










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 n.4 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i4.19
ARTICLES
Effects of selenium and flaxseed on selenium content and antioxidant properties of eggs and immune response in hens
H. MoslehiI; B. NavidshadI, #; S.D. SharifiII; F.M. AghjegheshlaghI
IDepartment of Animal Science, University of Mohaghegh Ardabili, Ardabil, Iran
IIDepartment of Animal Science, Abureyhan Campus, University of Tehran, Iran
ABSTRACT
This study aimed to evaluate effects of supplementing the diets of laying hens with different sources of Se and different levels of flaxseed on egg Se concentration, yolk antioxidant properties and immune response. In a completely randomized design, 384 Hy-Line W-36 hens (aged 50 weeks) were assigned to a 3 ÷ 4 factorial arrangement, comprising four replicates of eight hens. For 10 weeks, the birds were fed one of three basal diets containing 0, 50, and 100 g/kg of flaxseed and regular Se content from mineral premix, supplemented with no additional Se (SN) and 1 mg/kg of Se from sodium selenite (SS), Se-enriched yeast (SY), and hydroxy selenomethionine (soH). The greatest Se concentration in the yolk was observed in diets containing organic Se sources. The SOH diet produced the greatest albumen Se concentration, followed by SY, and then SS. Using any Se supplementation improved glutathione peroxidase activity and total antioxidant capacity, compared with SN. However, superoxide dismutase activity and malondialdehyde were not affected by Se supplementation. Cell-mediated immune response was improved by Se supplementation, especially SS. Antibody response against sheep red blood cell (SRBC) was not affected by Se source, except that IgM was greater in the hens that received SS and SY. Neither flaxseed supplementation, nor its interaction with Se source was significant for any trait. Thus, supplementation of diets with 1 mg/kg Se, especially from organic sources, enriched the egg and improved some yolk antioxidant properties and immune system function. Supplemental flaxseed did not influence any measured traits.
Keywords: egg albumin, egg yolk, Immune function, inorganic selenium, organic selenium
Introduction
Eggs are an important foodstuff in basic nutrition, serving as a source of protein, fat, and micronutrients. Manipulation of egg yolk composition through improving its polyunsaturated fatty acid (PUFA) content, particularly omega-3 fatty acids, is considered a common goal in developing eggs as a functional food (Miranda et al., 2015). Flaxseed oil, which contains about 70% PUFA, mostly alpha-linolenic acid (ALA), and has the highest PUFA/SFA ratio among oilseeds, is known to be an appropriate source to increase omega-3 in diets (Zambiazi et al., 2007).
Enrichment of egg yolk with omega-3 PUFA is often associated with increased susceptibility to lipid peroxidation, owing to increased free radical production, especially in high stress conditions. To protect the yolk, tissues and cells from oxidative damage, dietary supplementation can be implemented, with suitable doses of natural antioxidants, including vitamin E, carotenoids, and Se. Antioxidants may be transferred from the feed into the egg yolk, thus producing antioxidant-enriched eggs (Surai & Fisinin, 2012).
Supplementing animal feeds with Se can provide a means to increase human Se intake. As an antioxidant, Se is likely to contribute to better shelf-life in animal products. The most important contributor to its protective antioxidant function is glutathione peroxidase (GSH-Px), which acts against harmful reactive oxygen species at various sites in the body, including the gastrointestinal tract, plasma, and in biological membranes (Fasiangova & Borilova, 2017).
Sodium selenite (SS) is the most common inorganic form of Se supplementation because of its low cost, but it has limited biological utilization. Organic Se compounds such as selenomethionine (Se-Met) and selenocysteine (Se-Cys) are part of proteins, and have higher bioavailability and lower toxicity (Surai & Fisinin, 2014; Jing et al., 2015). Some new sources of organic selenium supplements are currently being used in poultry nutrition (Suchy et al., 2014), such as Se-enriched yeast (SY), Se-enriched unicellular alga chlorella, and Se chelates such as Zn-L-Se-Met (zinc selenomethionine), with Se-Met as the major component. Recently, a new organic Se source, known as 2-hydroxy-4 methylselenobutanoic acid (HMSeBA or hydroxyl Se-Met or Selisseo) (SOH), has been developed and is reported to have increased bioavailability compared with SY and SS (Jlali et al., 2014).
Numerous studies have found that the level and form of Se can variously affect the egg Se reserves, antioxidant stability, and the immunity response of birds. According to European regulations for feed additives, the suitable amount of Se supplementation is 0.3 mg/kg of complete feed. However, other studies have found that a higher level of this element can be used in diets for poultry, resulting in better antioxidant properties and higher Se content in eggs (Suchy et al., 2014; Fasiangova & Borilova, 2017; Fasiangova et al., 2017). Supplementation with SY increased the Se content in eggs, especially in albumen, by up to twice the level that was achieved using SS (Aljamal, 2011).
