occurred. Feed conversion ratio was calculated weekly as the ratio of FI to BWG.
At 35 days old, 18 quail (9 female and 9 male) were selected from each group, because their BWs were similar to the group average, and leg banded and weighed after an eight-hour fasting period. An injection of sodium pentobarbital (100 mg/kg) was applied as anaesthesia to these quail before slaughtering.
A total of 90 quail were slaughtered by severing the jugular vein to determine the pre-slaughter BW, the relative weight of certain internal organs, and hot and cold carcass yields. When the quail were completely immobilized, their carcasses were scalded at 55 °C to 60 °C for 45 seconds and the feathers were removed in a rotary plucker. Immediately after being plucked, the carcasses were processed (removal of head and feet), eviscerated (removal of gastrointestinal tract), weighed and then chilled overnight in a refrigerator (4 °C). Data collected from the individual birds included the weights of the hot and cold carcasses, gizzard, liver, proventriculus, spleen, small intestine, and abdominal fat. The relative weights of the internal organs and hot and cold carcass yields were calculated as a percentage of the pre-slaughter BW.
Blood samples of the 90 slaughtered quail were collected in test tubes without anticoagulant and centrifuged at 3500 rpm for 15 min. at +4 °C. After centrifugation, the serum was removed and stored in Eppendorf tubes at -80 °C until analysis at the Biochemistry Laboratory of the Medicine Faculty of Tokat Gaziosmanpasa University with an auto-analyser using the test kits for total protein, albumin, globulin, uric acid, total cholesterol and triglyceride levels, as described by the manufacturers.
The protein efficiency ratio (PER), energy efficiency ratio (EER) and European efficiency factor (EEF) were calculated for 0-35 days:

One-way analysis of variance (SPSS, Inc., Cary, Illinois, USA) was applied to the data to determine the significance of the effects of the diet. Treatment means were separated with Duncan's multiple range test (Duncan, 1955). The results were presented at least square means and standard error. All statements of significance were based on P <0.05.
Results and Discussion
The diets supplemented with FM, D25 and D50 increased the final BW of quail compared with D75 and D100 (P <0.05) (Table 3). This result agrees with the findings of Pretorius (2011), for whom a 25% housefly larvae meal diet yielded significantly greater average live weights of broiler at slaughter compared with those that were fed a higher level of this meal. The current results for BW also concur with the findings of Dutta et al. (2012) and Jintasataporn (2012), who reported that the complete replacement of FM with silkworm pupae meal reduced the final BW of broilers significantly compared with a diet containing FM. Sapcota et al. (2003) also pointed out that replacing FM with silkworm at a level of 50% significantly decreased the final BW of broilers compared with a diet containing FM. However, Zadeh et al. (2019) reported that replacing 75% and 100% of dietary FM with YMLM significantly increased the final BW of Japanese quail compared with quail that were fed the basal diet (FM) and diets replaced with lesser amounts of YMLM.
The diets supplemented with FM, D25 and D50 increased BWG from 0 to 35 days compared with the D75 and D100 diets (P <0.05). This result agrees with the findings of Pretorius (2011) that a 25% housefly larvae meal diet yielded significantly greater average daily weight gain of broilers compared with a diet in which housefly larvae meal replaced 50% of the FM. But Awoniyi et al. (2003) reported that replacement of FM with maggot meal at the levels of 75 and 100% significantly reduced BWG of broilers compared with diets containing only FM and 25% maggot meal instead of FM. Moreover, Dutta et al. (2012) indicated that the dietary replacement of FM with silkworm pupae meal also significantly reduced the BWG of broilers. However, Agunbiade et al. (2007) reported that replacement of FM with increasing levels of maggot meal did not influence the daily BWG of layer hens significantly. Likewise, Zadeh et al. (2019) indicated that the use of YMLM to replace 75% and 100% of FM increased the BWG of the significantly quail compared with isocaloric diets containing larger amounts of FM. Moreover, Sapcota et al. (2003) pointed out that completely replacing FM with silkworm similarly increased the BWG of broilers compared with a diet containing FM. In addition, Biasato et al. (2017) and Hussain et al. (2017) reported that increasing dietary supplementation of YMLM produced a significant linear increase in the BWG of broilers.
