










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.50 no.3 Pretoria 2020
http://dx.doi.org/10.4314/sajas.v50i3.7
Incorporating oldman saltbush hay and prickly pear in diets for red Sindhi calves
J.N. AlvesI; G.G.L. AraújoII; L.G.R. PereiraIII; S. Gonzaga NetoI; D.R. MenezesIV; T.V. VoltoliniII; G.C. GoisIV, #; L.C. SouzaI; F.S. CamposV
IFederal University of Paraíba, Animal Production Department, Areia - PB, Brazil
IIBrazilian Agricultural Research Corporation - Embrapa Semiárido, Petrolina - PE, Brazil
IIIBrazilian Agricultural Research Corporation, Juiz de Fora - MG, Brazil
IVPostgraduate Program in Veterinary Sciences in the Semiarid, Federal University of San Francisco Valley, Petrolina - PE, Brazil
VAcademic Unit of Garanhuns, Federal Rural University of Pernambuco, Garanhuns - PE, Brazil
ABSTRACT
This study was designed to evaluate the nutrient intake, digestibility, degradability, and ruminal characteristics of Sindhi heifers fed diets that contained a combined total of 75% oldman saltbush hay (hereafter saltbush hay) and prickly pear cactus. Eight 12-month old intact male red Sindhi calves (four fistulated and four non-fistulated) with an initial mean weight of 170 ± 5 kg were assigned to 4 * 4 Latin squares, where factors consisted of four diets, namely 15% hay and 60% cactus; 30% hay and 45% cactus; 45% hay and 30% cactus; and 60% hay and 15% cactus, and four times at which rumen fluid was collected. Neutral detergent fibre (NDF) and acid detergent fibre (ADF) intakes in kg/day and NDF in percentage live weight (LW), water intake, salinity, and conductivity increased with hay level. Intake and digestibility of non-fibrous carbohydrates were curvilinear with higher values in diets containing 30% saltbush hay. The apparent digestibility of dry matter (DM) and organic matter decreased linearly as the concentration of saltbush hay increased in the diet. The pH of the rumen fluid was within the acceptable range for favourable microbial growth. The low temperature and high salinity and conductivity indices in the diets should be viewed with caution at higher concentrations of saltbush hay, because of a possible decrease in nutrient absorption and the development of health problems in the animals. Apparent degradability coefficients of DM and NDF were affected significantly by inclusion of prickly pear and saltbush hay in the diets.
Keywords: animal nutrition, apparent digestibility, Atriplex nummularia Lindl, Opuntia ficus, ruminal degradation, semi-arid environment
Introduction
The market for animal products encourages production systems to focus on efficiency and final product quality to minimize seasonality (Alao et al., 2017). Although Brazil is rich in natural resources, cattle production systems in the semi-arid region show low yields from continuous exposure of animals to stressful conditions. Such poor production is associated with animal nutritional deficiencies, the use of exotic animal breeds that are unsuitable for the local climate, the high cost of supplemental fodders, and low forage availability (Alves et al., 2017).
In the semi-arid region, several food sources can be used as ruminant feed. However, the nutritional value and quality of these sources are determined by a complex interaction between ingested nutrients and microbial action in the digestive tract, which involves digestion, transport, metabolite uses, and animal physiological status (Nunes et al., 2015, Celi et al., 2017).
Oldman saltbush (Atriplex nummularia Lindl) has been used in several arid and semi-arid regions worldwide as fodder to supplement ruminant diets. This species is a halophyte, and its fodder has a high salt concentration (Helal et al., 2018). However, it is necessary to evaluate the potential of halophytes as fodder in association with other plants that are adapted to Brazilian semi-arid conditions (Norman et al., 2012). For example, prickly pear (Opuntia ficus) can be used with other halophyte species since it is a succulent carbohydrate-rich food source that is critical to cattle production, especially during prolonged drought when it can meet most of the water needs of animals (Silva et al., 2017).
The objective of this study was to evaluate nutrient intake, digestibility, degradability, and ruminal characteristics of the feed by Sindhi calves fed diets containing saltbush hay and prickly pear.
Materials and Methods
The experiment was conducted in the Animal Metabolism Unit at Caatinga Experimental Station of Embrapa Semiárido, Petrolina, Pernambuco, Brazil (09°09' S, 40°22' W, 365 m). All the animals were cared for in accordance with the guidelines of the National Council for the Control of Animal Experimentation (CONCEA, 2008). The mean annual rainfall in the study region is 570 mm, with mean annual maximum and minimum temperatures of 33.8 °C and 21.5 °C (EMBRAPA, 2014).
Eight Sindhi male calves (four fistulated and four non-fistulated), with an initial mean weight of 170 ± 5 kg and a mean age of 12 months, were distributed in a double 4 x 4 Latin square, where factors consisted of four dietary treatments (Table 1) and times at which rumen fluid was collected. Before starting the study, animals were weighed, identified with numbered ear tags, treated for endo- and ecto-parasites, and assigned to individual pens (1.00 * 1.60 m) with feeders, water dispensers, and salt troughs.
