










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.50 no.1 Pretoria 2020
http://dx.doi.org/10.4314/sajas.v50i1.6
ARTICLES
Effects of polyunsaturated fatty acids in diets fed to sows on fatty acids in brain, muscle and skin of their piglets
I. De GasperínI; J.G. VicenteI, #; J.M. Pinos-RodríguezI; F. MontielI; R. LoezaI; J.M. Martínez-HernándezI; B. IsabelII
IFacultad de Medicina Veterinária y Zootecnia, Universidad Veracruzana. 91710. Veracruz, México
IIFacultad de Veterinaria, Universidad Complutense de Madrid, 28040, Madrid, Espana
ABSTRACT
The aim of this research was to determine fatty acid profiles in piglet brain, skin, and muscle, and in the milk of sows fed fat with different saturation grades during gestation and lactation. At 42 days of gestation, 50 multiparous sows were randomly allocated to one of two treatments, namely a diet containing pork lard (n = 25) and a diet containing soybean oil (n = 25). The fats were provided at 3.6% during gestation and at 4% during lactation. The experimental diets were offered through the weaning of the piglets. The fatty acid profile of the milk was determined fourteen days after parturition. At weaning (21 days postpartum) and seven days later, one of the piglets (n = 64) from 16 sows allocated to each treatment was selected at random to determine fatty acid profiles in brain, skin and muscle. Saturated and monounsaturated fatty acids were higher in the diet with pork lard than in that with soybean oil, in which the polyunsaturated fat content was higher. A higher saturation of fatty acids was found in milk from the sows that consumed pork lard, which contained more saturated fatty acids than the milk from sows that consumed soybean oil. The fatty acid profiles in muscle and skin of the piglets were affected by the diet of the sows. However, the fatty acid profile of the piglets' brains was not affected by the diet of their mothers.
Keywords: fat saturation, lard, piglet survival, sow feeding, soybean oil
Introduction
Fatty acids (FA) of the n-3 series are a structural component of the phospholipid membrane of neurons and are found in high concentrations in the brain and retina. Their use in diets for sows has been shown to improve foetal development and neonatal vigour (Tanghe et al., 2014; Francine et al., 2010). The synaptic membranes of the brain contain high amounts of docosahexaenoic acid (DHA) and arachidonic acid (ARA), which have been shown to intervene in the learning process, exploration and survival of piglets (Farmer et al., 2010). Docosahexaenoic acid is essential and is produced naturally. However, it can be synthesized using a-linolenic acid as a precursor, but this process requires adequate and necessary ratios of omega 6 to omega 3 (n6/n3) owing to high competition for elongase and desaturase enzymes (Brenna et al., 2009). The n-3 fatty acids are of great importance in the diet of sows during pregnancy, since the foetus depends on their transfer through the placenta and during lactation for piglet development (Chung et al., 2008). On the other hand, growing concern about the relationship between human diet and health has increased interest in modifying the FA profile of products derived from pigs (Tous et al., 2012). Several studies (Cordero et al., 2011; Laws et al., 2012; Bai et al., 2017) have reported that it is possible to modify the FA profile of colostrum and milk from sows, depending on the type of fat that is added to their diets. These fats pass directly to the milk and colostrum with minimal or with no modification and are deposited in the tissues of the piglets to be mobilized later during the weaning phase. The objective of the present study was to determine the effect of soybean oil as a source of highly polyunsaturated FA in sow diets on the FA profile of milk and on the lipid composition of piglet nerve tissue (encephalon) and on that of tissues of commercial value (muscle and skin) to determine the possibility of modifying the FA profile of the piglet carcass.