The replacement of FM with increasing levels of DMLM decreased the FI of quail between 0 and 35 days old compared with the diet containing FM (P <0.05). These findings agree with those of Zadeh et al. (2019), who also reported that replacement of FM with 25%, 50%, 75%, and 100% YMLM significantly reduced the FI of quail. Further, the present results concur with those of Khatun et al. (2003), who reported that replacing FM with silkworm pupae meal decreased the FI significantly of broilers compared with the diet that contained only FM. However, Sapcota et al. (2003) and Dutta et al. (2012) reported that replacing FM with increasing amounts of silkworm meal did not significantly influence the FI of broilers compared with a diet containing only FM. Likewise, Biasato et al. (2017, 2018) and Hussain et al. (2017) indicated that dietary supplementation of YMLM at the increasing levels of 0, 50, 100, and 150 g/kg did not affect the FI of broilers.
The D25 and D50 diets improved the FCR of quail over the 35-day feeding period compared with the FM, D75 and D100 diets (P <0.05). Awoniyi et al. (2003) reported that replacement of FM with maggot meal at 75% and 100% levels did not affect the FCR of broilers compared with a diet using only FM. Likewise, Ijaiya and Eko (2009) pointed out that the FCR of broilers was not influenced by replacing 75% and 100% of dietary FM with silkworm caterpillar meal when compared with a diet that contained only FM. However, Jintasataporn (2012) reported that complete replacement of FM with silkworm pupa meal worsened the FCR of broilers compared with the diet containing FM. In contrast, Zadeh et al. (2019) showed that replacing FM with YMLM at the levels of 75 and 100% had better FCR values compared with the other diets.
As a whole, 75% and 100% replacement of FM with DMLM compromised the growth performance of quail. The D75 and D100 diets contained 3.15% and 4.2% chitin, respectively. The chitin bonded with the protein and amino acids, reducing their availability. Unfortunately, the digestibility of chitin by poultry is low, which reduces the availability of protein and organic matter and, as a result, degrades the growth performance of quail (Khatun et al., 2003; Ojewolia et al., 2005; Oduguwa et al., 2005; Jintasataporn, 2012; Khan et al., 2017).
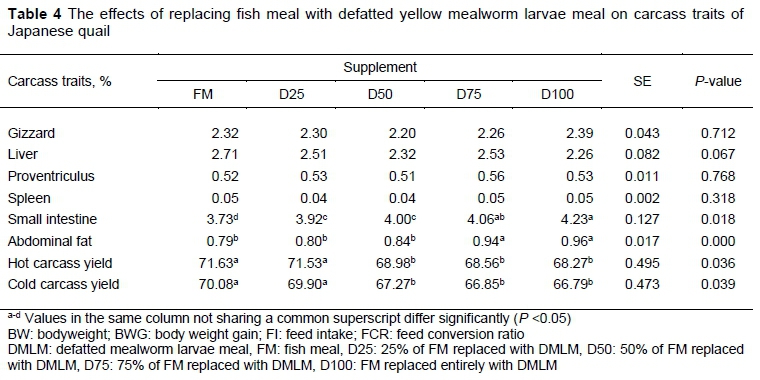
Replacement of FM with DMLM did not affect the relative weights of the gizzard, liver, proventriculus and spleen of the quail (P >0.05) (Table 4). The findings for the relative weights of the liver, gizzard and proventriculus are consistent with the results of Zadeh et al. (2019), who reported that replacing FM with YMLM at various levels did not influence the percentage weight of these organs. These results pertaining to the relative weight of liver concur with the findings of Bovera et al. (2016), who reported that the complete replacement of SBM with YWLM did not significantly affect the relative weight of the liver of broilers. Similarly, Khatun et al. (2003) and Biasato et al. (2017) also pointed out that the percentage weights of the liver and gizzard were not affected by various levels of YMLM in the diets of broiler chickens. In contrast, Ballitoc and Sun (2013) reported that the supplementation of YMLM to broiler diet increased the relative weight of gizzards compared with the control. Complete replacement of SBM with YWLM has also been shown to increase significantly in the relative weight of the spleen in broilers (Bovera et al., 2016)
The D75 and D100 diets increased the relative weight of the small intestine compared with FM, D25 and D50 (P <0.05). The D75 (3.15% chitin), and D100 (4.2% chitin) diets led to the consumption of high levels of chitin compared with FM (0% chitin), D25 (1.05% chitin), and D50 (2.10% chitin). Increased chitin consumption reduces protein digestibility and a low digestible level of diet forces the development of an increase in surface area of the small intestine for the absorption of nutrients in quail. Thus, the weight and length of the small intestine are increased (Borin et al., 2006; Bovera et al., 2015).