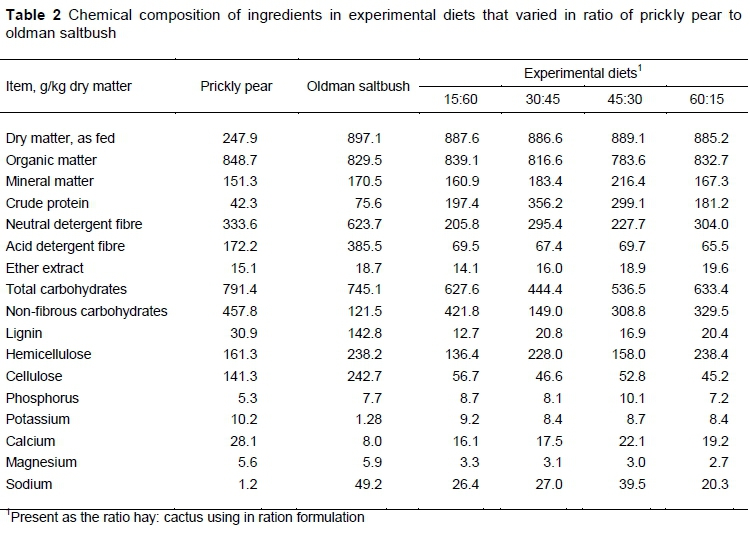
The study was divided into four 19-day periods, in which the first ten days were for animal adaptation to diets. Treatments consisted of four diets in which saltbush and prickly pear collectively made up 75% of the ration with varying percentages of each. For convenience, these diets are referred to by the percentage of hay. To make the hay, the oldman saltbush, approximately 1.5 m in height, was cut manually at 30 cm above the ground. The prickly pear was harvested at 24 months after regrowth and was offered fresh and processed daily to supply the animals. All materials were processed in a stationary forage chopper (PP-35, Pinheiro máquinas, Itapira, São Paulo, Brazil) to an average particle size of approximately 2.0 cm. The diets were provided with a total mixed ration (TMR). Nutrient composition of the oldman saltbush, prickly pear, and the concentrate mixtures used in the study are given in Table 2.
Feed was provided twice a day (at 9h00 and 14h00), and water was provided ad libitum. The leftovers were collected and weighed to determine intake and adjust the dry matter intake (DMI) to allow 10% leftovers in the trough. Samples of the food and leftovers were collected weekly for further laboratory analysis.
The DMI was obtained from the difference between the total DM of feed intake and the total DM in the leftovers. Nutrient intake was determined as the difference between the total nutrients in the feed intake and the total nutrients in the leftovers, on a total DM basis.
Dietary nutrient digestibility was estimated using an external indicator (enriched and purified lignin, LIPE®) at 0.5 g per animal. It was provided once daily in the morning, during the last four days of each experimental period. The faeces of each animal were collected twice a day (at 8h30 and 14h30) directly from a rectal funnel in labelled plastic bags, which were stored at -20 °C and processed at the end of each period. The LIPE concentration in faeces was measured by infrared spectroscopy, and faecal production was calculated by the logarithmic ratio of spectral bands (Saliba fibre, 2015).
Water intake was measured daily. It was supplied in buckets that were weighed before being supplied and 24 hours afterwards. To measure daily evaporation, three water-filled buckets were placed near the pens.
After thawing, samples of fodder, leftovers, and faeces were pre-dried in an oven at 55 °C for 72 hours and ground in Wiley mills with 1-mm sieves. Laboratory analyses were performed using the methods described by Association of Official Analytical Chemists (aoAC, 2016) for DM (method 967.03), mineral matter (M) (method 942.05), crude protein (CP) (method 981.10), and ether extract (EE) (method 920.29). Neutral detergent fibre (NDF) and acid detergent fibre (ADF) contents were determined as described by Van Soest et al. (1991). Lignin (LIG), phosphorus, potassium, calcium, magnesium, and sodium were measured by treating the ADF residue with 72% sulfuric acid (Silva & Queiroz, 2002). Total carbohydrate (TC) and cellulose (CEL) were estimated using the equations proposed by Sniffen et al. (1992):

Non-fibrous carbohydrate (NFC) content was measured as proposed by Hall (2003):

Hemicellulose (HEM) was calculated using the equation:

Nutrient apparent digestibility coefficients (ADC) were calculated using the equation:

Total digestible nutrients (TDN) were estimated using the equation developed by Sniffen et al. (1992):

On day 11 of each study period, four ruminal content samples were collected from each calf every four hours. The first sample was taken before the diet was offered in the morning (0h00), and the others 4, 8, and 12 hours after feeding.