Materials and methods
Animal management procedures were conducted in compliance with the Mexican official norm (NOM-033-ZOO-1995: Humanitarian slaughter of domestic and wild animals) and the Bioethics and Animal Welfare Committee of the Facultad de Medicina Veterinaria y Zootécnia of the Universidad Veracruzana (2018-014). The study was conducted on a commercial farm in Veracruz, Mexico (19 ° 27'28 "N; 96 ° 45'57" W), which has an average annual temperature of 26 °C and 43% relative humidity. Fifty Landrace x Large White multiparous sows that were 23 to 38 months old and 42 days into pregnancy were randomly allocated to one of two groups: i) (n = 25), a diet containing 3.6% pork lard, and ii) (n = 25), a diet containing 3.6% soybean oil, for the entire gestation period. Those fat sources were 4% during the lactation phase (21 days). The diets were prepared weekly at the farm feed factory with an automatic feed mixer machine and stored in a silo. All sows were housed in individual gestation cages (0.65 m wide x 2.20 m long) with concrete floor, nipple water dispenser and individual feeders. Seven days before the date of birth, the sows were housed in maternity cages (2.2 m x 2.4 m) with a plastic grid floor and a piglet area (1.0 m x 1.2 m).
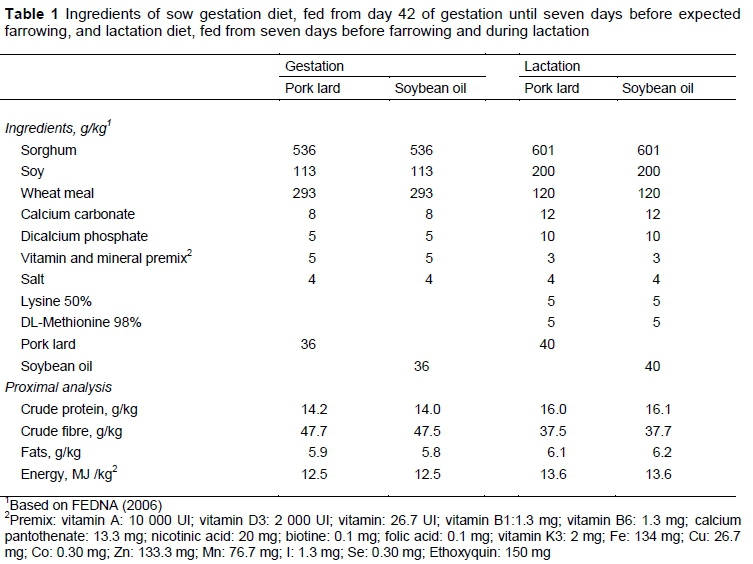
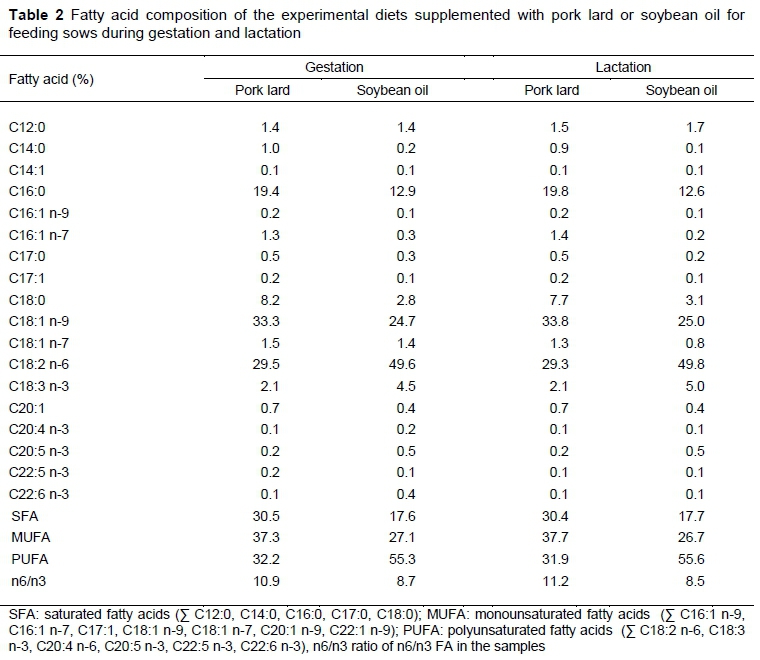
Experimental diets had similar levels of energy and protein, and were formulated for pregnant sows (from 42 days of pregnancy to one week before parturition) and lactating sows (from delivery to day 1 before weaning at 21 days), according to the FEDNA tables (2006) for swine (Table 1). Pregnant and lactating sows were fed diets with pork lard or soybean oil in a n6/n3 ratio of 11:1 and 8.5:1, respectively (Table 2). The two experimental diets were randomly assigned to 50 sows. Crude protein, crude fibre, ether extract and metabolizable energy of the experimental diets were quantified according to AOAC methodology (2005).