It has been suggested that the thiobarbituric acid reactive substance (TBARS) content in eggs is lower in hens that were supplemented with extra Se as a result of the increased glutathione (GSH) content and GSH-Px activity. However, the ameliorative effects of inorganic Se supplementation were mitigated during long periods of storage (Skrivan et al., 2010; Wang et al., 2010). Antioxidant stability of chicken was improved in the birds that received SY compared with those that received SS, again through higher GSH-Px, SOD, and total antioxidant capacity (TAC) (Suchy et al., 2014). Selenium is also essential for the activity of human and animal immune systems (Alhidary et al., 2015). It is necessary for the metabolism of prostaglandins, stimulation of neutrophil function, and proliferation of T and B lymphocytes (Pappas et al., 2005; Fasiangova & Borilova, 2017). A deficiency of Se affects humoral immunity response, resulting in reduced levels of the IgG and IgM antibodies. Organic forms of Se are said to be more effective than the inorganic forms in improving immune functions in broilers (Suchy et al., 2014).
In most studies that assess Se deposition in egg yolk and albumen, Se has been supplemented at 0.3 to 0.5 mg/kg of the diet for three to eight weeks (Fasiangova et al., 2017). In this study, Se was added to the diet at 1 mg/kg for a 10-week trial period to assess the effects of supplementing the diet of laying hens with various sources of Se, together with flaxseed, on egg Se deposition, yolk antioxidant status, and immune response of the birds. Haug et al. (2007) reported that a high dietary selenium dosage resulted in a higher concentration of the long-chain omega-3 fatty acids in eggs, and suggested that a high selenium intake might increase the synthesis of long-chain omega-3 fatty acids from alpha-linolenic acid (ALA) or decrease their rate of degradation. Pappas et al. (2004) found an increase in the selenium concentration of fertile eggs from broiler breeders fed fish oil. They suggested that omega-3 fatty acids may affect selenium metabolism and deposition in the embryo rather than selenium absorption from the diet. The authors could not find any reports on the effect of dietary omega-3 fatty acid on the selenium enrichment of eggs. This study was designed to investigate how diets that contained various levels of flaxseed as an omega-3 source might affect the selenium enrichment of eggs produced by layers that were fed selenium from sodium selenite, Se-enriched yeast, and hydroxy selenomethionine.
Materials and Methods
The care and use of animals for this study was approved by the Animal Welfare Committee of the University of Mohaghegh-Ardabili, Iran.
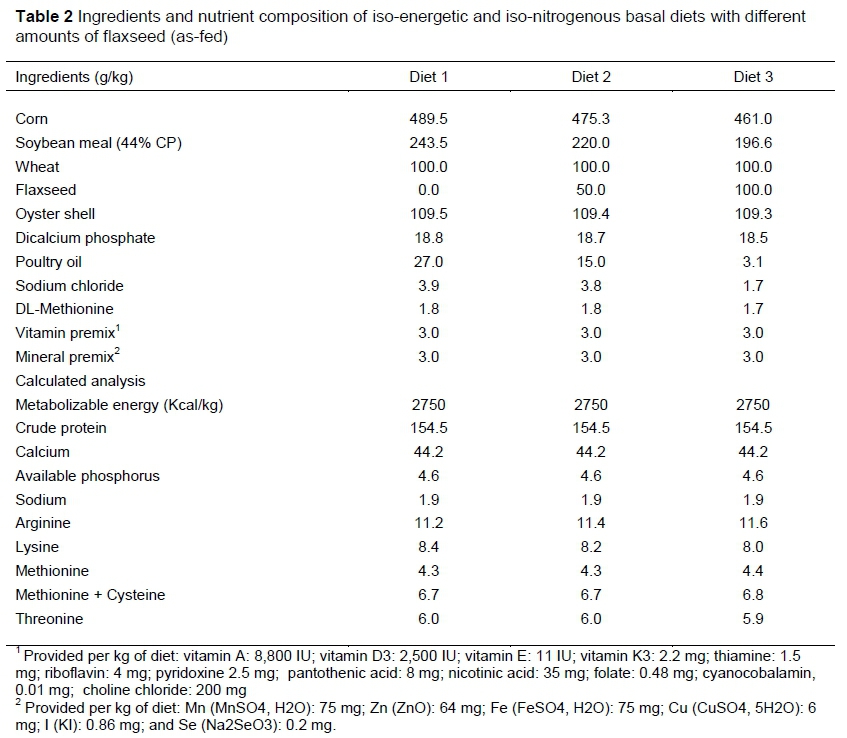
A total of 384 Hyline-W36 laying hens, aged 50 weeks, were assigned to a 3x4 factorial arrangement of treatments, comprising four replicates of eight birds for a 10-week trial. Birds were randomly placed in wire cages, so that each replicate consisted of two adjacent cages with separated feeder and four hens in each. The chemical composition of flaxseed in this experiment is presented in Table 1. Three iso-energetic and iso-nitrogenous basal diets, containing 0 g/kg, 50 g/kg and 100 g/kg of flaxseed, were formulated to meet the nutritional requirements of hens based on Hyline-W36 commercial management guides (Table 2). The basal diets, which contained 0.2 mg/kg Se from a mineral premix, were augmented with no additional Se, with 1 mg/kg of Se from SS, with 1 mg/kg of Se from SY (Sel-Plex, Alltech Int.) and with 1 mg/kg of Se from HMSeBA or hydroxy-selenomethionine (SOH) (Selisseo, Adisseo Co. Ltd., Beijing, PRC). Feed and water were provided ad libitum. The temperature and relative humidity of the house were approximately 24-26 °C and 50-60%, respectively. A lighting schedule of 16 hours light and 8 hours dark was implemented.