A 50 mL aliquot of ruminal content was taken from the middle of the rumen, between the solid and liquid phases, for each animal, time (0, 4, 8, and 12 hours), and study period. These samples were measured for pH, temperature, conductivity, and salinity. Two 20 mL subsamples were then collected to determine ammoniacal nitrogen (NH3-N) according to the method of Chaney and Marbach (1962).
Nutrient degradability analysis started on day 12 of each period. Incubations were performed in the morning, before the diets were administered, and incubation periods were 0, 4, 6, 12, 24, 48, and 96 hours, whereas at time zero the samples underwent only a washing under distilled water at 39 °C. Potential degradability (PD) and effective degradability (ED) were measured using the model proposed by 0rskov and McDonald (1979).
Data were subjected to analysis of variance (ANOVA) and regression using the SAS statistical software package (SAS Institute Inc., Cary, North Carolina, USA). Effects with P < 0.05 were considered significant and differences between the associated means were evaluated using Tukey's test.
Results and Discussion
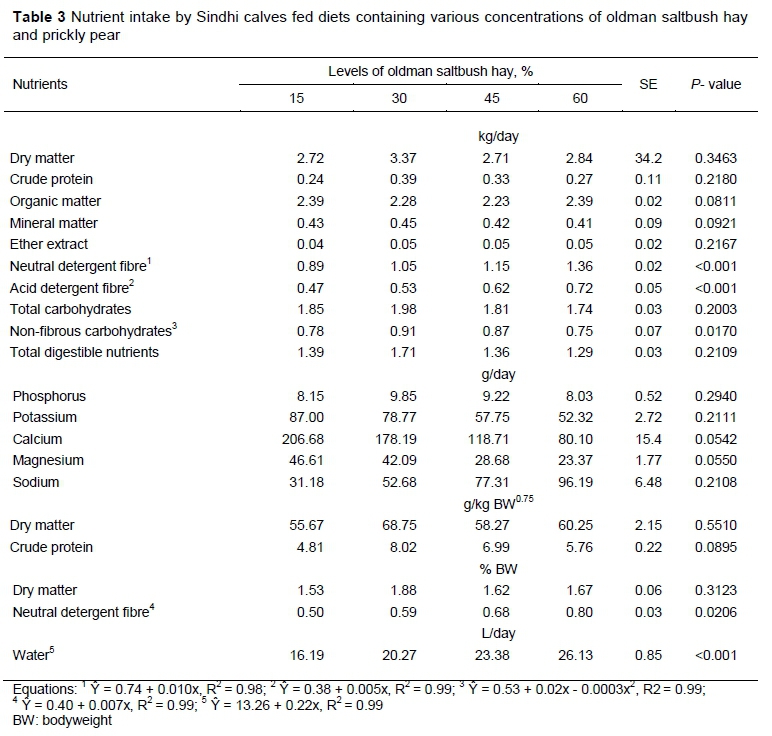
The NDF intake (in kg/day and in % LW) increased linearly (P <0.05) as levels of saltbush hay increased in the diets. A similar result was obtained for ADF intake (Table 3). NFC intake showed a quadratic response (P <0.05) and was highest using 34.83% of saltbush hay (Table 3). Water intake increased significantly (P <0.05) as the concentration of hay increased in the diets, with water intake of 219 mL for each 1% increase in hay ingestion (Table 3).
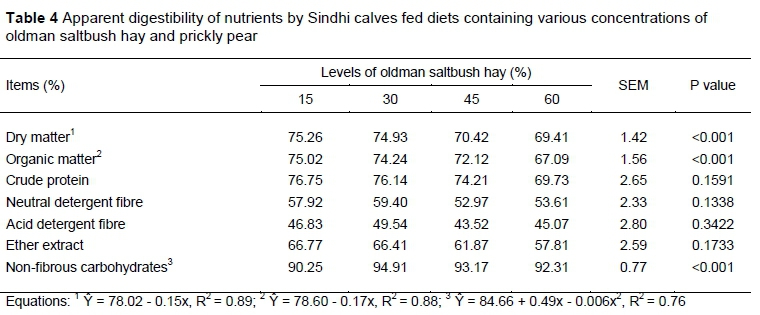
The ADC of DM and OM decreased linearly (P <0.05) as levels of saltbush hay increased in the diets, with a decrease of 0.14% and 0.17% for each 1% of ingested hay, respectively. The ADC of NFC had a quadratic response (P <0.05) and was highest in the diets containing 30% hay (Table 4).