Fourteen days after birth, 20 mL milk from each sow was sampled by manual milking and conserved to determine the FA profile. One day before weaning (20 days after birth), one piglet from each litter per treatment was taken randomly and slaughtered according to the Mexican official norm NOM ZAG-ZOO-033. In the same way, one piglet from each treatment was slaughtered one week after weaning (day 29 after delivery). Samples of the long dorsal muscle, encephalon, and skin were taken from each piglet. All tissues were lyophilized for further analysis. To determine the FA profile of experimental diets, weekly samples of each diet were taken during the entire study and kept frozen at -20°C until further analysis. The samples of food, milk, and tissues were analysed by the technique of Ruíz et al. (2004) and a modification of the technique of Berdeaux et al. (1999). To determine FA profile, the authors used a gas Chromatograph (HP™ model 6890 GC, California, USA) equipped with flame ionization detector and an HP-Innowax polyethylene glycol column (30 m x 0.316 mm x 0. 25 urn), (Hewlett-Packard™, California, USA) (injection at 170 °C and then increased 3.5 °C/min to 210 °C). The chromatograms were integrated by the pattern of retention times of polyunsaturated fatty acids (PUFA) of marine origin (PUFA Sigma-Aldrich, Darmstadt, Germany).
The experimental design consisted of random assignment. The UNIVARIATE procedure was used to test the normality of data distribution. Data were analysed with a MIXED procedure using SAS (2008), where the treatments were considered fixed components and sow and piglet random components. Also, a Student test was adopted to detect differences between means. A probability of less than 0.05 was considered statistically significant.
Results and discussion
The FA profile of the diets was proportional to the FA in the fat sources (Table 2), similar to that reported in studies (Rooke et al., 2001; Lauridsen & Danielsen, 2004; Bai et al., 2017).
The degree of unsaturation of the diets was increased with the addition of soybean oil, which contains 84% unsaturated fatty acids. In contrast, the diet with lard, which contains only 32% polyunsaturated fatty acids had a higher level of saturation. The n6/n3 ratio was lower in the diet with soybean oil due to its high content of alpha linolenic acid (C18:3 n-3).
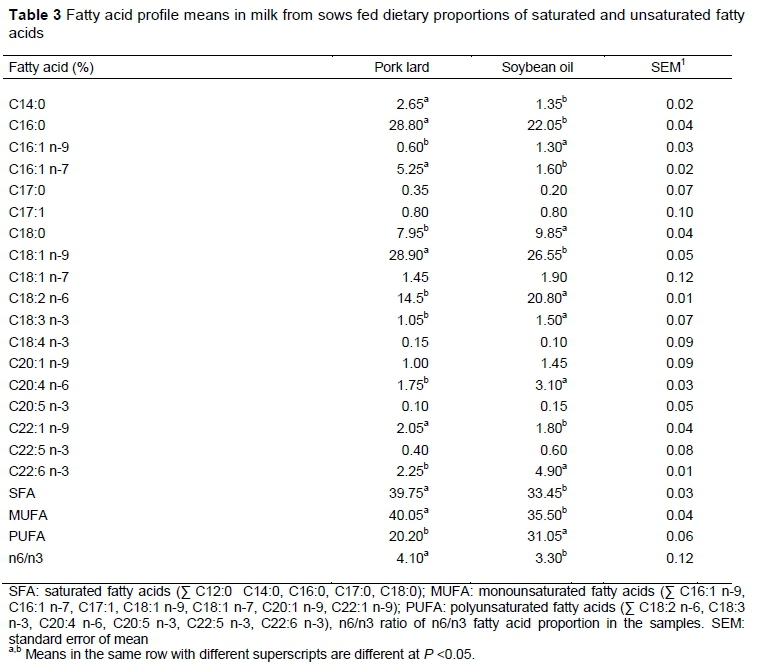
The FA profile of the milk on days 1 and 14 was proportional to that found in experimental diets. Greater polyunsaturation was observed in the milk of sows fed diets that included soybean oil (Table 3), corroborating the findings of several authors (Lauridsen & Danielsen, 2004; Leonard et al., 2010; Cordero et al., 2011), who indicated that fat is the only component of milk that can be modified by diet. The addition of polyunsaturated fats to the sow diet increases the polyunsaturation of milk and colostrum. The negative energy balance in the early postpartum period of the sow causes slow mobilization of body reserves and the use of dietary fat, rather than subcutaneous fat, for milk production is prioritized (Vicente et al., 2013).