The D75 and D100 diets increased the relative weight of abdominal fat compared with the FM, D25, and D50 diets (P <0.001). Jintasataporn (2012) also reported that the complete replacement of FM with silkworm pupa meal significantly increased the relative weight of the abdominal fat of broilers compared with a diet that contained FM. But the low nutrient digestibility because of the high chitin level in D75 and D100 diets may cause problems in the absorption of amino acids in chickens and result in an accumulation of abdominal fat (Jintasataporn, 2012).
The FM and D25 diets increased the hot and cold carcass yields of quail compared with D50, D75 and D100 (P <0.05) (Table 4). In the D25 diet, the level of chitin (1.05%) that was consumed was not high enough to reduce protein digestibility. This probably caused an increased rate of protein accumulation and carcass yield (Hwangbo et al., 2009). However, Khatun et al. (2003) reported that replacing dietary FM with silkworm pupae at the levels of 50%, 75%, and 100% increased the carcass yields of broilers significantly compared with the FM diet. Furthermore, Zadeh et al. (2019) pointed out that 100% replacement of FM with YMLM similarly affected the carcass yield of quail compared with the FM diet.
The FM and D25 diets significantly increased the serum total protein and uric acid levels of quail compared with the other diets (Table 5). Bovera et al. (2015) also reported that the total replacement of SBM with YMLM significantly decreased the serum uric acid levels of broilers compared with a SBM-based diet. Serum albumin levels of quail were also significantly reduced by supplementation with increasing DMLM levels (P <0.05). The present results are consistent with those of Biasato et al. (2017) and Loponte et al. (2017), who reported that the serum albumin levels of broilers and Barbary partridge, respectively, were reduced significantly with the increasing levels of TMLM. In contrast, Bahadori et al. (2017) showed that the inclusion of higher levels of earthworm in the diet increased serum total protein and albumin concentrations.
Uric acid is the major waste product of nitrogen metabolism in poultry. The serum uric acid levels of quail fed the D50, D75, and D100 diets decreased compared with the FM and D25 diets (P <0.05). These findings concur with the results of Bovera et al. (2015) and Loponte et al. (2017), who pointed out that replacing 50% to 100% of the dietary SBM with YMLM decreased serum uric acid levels in broilers and Barbary partridges, respectively. There is a direct relationship between the amount of digested protein and serum uric acid concentration. Therefore, changes in protein metabolism are reflected in serum uric acid concentration. The present serum albumin and uric acid results may be ascribed to the properties and levels of chitin in DMLM (Bovera et al., 2015). This might be because of the reduced digestibility of protein with higher levels of chitin, as in the D50, D75 and D100 diets, because chitin bonded with the protein and reduced its digestibility. This phenomenon would result in the levels of serum albumin and uric acid and the concentrations of metabolites of protein degradation being reduced (Loponte et al., 2017).