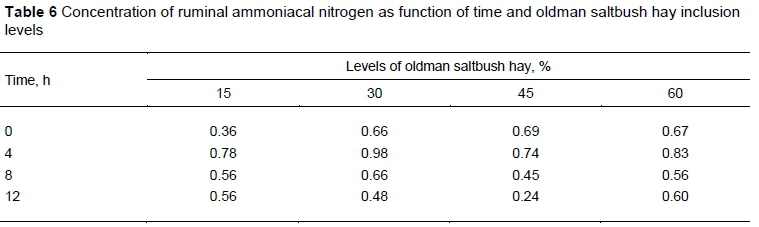
Concentrations of NH3-N were high at most time points. However, no interaction, of diet with time (Table 5). Compared with pH and temperature the level of ruminal NH3-N was highly variable (CV = 3.45 and 2.52% vs. 56.25%). For all diets the response over time was maximized at four hours of incubation (Table 6).
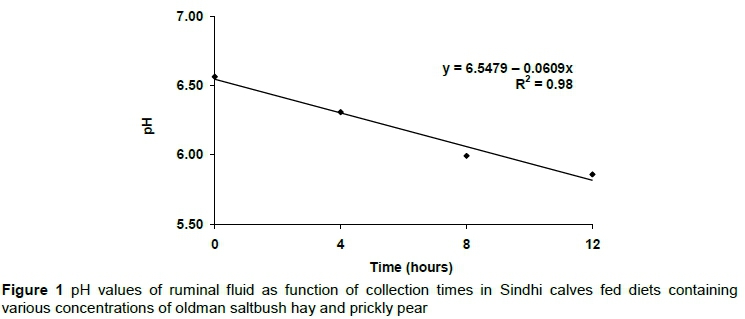
Ruminal pH showed a linearly decreasing function (P <0.05) with time, regardless of hay concentration (Figure 1). Temperature presented a quadratic function (P <0.05) at the evaluated time points (Table 5), with a maximum value at 7.66 hours, but decreased at the last time point, after 12 hours (Figure 2).
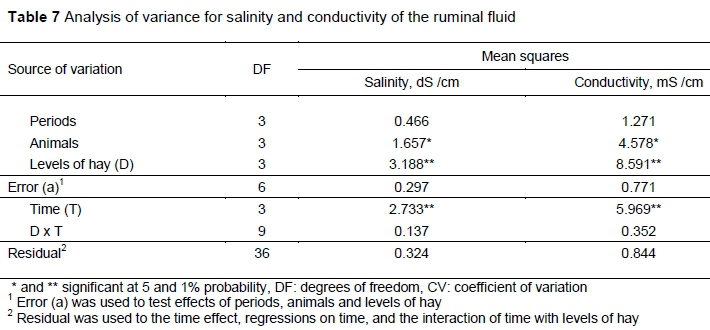
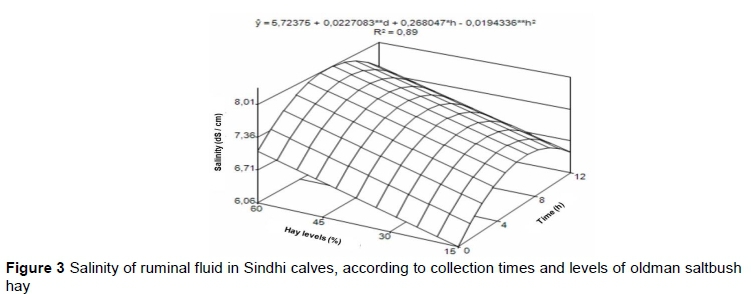
Salinity and conductivity exhibited similar relative variability with coefficients of variation of 8.02 and 7.60, respectively. The salinity and conductivity levels changed significantly (P <0.05) according to the hay levels and time points (Table 7). Treating the levels of saltbush hay as a continuous variable revealed significant linear effects for both salinity and conductivity (Figures 3 and 4) (P <0.01). No curvilinear effect or interaction of the linear effect with time was detected (P >0.05) for either of these response variables. Changes over time in salinity and conductivity both exhibited curvilinear effect (P >0.01) and also had a significant linear component (P <0.05 and P <0.01, respectively).
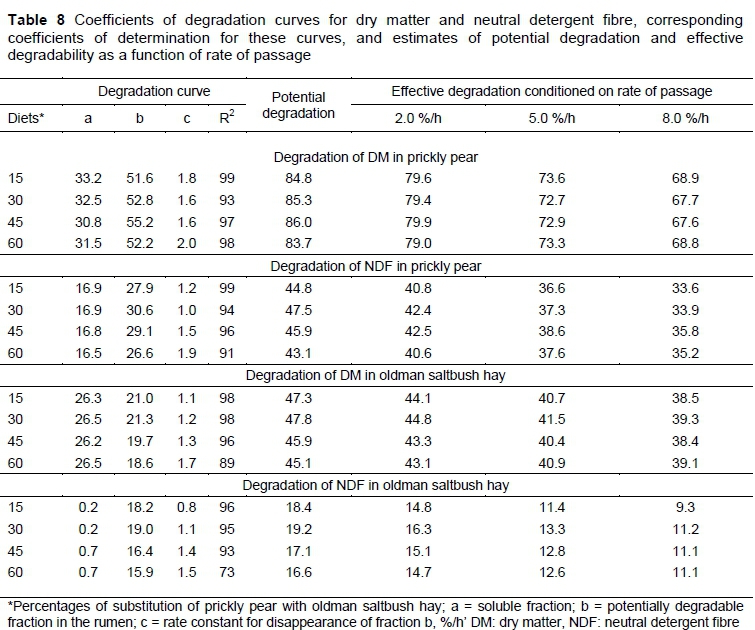
Diets that contained 15% saltbush hay had a higher degradation of fraction 'a' for prickly pear DM (33.2). The potentially degradable fraction 'b' of DM was higher (55.2) in animals fed diets containing 45% hay (Table 8). Degradation rates of fraction 'b' of DM (19.7% and 18.6%) and NDF (16.4% and 15.9%) were comparatively lower using 45% and 60% of saltbush hay in the diets (Table 8). The degradation rates of the soluble fraction of DM and NDF of both prickly pear and saltbush hay were higher in feeds containing 60% of hay (Table 8).