At the beginning of lactation, sows initially use their body reserves for the production of milk. However, around the ninth day of lactation the milk produced begins to limit piglet development. Being larger animals, they have greater ability to breastfeed, which intensifies milk production, and dietary fats for this purpose are used quickly (Morillo et al., 2013).
There are reports in the literature that assessed sow diets supplemented with omega 3 (Rooke et al., 2001; Missotten et al., 2009; Farmer et al., 2010), especially DHA. However, the benefits of adding this fatty acid can be nullified by the high content of omega 6 because of the competition between the 18-carbon n-3 and n-6 FA for the enzymes necessary for elongation (A5 and A6 desaturase). These enzymes have a greater affinity with n-3 FA than with n-6. In sows, the expression of these enzymes can be increased at muscular level by supplementing with fish or flax oil (Missotten, et al., 2009). Eicosatetraenoic acid (C20:4 n-3), from a-linoleic acid, being an n-3 FA has a high affinity with the enzyme A-5 desaturase, so that when metabolized, it is converted into eicosapentaenoic acid (C20:5, n-3). For this reason, it is detected in the diet, but not in the milk. On the other hand, arachidonic acid C20:4 n-6 can be mobilized from the carcass reserves of the sow or elongated and desaturated from linoleic acid (Laws et al., 2012; Bai et al., 2017).
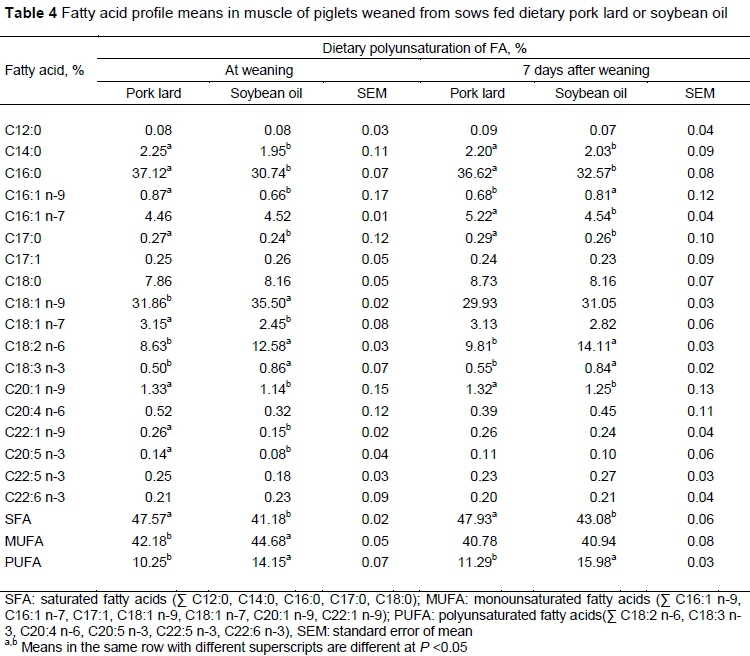
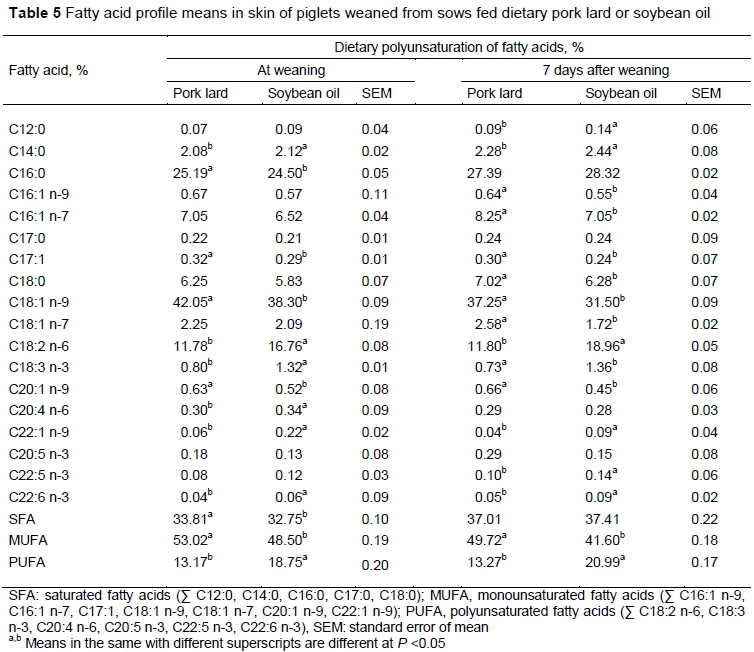
The FA profile and the level of saturation of milk were similar to those of the diets because the sows prioritize the use of ingested fat for milk production and reduce its accumulation in adipose tissue, so that FAs pass to the milk without major modifications (Lauridsen & Danielsen, 2004; Bai et al., 2017). The dietary FA profile also modified the FA profile of muscle (Table 4) and skin (Table 5) of the piglets.