To assess the chemical characteristics, three eggs were collected randomly from each replicate at the end of the fifth and tenth weeks of the experiment to determine their Se concentration and in the tenth week for other characteristics. Egg yolk and egg albumen of the samples were separated using a manual egg yolk separator. The yolks and the albumen from the same replicate were pooled separately, homogenized well using a blender, and stored in plastic micro-tubes at -20 °C until analysed. To determine the Se concentration in the eggs, the samples were dried at 65 °C for 12 hours and ground. Briefly, 0.5 g of egg yolk or egg albumen was digested in a mixture of 1 ml HNO3 and 9 ml deionized water until the solution was clear. Then, the solution was diluted with deionized water to a final volume of 25 ml. Se concentration of the sample was determined using an inductively coupled plasma-mass spectrometer (ICP/MS, Agilent 7500cx, Tokyo, Japan).
The homogenized yolks that had been stored were analysed at the end of the experiment to assess lipid peroxidation and antioxidant status. TBARS analysis was performed with some modification to determine MDA level. TBA levels were expressed as milligrams of MDA per kilogram of yolk (Salih et al., 1987). 1,1 diphenyl-2-picrylhydrazyl (DPPH) radical-scavenging activity as inhibition concentration (IC%) was determined to evaluate the TAC of yolks (Brand-Williams et al., 1995). The assay of SOD activity was performed using the method of inhibition of epinephrine auto-oxidation in alkaline medium and measuring the absorbance of the resulting product at 340 nm. The result was expressed as a value of unit per gram of extracted protein (U/g) (Wawrzykowski & Kankofer, 2011). Glutathione peroxidase activity in the yolk was measured indirectly by a coupled reaction with glutathione reductase. Glutathione peroxidase catalyses oxidation of glutathione (GSH) in the presence of tert-Butyl hydroperoxide as a substrate. Oxidized glutathione is then reduced in the presence of glutathione reductase and nicotinamide adenine dinucleotide phosphate (NADPH), and NADPH is oxidized to create NADP. The oxidation process is accompanied by a decrease in light absorbance at 340 nm. This GSH-Px activity was expressed as unit per gram of extracted protein (U/g) (Paglia & Valentine, 1967; Aljamal, 2011).
SRBC were used as an antigen to assess humoral immune response. At the end of the experiment, two hens from each replicate were randomly selected and injected intravenously (brachial vein) with 0.2 ml 9% SRBC diluted in PBC. After one week, 2 ml of blood samples were taken from each bird. The blood was then centrifuged (2500 ÷ g, 10 min, 17 °C). The serum was used to determine the antibody level against SRBC by the hemagglutination test (Ambrose & Donner, 1973; Heckert et al., 2002). IgG and IgM were also measured according to the SRBC method and microtiter was done using mercaptoethanol for agglutination (Delhanty & Solomon, 1966; Isakov et al., 2005).
Cell-mediated immune response was measured as the reaction to 2,4-dinitrochlorobenzene (DNCB). At the end of the experiment, two birds from each replicate were randomly selected and 0.1 ml DNCB was applied to an approximately 10 cm2 area of skin without feathers on the left lateral abdomen. The corresponding area on the right side was treated solely with the solvent (a mixture of acetone and olive oil). Skin thickness on both sides was measured with a digital calliper before and 24 hours after the challenge. The increase in skin thickness was calculated as the difference before and after the challenge (Verma et al., 2004).
Data related to immune response and yolk antioxidant properties were analysed by a general linear model (GLM) procedure for a completely randomized experimental design using SAS software (Version 9.1; SAS Institute, Inc.). The Se concentration of the yolk and albumen were analysed as repeated measures using the MIXED procedure of SAS. Means were compared using Tukey's Test. Effects were considered significant at P < 0.05.
Results
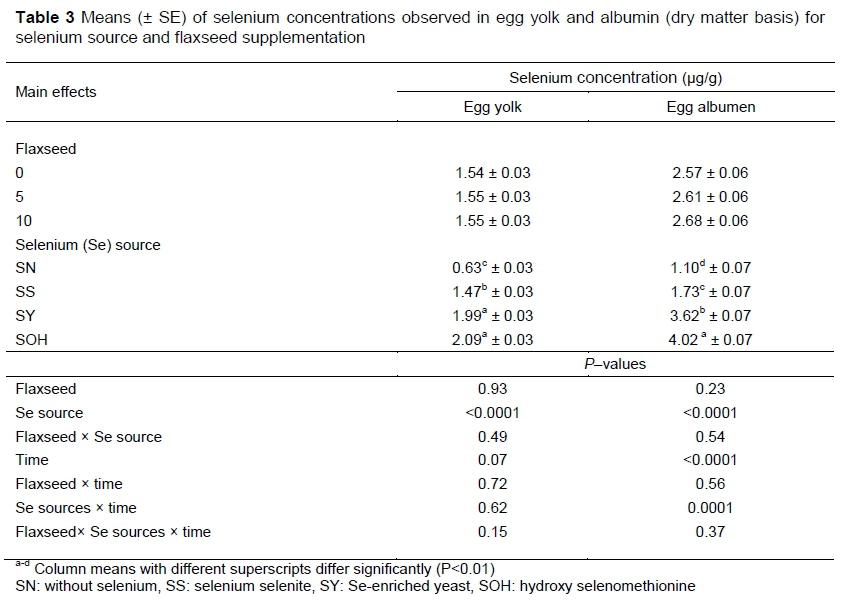
The Se concentrations of egg yolks and albumen are summarized in Table 3. Because no interactions were found between Se source and flaxseed, only the main effects are presented. In addition, these traits were not affected by flaxseed level in the diets. However, the Se dietary treatments resulted in significantly different Se concentration in egg. The greatest concentration of Se in the yolk was observed with diets containing SOH and SY, which were different from SS (P <0.01). The albumen Se concentration in the albumen was greatest in response to the diet supplemented with SOH, followed by SY and then SS (P <0.01). Thus, feeding the hens SS, SY and SOH caused about 133%, 215% and 231% relative increases in the Se content in yolks, respectively. Similarly, the relative increases in Se concentrations in the albumen were 57%, 229%, and 265%, respectively.