The PD of DM and NDF of prickly pear and saltbush hay was relatively lower in feeds containing 60% hay. However, the ED of DM in feeds containing 60% of hay was less affected by the rate of food passage and was decreased by 10.2% and 4% for prickly pear and hay, respectively, as the passage rate increased from 2.0% to 8.0%/h (Table 8).
The increase in NDF intake can be explained by its increased level in the diets (Table 1). According to the National Research Council (NRC, 2001), NDF intake should be approximately 1.2% of LW. The present results confirm this estimate. Sun et al. (2018) reported that diets with a fibre content lower than the animal requirements show a positive relationship between NDF intake and its content, within the same level of production. When NDF content is below 60%, as observed in this study, its intake is limited by energy demand rather than the effect of satiety.
The NRC (2001) indicated that ADF values lower than 30% promoted a high intake of DM. In the present study, despite the increasing ADF concentration in the diets and values being lower than 30%, DM intake was low (Table 2). This result may be related to the increased levels of lignin in the diets. That is, the differences in ADF intake may be associated with variations in the composition and nutritional value of the hay.
Van Soest (1994) showed that feeds with high NFC content are quickly fermented in the rumen, increasing the energy supply. The NFC intake did not exceed the limit of 43% in the diets, as suggested by the NRC (2001), probably because the amount of prickly pear decreased, increasing the NDF content by adding saltbush hay, hence reducing the NFC content in the diets (Table 2).
A mean TDN intake of 1.44 kg/day, as found in the present study, could meet about 52% of the daily weight gain requirements (500 g) for zebu cattle with a LW of 250 kg, which corresponds to 2.78 kg/day of TDN (Valadares Filho et al., 2009). The NRC (1996) recommends diets with a concentration of 54% of TDN (540 g/kg of DM). In this study, these levels were close to 50% (Table 2).
Given that DMI values showed no significant differences among the diets (Table 2), the concentrations of saltbush hay caused no increase in TDN intake, although the diets had NDF contents within the required limits (Table 2). However, the increasing concentrations of hay fostered the appearance of intake-limiting compounds, restricting ingestion, thus reducing nutrient availability and intake.
The increase in water intake can be explained by the high concentration of salt in the hay, increasing the need to ingest water as the hay level increased and the concentration of prickly pear decreased. Alves et al. (2017) found that the water intake of animals depended on a combination of feed type and its water content, and salty feeds increase such intake. Church (1993) indicated that the recommended intake of water for cattle weighing more than 100 kg of LW in regions with temperatures above 27 °C is 16.0 to 18.6 L/day. These values are lower than those obtained in the present study, probably because of the higher bodyweight (170 kg) of the animals.
Although it was not significant, sodium intake increased as hay intake was raised (Table 3). The sodium intake observed in diets with 30% or more hay was above the requirement of 50 g/day, as recommended by NRC (2001). However, according to Cunningham (2014), excessive intake of Na does not cause intoxication, since intakes above the recommended level are regulated through excretion via urine and faeces, and through an increase in water intake and a decrease in food intake.
The decrease in the ADC of DM and OM (Table 4) can be explained by the increase in the percentage of NDF in the diets, because NDF is negatively correlated with the ADC of DM and by the low DMI. The ADC values were 60% higher than those recommended by the Agricultural and Food Research Council (AFRC, 1993).
The high ADC of NFC presented a quadratic function and decreased with an inclusion of 30% or more hay, probably because of the decrease in the percentage of prickly pear, which has a high NFC concentration, and these carbohydrates are rapidly and fully digested by ruminants (mean of 98%). The treatment with 30% hay increased the ADC of NFC, which justifies the results obtained for DM and OM, and hence a higher TDN content.