In muscle tissue, the diet with soybean oil increased the percentages of oleic (C18:1 n-9) linoleic, and alpha linoleic acid and the general proportion of monounsaturated and polyunsaturated AG. Similar results were reported by Vicente et al. (2013), who found that the inclusion of lard in the diet of lactating or prepartum sows increased the concentration of MUFAs and reduced the percentage of PUFAs in the muscle of the piglets as a source of fat with low polyunsaturation.
The FA profile in the brains of piglets was not modified by treatments (Table 6). The highest levels of FA found in this tissue were palmitic (C16:0), stearic (C18:0), oleic (C18:1 n-9), arachidonic (C20:4 n-6) and docosahexaenoic (C22:6 n-3). Authors (Palmquist et al., 2009; Francine et al., 2010; Tanghe et al., 2014) mention that nerve tissue requires high amounts of DHA for neuronal function and mammals are able to synthesize DHA from its precursor Alpha-linoleic acid by a series of enzymatic reactions that enlarge it to C24:6 n-3, followed by beta-oxidation to convert it to DHA, which can be metabolically utilized. In the brain it is used mainly for phospholipid synthesis as a part of neuron membranes.
Childs et al. (2008) reported that large amounts of c22:5 n-3 were found in the brains of piglets born to sows whose diets were DHA restricted. They concluded that DHA can be replaced by structural phospholipids. However, because of its function in neurons, it cannot be replaced in its entirety. In the current study, the authors found high levels of DHA in the brain during weaning and a week after weaning, indicating that the presence of DHA, and its precursors (C18:3 n3, C20:5 n-3 and C22 5 n-3), in the diet of the sow and in milk was adequate. In that study, transfer of n-3 PUFA FA from the sow's diet to piglet tissues depended on the source of fat and the tissue in which they were deposited. Higher MUFA concentrations were observed with high saturated fat diets and PUFA in polyunsaturated diets in tissues except for liver and brain.