Antioxidant indices at the end of the trial are shown Table 4. Glutathione peroxidase activity was increased by supplementation of Se. Birds fed SOH and SY had increased TAC compared with birds that had not received supplemental Se (P<0.01). SOD activity and MDA level were not affected by Se source. Nor was any effect of flaxseed observed. Neither were interactions found between flaxseed and Se source.
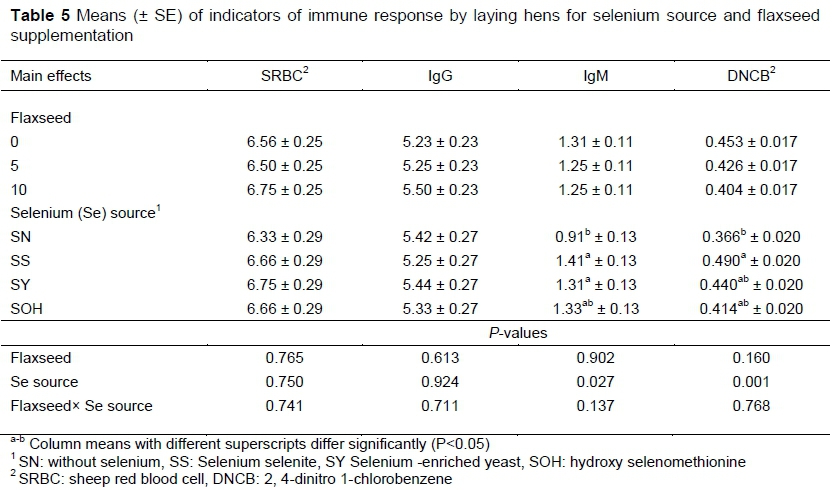
Indicators of the immune response of the hens at the end of the trial are shown in Table 5. The interactions between the level of flaxseed in the diets and sources of Se and the main effect of flaxseed were not significant. Antibody response against SRBC was not affected, except IgM was greater in birds fed diets containing SS and SY than those fed SN (P<0.05). Based on DNCB, birds that were provided with supplemental SS had increased cell-mediated response when compared with birds that received no supplemental Se (P<0.01). Interactions between flaxseed and Se source were not observed.
Discussion
The results support the general conclusion that Se concentration in eggs is a consequence of both dietary level and form of Se. Some reports suggested that Se sources such as SY produce greater deposition of Se in the whole egg compared with SS (Chinrasri et al., 2009; Briens et al., 2013; Fasiangova et al., 2017). It is believed that organic sources have a higher absorption efficiency compared with SS, with 50-75% of it being excreted (Hu et al., 2012). Chinrasri et al. (2009) reported that organic Se-Met is actively absorbed from the gut, and can be incorporated directly into body tissues and in products such as eggs as effectively as methionine, whereas inorganic forms are absorbed passively and are probably required for Se-Cys synthesis. Inorganic Se compounds are used primarily in the Se metabolism pathway to synthesize seleno-protein and not to replenish Se deposits in existing tissues.
In this study, organic Se sources improved the Se deposition in the egg yolk and albumen. The elative increase of Se in the eggs produced by hens that consumed SOH was greater than that of those that consumed SY. In fact, SOH is a relatively new organic Se source, with increased bioavailability compared vith SY and SS, which has been introduced to the animal feed industry. Also, the increased efficacy of SOH may arise from its greater purity (99%) as a precursor of Se-Met, compared with SY, which contains about 60-65% Se in the form of Se-Met, and the remaining 37% is bound in other forms or as inorganic Se (Jlali et al., 2014). In fact, organic Se is mostly retained in the body, but there is a high excretion of inorganic Se through the gut and from the tissues (Rajasekaran & Kalaivani, 2015).
It has been shown that SOH influences the egg yolk quality positively through providing a greater Se supply to the egg (Tufarelli et al., 2015). Previous researchers demonstrated that the organic and inorganic forms of Se were deposited primarily in egg albumen and yolks, respectively (Fasiangova et al., 2017; Pappas et al., 2006). Together with the results of the present study, the previous research suggested that SOH had a remarkable tendency towards egg Se deposition, especially in the albumen. However, relatively greater Se enrichment of the yolk relative to albumen has been reported from SY (Pappas et al., 2006).