The D25, D50, D75, and D100 diets decreased the serum globulin level (P <0.001) and the albumin to globulin ratio (P <0.05) of quail compared with the FM diet. Bovera et al. (2015) and Loponte et al. (2017) also observed that the albumin to globulin ratio decreased when broilers were fed YMLM. Griminger and Scanes (1986) and Biasato et al. (2018) stated that a low albumin to globulin ratio indicated enhanced disease resistance and immune response of birds. This result might be derived from the properties of the chitin content of insect meal. Chitin can be fermented only by the microbiota of the large intestine, acting as a prebiotic (Loponte et al., 2016). The prebiotic effect of chitin results from the higher production of butyric acid in the caecal content of broilers fed T. molitor (Loponte et al. (2016). Thus, chitin can reduce the prevalence and duration of some infectious agents and improve the health status of the digestive system and the immune response of the birds (Choi et al., 2018; Selaledi et al., 2019). In addition, chitin exhibits a bacteriostatic effect on gram negative bacteria (Vidanarachchi et al., 2010) and has antimicrobial and antifungal properties (Khoushab & Yamabhai, 2010). The use of insect meal in the diets of poultry under intensive farm conditions may decrease the use of antibiotics. However, Biasato et al. (2018) found that increasing dietary YMLM did not influence the serum total protein and uric acid levels of broilers.
The D25, D50, D75, and D100 diets decreased the serum cholesterol and triglyceride levels of quail compared with birds fed the FM diet (P <0.05). Chitin in DMLM binds with anionic carboxyl groups of fat and bile acids in the intestine and interferes with the emulsification with neutral lipids by binding them with hydrophobic bonds, and blocks the absorption of cholesterol and lipid. Moreover, chitin prevents the reabsorption of biliary cholesterol, which increases the excretion of faecal cholesterol and lipids. As a result, serum cholesterol and lipid levels are decreased (Simunek & Bartonova, 2005). These findings do not agree with the results of Bovera et al. (2015), Biasato et al. (2017, 2018) and Loponte et al. (2017), who reported that YMLM inclusion in their diets did not influence the serum total cholesterol and triglyceride levels of broilers significantly.
The D25 and D50 groups had significantly increased protein and energy efficiency ratios of compared birds fed the other diets (Table 6). In contrast, it was reported that the replacement of FM with silkworm caterpillar meal in broiler diets did not influence their PER (Ijaiya & Eko, 2009). Moreover, the European efficiency factor (EEF) was increased by the D25 diet compared with the other diets (P <0.05). In addition, the D50 diet increased the EEF of the experimental groups compared with the FM, D75, and D100 diets (P <0.05). High EEF shows that the D25 and D50 diets had a positive effect on the growth performance of broilers compared with the FM, D75 and D100 diets, because EEF value is related to growth performance. Moreover, a reduction in the EEF value indicated that production efficiency had worsened owing to fat deposition (Bovera et al., 2015).
Conclusion
As a source of protein in the diet of Japanese quail, 25% to 50% of FM could be replaced with DMLM without compromising growth performance, serum albumin to globulin ratio, cholesterol and triglyceride levels, or production efficiency. By acting as a prebiotic, the chitin in the DMLM may also decrease the use of antibiotics.
Acknowledgments
This work was supported financially by the authors.
Authors' Contributions
The authors developed the original hypotheses, designed the experiments, collected the data, conducted the statistical analyses, collaborated in interpretation of the results, wrote the initial draft of this manuscript and finalized the manuscript. All authors have read and approved the finalized manuscript.
Conflict of Interest Declaration
There is no conflict of interest.