Because bacteria preferentially hydrolyse simpler carbohydrates, the addition of easily digestible carbohydrates to feeds such as starch reduces crude fibre digestibility and, given that the energy-protein ratios of the studied diets were not synchronized, the high ADC of CP can be justified. This is because these values result from the high protein degradation in the rumen, thus resulting in low DM intake (Persichetti Júnior et al., 2014; Gomez et al., 2016).
The absence of significant results for NH3-N may be because of the effect of a high salt concentration in diets containing halophyte forage on nitrogen uptake and transport. Therefore, high salt concentrations may have inactivated enzymes and inhibited protein synthesis, because diets with increasing levels of saltbush hay presented a sodium concentration between 1.77% and 3.46% (Table 2), which is much higher than the value recommended by the NRC (2001) (0.06 - 0.08%).
The pH values recorded in this study are explained by the low levels of ingested proteins and higher fermentation after feeding because of the excess NFC in the diet (Figure 1), which decreases the ruminal pH and the efficiency of microbial protein synthesis (Costa et al., 2015). Katole et al. (2016) found that the acceptable pH range for maximum microbial growth and maximum fibre digestion was 6.2 - 6.8. Costa et al. (2015) observed that pH values lower than 6.2 inhibited the rate of digestion and increased the time for cell wall colonization.
Okoruwa et al. (2016) showed that rumen temperature is maintained relatively constant at 38 °C to 40 °C, which is higher than the values found in the present study (Figure 2). This result may be because the temperatures were not measured directly in the rumen in this study but in the collected rumen fluid.
The increasing levels of saltbush hay in diets increased salt concentrations (Figure 3). Moreno et al. (2015) observed that this forage plant is well adapted to saline soils and can extract large amounts of salt from the soil, which could be transferred to the hay. Abd El-Hack et al. (2018) reported that high salt levels in ruminant feed increased the concentration of sodium and chloride in the urine and faeces and might limit nutrient uptake, especially in arid soils and irrigated areas. The sodium concentration in the hay was 49.20 g/kg (Table 1), which is higher than the daily requirement (1.40 g/kg) for animals with a LW of 150-600 kg at an ambient temperature of approximately 30 °C.
The electrical conductivity above 10 mS/cm (Figure 4) that was found in this study is considered high and may cause health problems, especially in the kidneys, with a consequent reduction in animal productivity (German et al., 2008).
A critical characteristic of prickly pear is the rapid degradation of DM owing to the high proportion of NFC with high digestibility. Non-fibrous carbohydrates degraded rapidly in the rumen, which increases the energy supply and favours microbial growth, and hence DM degradation (Silva et al., 2018). The presence of 60% saltbush hay in the diets decreased the fraction 'b' of NDF in both saltbush hay and prickly pear. This result can be accounted for partly by the effect of the less degradable fraction of the hay on ruminal microbiota efficiency in animals fed this diet.
Despite the lower values of fraction 'b' of NDF for diets with an inclusion level of 60% saltbush hay, this treatment had the highest degradation rate of fraction 'c'. Fraction 'c' is crucial since it determines nutrient degradation as a function of retention time of food in the rumen (Belanche et al., 2014).
Conclusion
The inclusion of increasing levels of saltbush hay in diets for Sindhi calves did not induce differences in nutrient consumption. However, its inclusion in diets at levels above 30% should be done with caution owing to the possibility of decreasing the acceptability of the diet. Diets with a lower proportion of hay promoted lower water intake, owing to the higher moisture content of the prickly pear. The pH values of the rumen contents were within the acceptable range for good microbial development. However, the low temperature and observed high levels of salinity and conductivity may decrease nutrient absorption and cause health problems. These diets had an influence on the DM and NDF degradability coefficients of prickly pear and saltbush hay. The response to substitution of saltbush for prickly pear was curvilinear with an intermediate optimum. Further studies to evaluate the effects of the saltbush feeding on the internal organs and to fine-tune the level of to be fed are recommended. These studies should allow for an adequate recommendation for the dietary level at which this feedstuff can be fed without causing damage to animal health.
Authors' Contributions
JNA, GCG, and FS participated in designing the study, laboratory analysis, and manuscript writing. GGLA, LGRP, SGN, DRM, and TVV were involved in drafting and revising the manuscript for intellectual content. JNA, GCG, LCS, and FSC did the data analysis and interpretation, and were involved in the preparation and revision of the manuscript. JNA, GCG, and FSC contributed to the acquisition, analysis and interpretation of data.
Conflict of Interest Declaration
The authors declare that there are no conflicts of interests in this project.