Gurbuz et al. (2012) provided SY and SS to hens at 0.3 mg/kg diet, and observed that Se content of the eggs produced by hens that were given SY was greater on the 60th and 90th days of the experiment, with no difference being observed the 30th day. Kralik et al. (2016) showed that albumen Se content was affected by supplementation with a low level of Se-Met by the 26th day of the experiment, but the yolk was not affected at that time. It has also been shown that both the concentration and form of dietary Se can influence Se content of the yolk, so that organic sources supplemented at a higher level (0.6 ppm compared with 0.3 ppm) resulted in greater deposition (Asadi et al., 2017). In confirmation of results from Jlali et al., 2014, the present results show that Se deposition in the yolk and albumen increases 3- to 3.5-fold as a consequence of supplementation with organic sources, while SS increased Se deposition in the yolk more than in albumen (approximately 2.5-fold versus1.5-fold increase rate).
Dalle-Zotte et al. (2015) reported that malondialdehyde (MDA) index increased in birds that were fed omega-3 PUFA sources such as flaxseed. However, it has been reported that omega-3 PUFA enhances the antioxidative status, such as GSH-Px activity and TAC, and decreases lipid peroxidation in the serum of laying hens (Ebeid, 2011). Further, flaxseed-augmented diets that were fed to geese improved liver SOD and GSH-Px in their progeny (Chen et al., 2015). Increased dietary omega-3/omega-6 PUFA may increase antioxidant capacity and alleviate inflammation by inhibiting synthesis of omega-6 PUFA (Chen et al., 2015). However, no effect was observed in the current study of dietary n-3 PUFA from flaxseed on the antioxidant indices in the yolk and plasma of the birds. This study was conducted with fresh eggs, so possibly changes might become apparent only after a period of storage, during which more peroxidation occurs in the yolk.
Contrary to other studies (Mohiti-Asli et al., 2008; Wang et al., 2010, Fasiangova & Borilova, 2017) that indicated effects of Se supplementation on the antioxidant content of serum or yolk, in this study neither MDA nor SOD responded differently to the Se sources. However, the present results are consistent with the findings of other researchers (Petrovic et al., 2006; Lesson et al., 2008). In contrast, TAC and GSH-Px increased when Se was supplemented in this study. Among indices of antioxidant status in biological systems, TAC offers an integrated measure of enzymatic and non-enzymatic antioxidants, and it is assumed to be a better index of biological function than a single antioxidant (Mahmoud & Hijazi, 2007). The results from the current study for TAC are consistent with the effect of Se supplementation, especially from organic sources, enhancing antioxidant response. According to Boostani et al. (2015), antioxidant enzyme activities such as GSH-Px act as a protective response to oxidative stress in broilers and increase when the diet is supplemented with Se. In addition, Se supplementation can lower the level of MDA. However, Buckiuniene et al. (2018) reported that MDA content in yolks is greater when the diet is supplemented with Se-Met. A plausible explanation for these differences among studies may be that when birds are not exposed to sufficient stress, there is less opportunity for Se supplementation to be effective.
The lack of differences in GSH-Px that were observed in this study may result from both the organic and inorganic sources being converted to Se-Cys, which, in turn, leads to synthesizing additional seleno-protein enzymes (Papazyan et al., 2006). Rajashree et al. (2014) found the activity of GSH-Px in the albumen and yolk to be greater with dietary organic Se compared with Se from an inorganic source. However, some researchers reported that GSH-Px activity in laying hens was not affected by the source and dose of Se (Lesson et al., 2008; Delezie, 2014). It has also been observed that SS may be regarded as a pro-oxidant, which can stimulate lipid peroxidation, and thus decreases the absorption of various nutrients such as antioxidants (Papazyan et al., 2006; Skrivan et al., 2010).
In avian species, SOD and GSH-Px activities that function in cell adaptation to stress conditions such as oxidative stress and metabolic pressure can be influenced by dietary Se levels and forms (Chen & Wu, 2014; Surai, 2016). Greater SOD and GSH-Px activities and TAC and reduced MDA content were found in plasma of laying hens fed SY and Se-Met compared with hens fed SS (Chen & Wu, 2014; Jing et al., 2015; Surai, 2016). Furthermore, Se supplementation increases SOD activity and TAC in the serum of laying hens, probably owing to the synergy of Se with vitamins E and A (Zdunczyk et al., 2013). In agreement with the present results, Jing et al. (2015) observed that organic Se sources have greater ability to increase GSH-Px activity in the albumen and yolk, compared with SS. However, Jing et al. (2015) observed no difference in SOD activity between organic Se and SS. The current results confirm the findings of Petrovic et al. (2006), who demonstrated that GSH-Px activity in various tissues depended on there being Se in the diet rather than on the source or amount of supplementation.
It has been suggested that Se sources, especially organic ones, and PUFA may interact. This interaction may occur through Se-dependent GSH-Px, which plays an important role in regulating the biosynthesis of prostaglandins from their precursor eicosanoids. However, the precise nature of this role is not fully understood (Mohiti-Asli et al., 2008). No interactions between flaxseed and Se source were observed in this study that affect GSH-Px.