References
Agunbiade, J.A., Adeyemi, O.A., Ashiru, O.M., Awojobi, H.A., Taiwo, A.A., Oke, D.B. & Adekunmisi, A.A., 2007. Replacement of fish meal with maggot meal in cassava based layers'diets. J. Poult. Sci. 44, 278-282. [ Links ]
AOAC, 2007. Official methods of analysis (18th ed.). Association of Official Analytical Chemists. Washington DC, USA. [ Links ]
Awoniyi, T.A.M., Alletor, V.A. & Aina, J.M., 2003. Performance of broiler chickens fed on maggot meal in place of fish meal. Int. J. Poult. Sci. 2 (4), 271-274. [ Links ]
Bahadori, Z., Esmaielzadeh, L., Karimi-Torshizi, M.A., Seidavi, A., Olivares, J., Rojas, S., Salem, A.Z.M., Khusro, A. & Lopez, S., 2017. The effect of earthworm (Eisenia foetida) meal with vermi-humus on growth performance, hematology, immunity, intestinal microbiota, carcass characteristics and meat quality of broiler chickens. Lives. Sci. 202, 74-81. [ Links ]
Ballitoc, D.A. & Sun, S., 2013. Ground yellow mealworms (Tenebrio molitor L.) feed supplementation improves growth performance and carcass yield characteristics in broilers. Open Sci. Repos. Agric. e23050425. DOI: 10.7392/openaccess.23050425 [ Links ]
Biasato, I., De Marco, M., Rotolo, L., Renna, M., Dabbou, S., Capucchio, M.T., Biasibetti, E., Tarantola, M., Costa, P., Gai, F., Pozzo, L., Dezzutto, D., Bergagna S., Gasco, L. & Schiavone, A., 2016. Effects of dietary Tenebrio molitor meal inclusion in free-range chickens. J. Anim. Physiol. Anim. Nutr. 100, 1104-1112. [ Links ]
Biasato, I., Gasco, L., De Marco, M., Renna, M., Rotolo, L., Dabbou, S., Capucchio, M.T., Biasibetti, F., Tarantola, M., Bianchi, C., Cavallarin, L., Gai, F. Pozzo, L., Dezzutto, D., Bergagna, S. & Schiavone, A., 2017. Effects of yellow mealworm larvae (Tenebrio molitor) inclusion in diets for female broiler chickens: implications for animal health and gut histology. Anim. Feed Sci. Technol. 234, 253-263. [ Links ]
Biasato, I., Gasco, L., De Marco, M., Renna, M., Rotolo, L., Dabbou, S., Capucchio, M.T., Biasibetti, E., Tarantola, M., Sterpone, L., Cavallarin, L., Gai, F., Pozzo, L., Bergagna, S., Dezzutto, D., Zoccarato, I. & Schiavone, A., 2018. Yellow mealworm larvae (Tenebrio molitor L.) inclusion in diets for male broiler chickens: Effects on growth performance, gut morphology and histological findings. Poult. Sci. 97, 540-548. [ Links ]
Borin, K., Lindberg, J.E. & Ogle, R.B., 2006. Digestibility and digestive organ development in indigenous and improved chickens and ducks fed diets with increasing inclusion levels of cassava leaf meal. J. Anim. Physiol. Anim. Nutr. 90 (5-6), 230-237. [ Links ]
Bovera, F., Piccolo, G., Gasco, L., Marono, S., Loponte, R., Vassalotti, G., Mastellone, V., Lombardi, P., Attia, Y. A. & Nizza, A., 2015. Yellow mealworms larvae (Tenebrio molitor L.) as protein source for broilers: Effects on growth performance and blood profiles. Br. Poult. Sci. 56, 569-575. [ Links ]
Bovera, F., Loponte, R., Marono, S., Piccolo, G., Parisi, G., Laconisi, V., Gasco, L. & Nizza, A., 2016. Use of Tenebrio molitor larvae meal as protein source in digestibility and carcass and meat traits. J. Anim. Sci. 94, 639-647. [ Links ]
Choi, I.H., Kim, J.M., Kim, J.D., Park, C., Park, J.H. & Chung, T.H., 2018. Replacing fish meal by mealworm (Tenebrio molitor) on the growth performance and immunological responses of white shrimp (Litopenaeus vannamei). Acta Sci. Anim. Sci. 40, 1-9. [ Links ]