There is no information in the literature about the maximal n6/n3 FA ratio that limits endogenous brain synthesis of DHA and ARA in pigs. In the current study, however, the proportions of these FA in sow diets or in milk were the same in the brain.
Conclusions
The current results indicate that it is possible to increase the degree of unsaturation of fat in sow milk and in the muscle and skin of piglets by increasing unsaturation of the dietary fat. Under the experimental conditions of the current study, there was no effect of PUFA levels in the diet on the fatty acid profile of piglet brains. For future studies, it is recommended that more FA sources should be tested and that, additionally, the FA profile in tissues from finished fattening swine's be determined.
Acknowledgments
The authors thank the National Council of Science and Technology (México) for the grant awarded number 443177.
Authors' Contributions
All authors contributed equally.
Conflict of Interest Declaration
None.
References
AOAC, 2005 Official methods of analysis, 18th ed. Association of Official Analytical Chemists, Arlington, VA, USA. [ Links ]
Bai, Y.S., Wang, C Q., Zhao, X., Shi, B. & Shan, A.S., 2017. Effects of fat sources in sow on the fatty acid profiles and fat globule size of milk and immunoglobulins of sows and piglets. Anim. Feed Sci. Tech. 234, 217-227. http://doi.org/10.1016/j.anifeedsci.2017.10.006 [ Links ]
Berdeaux, O., Márquez, G.R. & Dobarganes, C., 1999. Selection of methylation procedures for quantitation of short-chain glycerol-bound compounds formed during thermoxidation. J. Chromatogr. 2, 171-181. http://doi.org/.org/10.1016/S0021-9673(99)00974-7 [ Links ]
Brenna, J.T., Salem, N., Sinclair, A.J. & Cunnane, S.C. 2009. a-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fatty Acids 80, 85-91. http://doi.org/10.1016/j.plefa.2009.01.004 [ Links ]
Childs, S., Hennessy, A.A., Sreenan, J.M., Wathes, D.C., Cheng, Z. & Stanton, C., 2008. Effect of level of dietary n-3 polyunsaturated fatty acid supplementation on systemic and tissue fatty acid concentrations and on selected reproductive variables in cattle. Theriogenology 70, 595-611. http://doi.org/10.1016/j.theriogenology.2008.04.002 [ Links ]
Chung, W.L, Chen, J.J. & Su, H.M., 2008. Fish oil supplementation of control and (n-3) fatty acid-deficient male rats enhances reference and working memory performance and increases brain regional docosahexaenoic acid levels. J. Nutr. 138, 1165-1171. http://doi.org/10.1093/jn/138.6.1165 [ Links ]
Cordero, G., Isabel, B., Morales, J., Menoyo, D., Dazac, C. & López-Bote, C.J., 2011. Conjugated linoleic acid (CLA) during last week of gestation and lactation alters colostrum and milk fat composition and performance of reproductive sows, Anim. Feed Sci. Tech, 168: 232-240. http://doi.org/10.1016/j.anifeedsci.2011.04.085 [ Links ]
Farmer, C., Giguere, A. & Lessard, M., 2010. Dietary supplementation with different forms of flax in late gestation and lactation: Effects on sow and litter performances, endocrinology, and immune response. J. Anim. Sci. 88, 225-237. http://doi.org/10.2527/jas.2009-2023 [ Links ]
Federación Espanola para el Desarrollo de la Nutrición Animal (FEDNA). 2006. Necesidades nutricionales para el ganado porcino. In: De Blas, C. y Mateos, G. Normas FEDNA. Escuela Superior de Ingenieros Agrónomos. Madrid, Espana. pp 5-24. (in Spanish) [ Links ]