Diets that are high in omega-3 PUFA reportedly increase serum IgG of hens and egg IgY, with the dietary ratio of omega-6/omega3 PUFA being the main factor in modulating IgG formation (Ebeid, 2011; Cherian & Quezada, 2016). The yolks of eggs from hens that were fed flaxseed oil or ground flaxseed have also been shown to be enriched in omega-3 fatty acids. (Ehr et al., 2017). Enriching eggs with omega-3 PUFA may enhance their antioxidant status and reduce lipid peroxidation, which may improve the immune response in laying hens. The protective effect of dietary omega-3 fatty acids from sources such as fish oil may be associated with EPA as a metabolite of arachidonic acid (Huang, 2016). The ratio of omega-6/omega3 PUFA can also influence cell-mediated immunity, but not the antibody response in broiler breeders (Khatibjoo et al., 2011). Fu et al. (2017) indicated that greater levels of linoleic acid, as the main omega-6 fatty acid in the diet, can undermine the immune response, while higher levels of omega-3 sources such as fish oil enhance the antibody titre, especially IgM in broilers. Although these references provide a good body of evidence that exhibits the effect of flaxseed on the functionality of immunity in poultry, no such response was recorded in this study.
Selenium is known to be vital for stimulation of neutrophil function, as well as the functioning of T and B lymphocytes in humans and animals. In fact, GSH-Px acts in cellular redox and other physiological reactions such as signal transduction and regulation of pro-inflammatory cytokine production (Pappas et al., 2005; Fasiangova & Borilova, 2017). It was demonstrated that GSH-Px activity can improve immune response through the production of antibody titre in aged laying hens, as a result of its antioxidant properties. In addition, GSH-Px tends to protect neutrophils from oxygen-derived radicals (Ebeid, 2011). It was also indicated that the interaction of Se and vitamin E affects antibody production against SRBC, including the total IgG and IgM in laying hens (Ziaei et al., 2013). Although the main cause of this influence is unknown, it may be due to their antioxidant properties.
It has been suggested that the insufficiency of Se affects humoral immune response, which is manifest as reduced levels of IgG and IgM antibodies, and that organic forms of supplemental Se are more effective than inorganic ones in improving immune functions in broilers (Suchy et al., 2014). Likewise, dietary supplementation of nano-Se increased humoral and cellular immune responses compared with SS and control groups in growing layers (Mohapatra & Swain, 2014). According to Boostani et al. (2015), oxidative conditions can decrease the weights of lymphoid organs and the negative effect may be attenuated by dietary supplementation of Se. They observed increased levels of IgG and IgM from supplementation with either organic Se or nano-Se. In contrast, Mohiti-Asli et al. (2010) observed that neither SY nor SS could alleviate the low level of SRBC caused by heat stress in laying hens.
The results of this study, for the most part, agree with the existence of an effect of Se supplementation on cellular immune function. Humoral immunity, contrary to these studies, total SRBC and IgG remained unchanged in the present study and only IgM was positively affected. Thus for humoral immunity, organic and inorganic Se forms were equally effective. This result confirms the findings by Boostani et al. (2015) that organic Se can result in higher serum IgM concentration in broilers. The presence of Se, irrespective of source, is an important factor in immune function and organic and inorganic forms have the same bioavailability.
Conclusion
No interactions were observed between dietary selenium sources and flaxseed levels that affected the selenium content in eggs, antioxidant status of egg yolk, and immune response of the laying hens. Nor were any effects of adding up to 100 g/kg flaxseed to the diet observed for these traits. However, dietary inclusion of Se at a greater level (1 mg/kg diet) than is normally used could improve Se deposition in the egg without considerable adverse effects. Organic sources, especially hydroxyl Se-Met, produce greater reserves in eggs, particularly in albumen, so it may be a better choice for enrichment of eggs. Selenium supplementation may also improve antioxidant status and some components of immune response.
Acknowledgements
The authors would like to thank Arona Co., Javaneh Khorasan Co., Tehran Agriculture Research & Education Center, and the University of Mohaghegh-Ardabili for the technical support in the conduct of this study. They also appreciate the assistance provided by Mr. Majid Afshar and Ms. Nahid Hassanabadi.
Authors' Contributions
All of the authors cooperated in implementing the trial and statistical analysis as well as interpreting and reviewing the results of the study.
Conflict of Interest Declaration
The authors declare that they have no conflict of interests.