Cullere, M., Tasoniero, G., Giaccone, V., Miotti-Scapin, R., Claeys, E., De Smet, S., & Dalle Zotte, A., 2016. Black soldier fly as dietary protein source for broiler quails: Apparent digestibility, excreta microbial load, feed choice performance, carcass and meat traits. Anim. 10, 1923-1930. [ Links ]
Dabbou, S., Gasco, L., Lussiana, C., Brugiapaglia, A., Biasato, I., Renne, M., Cavallarin, L., Gai, F. & Schiavone, A., 2019. Yellow mealworm (Tenebrio molitor L.) larvae inclusion in diets for free-range chickens: Effects on meat quality and fatty acid profile. Renew. Agric. Food Syst. 1-8. https://doi.org/10.1017/S1742170519000206 [ Links ]
De Marco, M., Martinez, S., Hernandez, F., Madrid, J., Gal, F., Rotolo, L., Belforti, M., Bergero, D., Katz, H., Dabbou, S., Kovitvadhi, A., Zoccarato, I., Gasco, L. & Schiavone, A., 2015. Nutritional value of two insect meal (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent nutrient digestibility, apparent ileal amino acid digestibility and apparent metabolizable energy. Anim. Feed Sci. Technol. 209, 211-218. [ Links ]
De Marcu, A., Vacaru-Opriç, I., Dumitrescu, G., Petculescu Ciochina, L., Marcu, A., Nicula, M., Pet, I., Dronca, D., Kelciov, B. & Mariç, C., 2013. The influence of genetics on economic efficiency of broiler chickens' growth. J. Anim. Sci. Biotechnol. 46, 339-346. [ Links ]
Doskovic, V., Bogosavljevic-Boskovic, S., Pavlovski, Z., Milosevic, B., Skrbic, Z., Radonjac, S. & Petricevic, V., 2012. The effect of protease on productive and slaughter traits in broiler chickens. Biotechnol. in Anim. Husb. 28 (4), 817-826. [ Links ]
Duncan, D.B., 1955. Multiple range and multiple F tests. Biometrics 11, 1-42. [ Links ]
Dutta, A., Dutta, S. & Kumari, S., 2012. Growth of poultry chicks fed on formulated feed containing silkworm pupae meal as protein supplement and commercial diet. Online J. Anim. Feed Res. 2 (3), 303-307. [ Links ]
Finke, M.D., 2007. Estimate of chitin in raw whole insects. Zoo Biol. 26, 105-115. [ Links ]
Finke, M.D., 2013. Complete nutrient content of four species of feeder insects. Zoo Biol. 32, 27-36. [ Links ]
Ghaly, A.E. & Alkoaik, F.N., 2009. The yellow mealworm as a novel source of protein. Am. J. Agric. Biol. Sci. 4, 319-331. [ Links ]
Griminger, P. & Scanes, C.G., 1986. Protein metabolism. In: P.D. Sturkie (ed). Avian physiology. 4th ed. Springer Verlag, New York. Pp. 326-345. [ Links ]
Henry, M., Gasco, L., Piccolo, G. & Fountoulaki, E., 2015. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 203,1-22. [ Links ]
Hussain, I., Khan, S., Sultan, A., Chand, N., Khan, R., Alam, W. & Ahmad, N., 2017. Meal worm (Tenebrio molitor) as potential alternative source of protein supplementation in broiler. Int. J. Biosci. 10 (4), 225-262. [ Links ]
Hwangbo, J., Hong, E.C., Jang, A., Kang, H.K., Oh, J.S., Kim, B.W. & Park, B.S., 2009. Utilization of house fly maggots, a feed supplement in the production of broiler chickens. J. Environ. Biol. 30 (4), 609-614. [ Links ]
Ijaiya, A.T. & Eko, E.O., 2009. Effect of replacing dietary fish meal with silkworm (Anaphe infracta) caterpillar meal on growth, digestibility and economics of production of starter broiler chickens. Pakistan J. Nutr. 8 (6), 845-849. [ Links ]
Jintasataporn, O., 2012. Production performance of broiler chickens fed with silkworm pupa (Bombyx mori). J. Agric. Sci. Technol. ISSN 1939-1250. [ Links ]