Francine, Q., Gaelle, B. & Jacques, M., 2010: Linseed oil in the maternal diet increases long chain-PUFA status of the foetus and the newborn during the suckling period in pigs. Brit. J. Nutr. 104, 533-543. http://doi.org/10.1017/S0007114510000772. [ Links ]
Lauridsen, C. & Danielsen, V., 2004. Lactational dietary fat levels and sources influence milk composition and performance of sows and their progeny. Livest. Prod. Sci. 91, 95-105. http://doi.org/10.1016/j.livprodsci.2004.07.014 [ Links ]
Laws, J., Amusquivar, E., Law, A., Herrera, E., Lean, J.I., Dodds, P.F. & Clarke, L., 2012. Supplementation of sow diets with oil during gestation: Sow body condition, milk yield and milk composition. Liv. Prod. Sci. 123: 88-86. http://doi.org/.org/10.1016/j.livsci.2008.10.012 [ Links ]
Leonard, S.G., Sweeney, T., Bahar, B., Lynch, B.P. & O'Doherty, J.V., 2010. Effect of maternal fish oil and seaweed extract supplementation on colostrum and milk composition, humoral immune response, and performance of suckled piglets. J. Anim. Sci. 88, 2988-2997. http://doi.org/10.2527/jas.2009-2764 [ Links ]
Missotten, J., De Smet, S. Raes, K. & Doran, O., 2009. Effect of supplementation of the maternal diet with fish oil or linseed oil on fatty-acid composition and expression of A5- and A6-desaturase in tissues of female piglets. Animal 3, 1196-1204. http://doi.org/10.1017/S1751731109004455 [ Links ]
Morillo, A.A., Álvarez, R.J., Villalba, M.G. & Cano, L.G., 2013. La composición de la dieta de las cerdas lactantes y la producción láctea FEDNA, Madrid, Espana. 40. pp. [ Links ]
Palmquist, D.L., 2009. Omega-3 fatty acids in metabolism, health and nutrition and for modified animal products foods. Prof. Anim. Sci. 25, 207-249. http://doi.org/10.15232/S1080-7446(15)30713-0 [ Links ]
Rooke, J.A., Sinclair, A.G., Edwards, S.A., Cordoba, R., Pkiyach, S., Penny, P.C., Penny, P., Finch, A.M. & Horgan, G.W., 2001. The effect of feeding salmon oil to sows throughout pregnancy on pre-weaning mortality of piglets. J. Anim. Sci. 73: 489-500. http://doi.org/10.1017/s135772980005846x [ Links ]
Ruiz, J., Antequera T., Andres, A.I., Petron, M.J. & Muriel, E., 2004. Improvement of a solid phase extraction method for analysis of lipid fractions in muscle foods. Anal. Chim. Acta. 520, 201-205. http://doi.org/10.1016/j.aca.2004.04.059 [ Links ]
SAS, 2008. Statistical Analysis System user's guide (release 9.1.3). SAS Institute, Inc., Cary, North Carolina, USA. [ Links ]
Tanghe, S., Millet, S., Missotten, J., Vlaeminck, B. & De Smet, S., 2014. Effects of birth weight and maternal dietary fat source on the fatty acid profile in piglet tissue. Animal 8, 1857-1866. http://doi.org/10.1017/s1751731114001724 [ Links ]
Tous, N., Theil, P.K., Lauridsen, C., Lizardo, R., Vilà, B. & Esteve-Garcia, E., 2012. Dietary conjugated linoleic acid modifies gene expression in liver, muscles, and fat tissues of finishing pigs. J. Anim. Sci. 90, 340-342. http://doi.org/10.2527/jas.53768 [ Links ]
Vicente, J.G., Isabel, B., Cordero, G. & Lopez-Bote, C.J., 2013. Fatty acids profile of the sow diet alters fat metabolism and fatty acid composition in weanling pigs. Anim. Feed Sci. Tech. 181, 45-53. http://doi.org/10.1016/j.anifeedsci.2013.02.002 [ Links ]
Received 11 October 2019
Accepted 4 January 2020
First published online 16 February 2020
# Corresponding author: jvicente@uv.mx


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}