References
Abd El-Hack, M.E., Samak, D.H., Noreldin, A.E., Arif, M., Yaqoob, H.S. & Swelum, A.A., 2018. Towards saving freshwater: Halophytes as unconventional feedstuffs in livestock feed: A review. Env. Sci. Poll. Res. 25, 1439714406. https://doi.org/10.1007/s11356-018-2052-9. [ Links ]
AFRC (Agricultural and Food Research Council), 1993. Energy and protein requirements of ruminants. CAB, Wallingford. 176 pp. [ Links ]
Alao, B.O., Falowo, A.B., Chulayo, A. & Muchenje, V., 2017. The potential of animal by-products in food systems: Production, prospects and challenges. Sust. 9, 1-18. https://doi.org/10.3390/su9071089. [ Links ]
Alves, J.N., Araújo, G.G.L., Gonzaga Neto, S., Voltolini, T.V., Santos, R.D., Rosa, P.R., Guan, L., Mcallister, T. & Neves, A.L.A., 2017. Effect of increasing concentrations of total dissolved salts in drinking water on digestion, performance and water balance in heifers. J. Agric. Sci. 155, 847-856. https://doi.org/10.1017/S0021859617000120 [ Links ]
Association of Official Analytical Chemists (AOAC), 2016. Official methods of analysis. 20th ed. Washington DC. [ Links ]
Belanche, A., Weisbjerg, M.R., Allison, G.G., Newbold, C.J. & Moorby, J.M., 2014. Measurement of rumen dry matter and neutral detergent fibre degradability of feeds by Fourier-transform infrared spectroscopy. J. Dairy Sci. 97, 2361-2375. https://doi.org/10.3168/jds.2013-7491 [ Links ]
Celi, P., Cowieson, A.J., Fru-Nji, F., Steinert, R.E., Kluenter, A.M. & Verlhac, V., 2017. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Tech. 234, 88100. https://doi.org/10.1016/j.anifeedsci.2017.09.012 [ Links ]
Chaney, A.L., Marbach, E.P., 1962. Modified reagents for determination of urea and ammonia. Clin. Chem. 8, 130-146. https://www.ncbi.nlm.nih.gov/pubmed/13878063. [ Links ]
Church, D.C., 1993. The ruminant animal: Digestive physiology and nutrition. 3rd edition. Acribia, Zaragoza. 640 p. [ Links ]
Council for the Control of Animal Experimentation (CONCEA), 2008. Procedures for the Scientific Use of Animals. Based on Clause VII of the 1st Paragraph in Article 225 of the Brazilian Federal Constitution. Brasília, DF, Brazilian Government through the National Council for the Control of Animal Experimentation (CONCEA) and Institutional Animal Care and Use Committees (CEUA), Brazil. [ Links ]
Costa, N.L., Monteiro, A.L.G. & Silva, A.L.P., 2015. Considerations on the fibre degradation of tropical forage associated with energy or nitrogenous supplements. Arch. Zootec. 64, 31-41. https://doi.org/10.21071/az.v64i247.504 [ Links ]
Cunningham, J.G., 2014. Tratado de fisiologia veterinária. 5th edition. Guanabara Koogan, Rio de Janeiro 624 p. [ Links ]
EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária), 2014. Dados meteorológicos. http://www.cpatsa.embrapa.br:8080/servicos/dadosmet/ceb-dia.html [ Links ]
German, D., Thiex, N. & Wright, C., 2008. Interpretation of water analysis for livestock suitability. C274. South Dakota State Univ., Brookings, South Dakota, USA. [ Links ]
Gómez, L.M., Posada, S.L. & Olivera, M., 2016. Starch in ruminant diets: A review. Rev. Colomb. Ci. Pec. 29, 77-90. https://doi.org/10.17533/udea.rccp.v29n2a01 [ Links ]
Hall, M.B., 2003. Challenges with nonfibre carbohydrate methods. J. Anim. Sci. 81, 3226-3232. https://doi.org/10.2527/2003.81123226x. [ Links ]
Helal, H.G., Eid, E.Y., Nassar, M.S., Badawy, H.S. & El Shaer, H.M., 2018. Comparative nutritional studies of ewes and does fed salt tolerant plants under desert condition. Nat. Sci. 16, 62- 72. https://doi.org/10.7537/marsnsj160618.10. [ Links ]
Katole, S., Lende, S. & Borah, L., 2016. Nutrition and acidosis in ruminants. Int. J. Anim. Vet. Sci. 3, 35-39. http://jakraya.com/journal/pdf/3-ijavsArticle_10.pdf. [ Links ]
Moreno, G.M.B., Borba, H., Araújo, G.G.L., Sanudo, C., Silva Sobrinho, A.G., Buzanskas, M.E., Lima Júnior, D.M., Almeida, V.V.S. & Boaventura Neto, O., 2015. Meat quality of lambs fed different saltbush hay (Atriplex Nummularia) levels. Italian J. Anim. Sci. 14, 251-259 3302. https://doi.org/10.4081/ijas.2015.3302 [ Links ]