Kamran, Z., Sarwar, M., Nisa, M., Nadeem, M.A., Mahmood, S., Babar, M.E. & Ahmed, S., 2008. Effect of low protein diets having constant energy to protein ratio on performance and carcass characteristics of broiler chickens from one to thirty-five days of age. Poult. Sci. 87, 468-474. [ Links ]
Karimi, A., 2006. The effects of varying fish meal inclusion levels (%) on performance of broiler chicks. Int. J. Poult. Sci. 5, 255-258. [ Links ]
Khan, S., Naz, S., Sultan, A., Alhidary, I.A., Abdelrahman, M.M., Khan, R.U., Khan, N.A., Khan, M.A. & Ahmad, S., 2016. Worm meal: A potential source of alternative protein in poultry feed. World's Poult. Sci. J. 72, 93-102. [ Links ]
Khan, S., Khan, R.U., Alam, W. & Sultan, A., 2017. Evaluating the nutritive profile of three insect meals and their effects to replace soyabean in broiler diet. J. Anim. Physiol. Anim. Nutr. 102, 662-668. [ Links ]
Khatun, R., Howlider, M.A.R., Rahman, M.M. & Hasanuzzaman, M., 2003. Replacement of fish meal by silkworm pupae in broiler diets. Pakistan J. Biol. Sci. 6 (11), 955-958. [ Links ]
Khatun, R., Azmal, S.A., Sarker, M.S.K., Rashid, M.A., Hussain, M.A. & Miah, M.Y., 2005. Effect of silkworm pupae on the growth and egg production performance of Rhade Island Red (RIR) pure line. Int. J. Poult. Sci. 4 (9), 718-720. [ Links ]
Khempaka, S., Mochizuki, M., Koh, K. & Karasawa, Y., 2006. Effect of chitin in shrimp meal on growth performance and digestibility in growing broilers. J. Poult. Sci. 43, 339-343. [ Links ]
Khempaka, S., Chitsatchapong, C. & Molee, W., 2011. Effect of chitin and protein constituents in shrimp head meal on growth performance, nutrient digestibility, intestinal microbial populations, volatile fatty acids and ammonia production in broilers. J. Appl. Poult. Res. 20, 1-11. [ Links ]
Khoushab, F. & Yamabhai, M., 2010. Chitin research revisited. Mar. Drugs 8, 1988-2012. [ Links ]
Kovitvadhi, A., Chundang, P., Thongprajukaew, K., Tinawattanawanich, C., Srikachar, S. & Chotimaothum, B., 2019. Potential of insect meals as protein sources for meat-type ducks based on in vitro digestibility. Anim. 9 (155), 110. [ Links ]
Loponte, R., Marono, S., Laconisi, V., Piccolo, G., Parisi, G. & Bovera, F., 2016. Caecal volatile fatty acid production of broilers fed Tenebrio molitor larvae meal. In: INSECTA 2016 International Symposium on Insects as Food, Feed and Non-Food. Magdeburg, Germany, 12 September 2016. [ Links ]
Loponte, R., Nizza, S., Bovera, F., De Riu, Nicola, Fliegerova, K., Lombardi, P., Vassalotti, G., Mastellone, V., Nizaa, A. & Moniello, G., 2017. Growth performance, blood profiles and carcass traits of Barbary partridge (Alectoris barbara) fed two different insect larvae meals (Tenebrio molitor and Hermetia illucens). Res. Vet. Sci. 115, 183188. [ Links ]
Marono, S., Piccolo, G., Loponte, R., Di Meo, C., Attia, Y.A., Nizza, A. & Bovera, F., 2015. In vitro crude protein digestibility of Tenebrio molitor and Hermetia illucens insect meals and its correlation with chemical composition traits. Italian J. Anim. Sci. 14, 338-343. [ Links ]
NRC, 1994. Nutrient requirements of poultry. 9th ed. National Academy Press, Washington DC. [ Links ]
Oduguwa, O.O., Fanimo, A.O. & Oso, M.J., 2005. Effect of replacing dietary fish meal or soyabean with shrimp waste meal on the performance of laying hens. Nigerian J. Anim. Prod. 32, 224-232. [ Links ]