References
Alhidary, I.A., Shini, S,A.l., Jassim, R.A.M., Abudabos, A.M. & Gaughan, J.B., 2015. Effects of selenium and vitamin E on performance, physiological response, and selenium balance in heat-stressed sheep. J. Anim. Sci. 93, 576-588. [ Links ]
Aljamal, A.A., 2011. The effect of vitamin E, selenomethionine, and sodium selenite supplementation in laying hens. Dissertation, University of Nebraska-Lincoln. Lincoln, USA. [ Links ]
Ambrose, C.T. & Donner, A., 1973. Application of the analysis of variance to hemaglutination titration. J. Immunolo. Meth. 3,165-210. [ Links ]
Asadi F., Shariatmadari, F., Karimi-Torshizi, M.A., Mohiti-Asli M. & Ghanaatparast-Rashti M., 2017. Comparison of different selenium sources and vitamin E in laying hen diet and their influences on egg selenium and cholesterol content, quality and oxidative stability. Iran. J. App. Anim. Sci. 7, 83-89. [ Links ]
Boostani A., Sadeghi A.A., Mousavi, S.N., Chamani, M. & Kashan, N., 2015. Effects of organic, inorganic, and nano-Se on growth performance, antioxidant capacity, cellular and humoral immune responses in broiler chickens exposed to oxidative stress. Livestock Sci. 178, 330-336. [ Links ]
Brand-Williams, W., Cuvelier, M.E. & Berset, C., 1995. Use of a free radical method to evaluate antioxidant activity. Lebensmittel-Wissenschaft & Technologie. 28, 25-30. [ Links ]
Briens, M., Mercier, Y. & Geraert. P.A. 2013. Comparative study of a new organic selenium source v. seleno-yeast and mineral selenium sources on muscle selenium enrichment and selenium digestibility in broiler chickens. Br. J. Nut. 110, 617-624. [ Links ]
Buckiuniene, V., Alencikiene, G., Miezeliene, A., Raceviciute-Stupeliene, A. & Bliznikas, S., 2018. Effect of sunflower and rapeseed oil, organic and inorganic selenium and vitamin E in the diet on yolk fatty acids profile, malondialdehyde concentration and sensory quality of laying hens' eggs. Veterinarija IR Zootechnika. 76(98): 21-28. [ Links ]
Chen, G., Wu, J. & Li, C., 2014. Effect of different selenium sources on production performance and biochemical parameters of broilers. J. Anim. Physiol. Anim. Nutr. (Berl). 98,747-754. [ Links ]
Chen, W., Jiang, Y.Y., Wang, J.P., Yan, B.X., Huang, Y.Q. & Wang, Z. X., 2015. Effect of flaxseed on the fatty acid profile of egg yolk and antioxidant status of their neonatal offspring in Huoyan geese. Animal. 9, 1749-1755. [ Links ]
Cherian, G. & Quezada, N., 2016. Egg quality, fatty acid composition and immunoglobulin Y content in eggs from laying hens fed full camelina or flax seed. J. Anim. Sci. Biotechnol. 7,1-8. [ Links ]
Chinrasri, O., Chantiratikul, P., Thosaikham, W., Atiwetin, P., Chumpawadee, S., Saenthaweesuk, S. & Chantiratikul, A., 2009. Effect of selenium-enriched bean sprout and other selenium sources on productivity and selenium concentration in eggs of laying hens. Asian-Austra. J. Anim. Sci. 22, 1661-1666. [ Links ],
Delezie, E., Rovers, M., Vander, A., Ruttens, A., Wittocx, S. & Segers. L., 2014. Comparing responses to different selenium sources and dosages in laying hens. Poult. Sci. 93,3083-3090. [ Links ]
Delhanty, J.J. & Solomon, J.B., 1966. The nature of antibodies to goat erythrocytes in the developing chicken. Immunology. 11,103-113. [ Links ]
Dalle Zotte, A., Andrighetto, I., Giaccone, V. & Marchesini, G., 2015. Dietary enrichment of n-3 PUFA for laying hens: effect of different sources on production, composition and quality of eggs. Anim. Sci. Papers. Reports. 4,411-424. [ Links ]
Ebeid, T.A., 2011. The impact of incorporation of n-3 fatty acids into eggs on ovarian follicular development, immune response, antioxidative status and tibial bone characteristics in aged laying hens. Animal. 5,1554-1562. [ Links ]
Ehr, I.J., Persia, M.E. & Bobeck, E.A., 2017. Comparative omega-3 fatty acid enrichment of egg yolks from first-cycle laying hens fed flaxseed oil or ground flaxseed. Poult. Sc. 96, 1791-1799. [ Links ]
Fasiangova, M. & Borilova, G., 2017. Impact of Se supplementation on the oxidation stability of eggs. World. Poult. Sci. J. 73, 175-184. [ Links ]
Fasiangova, M., Borilova, G. & Hulankova, R., 2017. The effect of dietary Se supplementation on the Se status and physico-chemical properties of eggs - a review. Czech. J. Food. Sci. 35, 275-284. [ Links ]
Fu, S., Guo, S. & Shen Z., 2017. Effects of PUFA on immune response of Avian. J. Chem. Pharma. Res. 9, 334-337. [ Links ]
Gurbuz, E., Balevi, T., Coskun, B. & Baris Citil, O., 2012. Effect of adding linseed and selenium to diets of layer hens on performance, egg fatty acid composition and selenium content. Kafkas. Univ. Vet. Fak. Derg. 18, 484-496. [ Links ]
Haug, A., Eich-Greatorex, E., Bernhoft, A., Wold, J.P., Hetland, H., Christophersen, O.A & Sogn, T. 2007. Effect of dietary selenium and omega-3 fatty acids on muscle composition and quality in broilers. Lipids. Health. Dis. 6, 29. [ Links ]