Norman, H.C., Masters, D.G. & Barrett-Lennard, E.G., 2012. Halophytes as forages in saline landscapes: Interactions between plant genotype and environment change their feeding value to ruminants. Env. Exp. Bot. 92, 96-109. https://doi.org/10.1016/j.envexpbot.2012.07.003. [ Links ]
NRC (National Research Council), 2001. Nutrient requirements of dairy cattle. 6th rev. ed. The National Academy Press, Washington DC. 381 pp. [ Links ]
NRC ((National Research Council), 1996. Nutrient requirements of beef cattle. The National Academy Press: Washington DC. 242 pp. [ Links ]
Nunes, A.T., Lucena, R.F.P., Santos, M.V.F. & Albuquerque, U.P., 2015. Local knowledge about fodder plants in the semi-arid region of Northeastern Brazil. J. Ethnobiol. Ethnomed. 11, 1-12. https://doi.org/10.1186/1746-4269-11-12. [ Links ]
Okoruwa, M.I., Bamigboye, F.O. & Agbadu, A., 2016. Rumen metabolites and thermo-physiological response of West African dwarf sheep as influenced by ficus foliage with differently processed breadfruit meals. Global J. Agric. Res. 4, 28-38. https://www.eajournals.org/journals/global-journal-of-agricultural-research-gjar/vol-4-issue-5-november-2016/rumen-metabolites-thermo-physiological-response-west-african-dwarf-sheep-influenced-ficus-foliage-differently-processed-breadfruit-meals/ [ Links ]
0rskov, E.R. & McDonald, I., 1979. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agr. Sci. 92, 449-453. https://doi.org/10.1017/S0021859600063048 [ Links ]
Persichetti Júnior, P., Almeida Júnior, G.A., Costa, C., Meirelles, P.R.L., Silveira, J.P.F., Panichi, A., Silva, M.G.B., Factori, M.A., Cavasano, F.A. & Mendonça, A.S., 2014. Nutritional value of high moisture corn silage in the diet of Holstein cows. Arq. Bras. Med. Vet. Zootec. 66, 1495-1503. http://dx.doi.org/10.1590/1678-5999. [ Links ]
Saliba, E.O.S., Faria, E.P., Rodriguez, N.M., Moreira, G.R., Sampaio, I.B.M., Saliba, J.S., Gonçalves, L.C., Borges, I. & Borges, A.L.C.C., 2015. Use of infrared spectroscopy to estimate fecal output with marker Lipe®. Int. J. Food Sci. Nutr. Diet. 4, 1-10. http://dx.doi.org/10.19070/2326-3350-SI04001 [ Links ]
SAS, 2015. SAS/Stat university user guide. Cary, NC. 8621 pp. [ Links ]
Silva, D.J. & Queiroz, A.C., 2002. Análise de alimentos: Métodos químicos e biológicos. UFV Viçosa. 235 pp. [ Links ]
Silva, R.C., Ferreira, M.A., Oliveira, J.C.V., Santos, D.C., Gama, M.A.S., Chagas, J.C.C., Inácio, J.G., Silva, E.T.S. & Pereira, L.G.R., 2018. Orelha de Elefante Mexicans (Opuntia stricta [Haw.] Haw.) spineless cactus as an option in crossbred dairy cattle diet. South Afr. J. Anim. Sci.48, 516-525. http://dx.doi.org/10.4314/sajas.v48i3.12 [ Links ]
Silva, T.G.F., Araújo, G.G.L., Moura, M.S.B. & Souza, L.S.B., 2017. Agrometeorological research on forage cactus and its advances in Brazil. Am. J. Plant Res. 2, 45-68. https://ajpr.online/wp-content/uploads/2017/11/Agrometereology-Silva-et-al.pdf [ Links ]
Sniffen, C.J., O'Connor, J.D. & Van Soest, P.J., 1992. A net carboydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci. 70, 3562-3577. https://www.ncbi.nlm.nih.gov/pubmed/1459919 [ Links ]
Sun, J., Xu, J., Shen, Y., Wang, M., Yu, L., Wang, H., 2018. Effects of different dietary ratio of physically effective neutral detergent fibre and metabolizable glucose on rumen fermentation, blood metabolites and growth performance of 8- to 10-month-old heifers. Asian-Austral. J. Anim. Sci. 31, 1230-1237. http://dx.doi.org/10.5713/ajas.17.0885 [ Links ]
Valadares Filho, S.C., Chizzotti, M.L. & Paulino, P.V.R., 2009. Nutrient requirements of beef cattle in Brazil: Challenges. Rev. Ceres 56, 488-495. http://www.ceres.ufv.br/ojs/index.php/ceres/article/view/3457/1355 [ Links ]
Van Soest, P.J., 1994. Nutritional ecology of the ruminant. 2nd edition. Cornell University Press, Ithaca. 476 pp. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fibre, neutral detergent fibre, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. https://doi.org/10.3168/jds.S0022-0302(91)78551-2 [ Links ]
Submitted 19 October 2019
Accepted 6 May 2020
First published online 21 June 2020
# Corresponding author: glayciane_gois@yahoo.com.br


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}