Ojewola, G.S., Okoye, F.C. & Ukoha, O.A., 2005. Comparative utilization of three animal protein sources by broiler chickens. Int. J. Poult. Sci. 4970, 462-467. [ Links ]
Onsongo, V.O., Osuga, I.M., Gachuri, C.K., Wachira, A.M., Miano, D.M. & Tanga, C.M., 2018. Effect of dietary replacement of soybean and fish meal with black soldier fly meal on broiler growth and economic performance. J. Econ. Entomol. 111, 1966-1973. [ Links ]
Pretorius, Q., 2011. The evaluation of larvae of Musca Domestica (common housefly) as protein source for broiler production. Stellenbosch University, MSc thesis, Stellenbosch, South Africa. [ Links ]
Ramos-Elorduy, J., Gonzalez, E.A., Hernandez, A.R. & Pino, J.M., 2002. Use of Tenebrio molitor (Coleoptera:Tenebrionidae) to recycle organic wastes and as feed for broiler chickens. J. Econ. Entomol. 95 (1), 214-220. [ Links ]
Sanchéz-Muros, M.J., Barrosa, F.G. & Manzano-Agugliaro, F., 2014. Insect meal as renewable source of food for animal feeding: A review. J. Clean. Prod. 65, 16-27. 2014. [ Links ]
Sapcota, D., Sheikh, I.U., Dutta, K.K., Sarma, S. & Ranjana, G., 2003. Effect of dietary muga silkworm supplementation on the performance of broilers. Indian Vet. J. 80, 19-22. [ Links ]
Selaledi, L., Mbajiorgu, C.A. & Mabelebele, M., 2019. The use of yellow mealworm (T. molitor) as alternative source of protein in poultry diets: a review. Trop. Anim. Health Prod. https://doi.org/10.1007/s11250-019-02033-7 [ Links ]
Schiavone, A., De Marco, M., Rotolo, L., Belforti, M., Martinez Miro, S., Madrid Sanchez, J., Hernandez Ruiperaz, F., Bianchi, C., Sterpone, L., Malfatto, V., Katz, H., Zoccarato, I., Gai, F. & Gasco, L., 2014. Nutrient digestibility of Hermetia illucens and Tenebrio molitor meal in broiler chickens. In. Proc. 1st Int. Confer. Insects to Feed the World, Wageningen, The Netherlands, 73. [ Links ]
Simunek, J. & Bartonova, H., 2005. Effect of dietary chitin and chitosan on cholesterolemia of rats. Acta Vet. Brno 74, 491-499. [ Links ]
SPSSWIN 2007. SPSS for Windows 6.1.4. SPSSWIN, Istanbul, Turkey. [ Links ]
Veldkamp, T. & Bosch, G., 2015. Insects: A protein-rich feed ingredient in pig and poultry diets. Anim. Frontier 5, 45-50. [ Links ]
Veldkamp, T., Van Duinkerken, G., Van Huis, A., Lakemand, C.M.M., Ottevanger, E., Bosch, G. & Van Boekel, M.A.J.S., 2012. Insects as a sustainable feed ingredient in pig and poultry diets - a feasibility study. Report 638, Wageningen Livestock Res. [ Links ]
Vidanarachchi, J.K., Kurukulasuriya, M.S. & Kim, S.K., 2010. Chitin, chitosan and their oligosaccharides in food industry. In: S.K. Kim, (ed). Chitin, chitosan, oligosaccharides and their derivatives: Biological activities and applications. CRC, New York, USA. Pp. 543-560. [ Links ]
WPSA,1989. European Table of Energy Values for Poultry Feedstuffs (3rd ed.). WPSA sub-Committee, Beekbergen, The Netherlands. [ Links ]
Zadeh, Z.S., Kheiri, F. & Faghani, M., 2019. Use of yellow mealworm (Tenebrio molitor) as a protein source on growth performance, carcass traits, meat quality and intestinal morphology of Japanese quails (Coturnix japonica). Vet. Anim. Sci. 8, 1-5. [ Links ]
Submitted 18 October 2019
Accepted 6 May 2020
First published online 2 August 2020
# Corresponding author: senay.sarica@gop.edu.tr


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}