Heckert, R.A., Estevez, I., Russek-Cohen, E. & Pettit-Riley, R., 2002. Effects of density and perch availability on the immune status of broilers. Poult. Sci. 81, 451-457. [ Links ]
Hu, C.H., Li, Y.L., Xiong, L., Zhang, H.M., Song, J. & Xia, M.S., 2012. Comparative effects of nano elemental selenium and sodium selenite on selenium retention in broiler chickens. Anim. Feed. Sci. Technol. 177, 204-210. [ Links ]
Huang, S., 2016. Effects of feeding extruded flaxseed on layer performance. Master's thesis, McGill University, Montreal, Quebec, Canada. [ Links ]
Isakov, N., Feldmann, M. & Segel, S., 2005. The mechanism of modulation of humoral immune responses after injection of mice with SRBC. J. Immunol. 128, 969-975. [ Links ]
Jing, C.L., Dong, X. F., Wang, Z.M., Liu, S. & Tong, J.M., 2015. Comparative study of DL-selenomethionine vs sodium selenite and seleno-yeast on antioxidant activity and selenium status in laying hens. Poult. Sci. 94, 965-75. [ Links ]
Jlali, M., Briens, M., Rouffi, F., Mercerand, F., Geraert, P.A. & Mercier, Y., 2014. Effect of 2-hydroxy-4- methylselenobutanoic acid as a dietary selenium supplement to improve the selenium concentration of table eggs. J. Anim. Sci. 91,1745-1752. [ Links ]
Khatibjoo, A., Kermanshahi, H., Golian, A. & Zaghari, M., 2011. The effect of dietary n-6:n-3 ratio and sex on broiler breeder immunity. Poult. Sci. 90, 2209-2216. [ Links ]
Kralik, Z., Grcevic, M., Radisic, Z., Kralik, I., Loncaric, Z. & Skrtic, Z., 2016. Effect of selenium-fortified wheat in feed for laying hens on table eggs quality. Bulg. J. Agri. Sci. 22, 297-302. [ Links ]
Leeson, S., Namkung, H., Caston, L., Durosoy, S. & Schlegel, P., 2008. Comparison of selenium levels and sources and dietary fat quality in diets for broiler breeders and layer hens. Poult. Sci. 87, 2605-2612. [ Links ]
Mahmoud, K.Z. & Hijazi, A.A., 2007. Effect of vitamin A and/or E on plasma enzymatic antioxidant systems and total antioxidant capacity of broiler chickens challenged with carbon tetrachloride. J. Anim. Physiol. Nut. 91, 333-340. [ Links ]
Miranda, J. M., Anton, X., Redondo-Valbuena, C. & Roca-Saavedra, P., 2015. Egg and egg-derived foods: Effects on human health and use as functional foods. Nutrients. 7, 706-729. [ Links ]
Mohapatra, P., Swain, SK., Mishra, T., Behera., P., Swain, N.C., Dhama, K., 2014. Effects of dietary nano-selenium supplementation on the performance of layer grower birds. Asian. J. Anim. Vet. Adv. 9, 641-652. [ Links ]
Mohiti-Asli, M., Shariatmadari, F., Lotfollahian, H., Mazuji, M.T., 2008. Effects of supplementing layer hen diets with selenium and vitamin E on egg quality, lipid oxidation and fatty acid composition during storage. Can. J. Anim. Sci. 88,475-483. [ Links ]
Mohiti-Asli, M., Shariatmadari, F. & Lotfollahian, H., 2010. The influence of dietary vitamin E and selenium on egg production parameters, serum and yolk cholesterol and antibody response of laying hen exposed to high environmental temperature. Arch. Gaflugelk. 74, 43-50. [ Links ]
Paglia, D.E. & Valentine, W.N., 1967. Studies o the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 70, 158-167. [ Links ]
Papazyan, T.T., Denev, S.A. & Surai, P.F., 2006. Selenium in poultry nutrition: Lessons from research and wild nature. Krmiv,. Zagreb 5, 275-283. [ Links ]
Pappas, A.C., McDevitt, R.M., Surai, P.F., Acamovic, T. & Sparks, N.H.C., 2004. The effects of selenium and PUFA supplementation in the diet of young broiler breeders on the incorporation of selenium in the egg and in the tissues of the day-old broiler chick. Spring meeting of the WPSA UK branch paper., Br. Poult. Sci. 45:sup1, S26-S27. [ Links ]
Pappas, A.C., Acamovic, T., Sparks, N.H., Surai, P.F. & McDevitt, R.M., 2005. Effects of supplementing broiler breeder diets with organic selenium and polyunsaturated fatty acids on egg quality during storage. Poult. Sci. 84, 865-874. [ Links ]
Pappas, C., Acamovic, T., Sparks, N.H.C., Surai, P.F. & McDevitt, R.M., 2006. Effects of supplementing broiler breeder diets with organoselenium compounds and polyunsaturated fatty acids on hatchability. Poult. Sci. 85,1584-1593. [ Links ]
Petrovic, V., Bodizarova, K., Faix, S., Mellen, M., Arpasova, H. & Leng. L., 2006. Antioxidant and selenium status of laying hens fed with diets supplement with selenite or Se-yeast. J. Anim. Feed. Sci. 15, 435-444. [ Links ]
Rajasekaran, A. & Kalaivani M., 2015. Designer foods and their benefits: A review. J. Food. Sci. Technol. 50,1-16. [ Links ]
Rajashree, K., Muthukumar, T. & Karthikeyan. N., 2014. Comparative study of the effects of organic selenium on hen performance and productivity of broiler breeders. Br. Poult. Sci. 55, 367-374. [ Links ]
Salih, A.M., Smith, D.M., Price, J.F. & Dawson, L.E., 1987. Modified extraction 2-thiobarbituric acid method for measuring lipid oxidation in poultry. Poult. Sci. 66,1483-1488. [ Links ]
Skrivan, M., Bubancova, I., Marounek, M. & Dlouha, G., 2010. Selenium and á-tocopherol content in eggs produced by hens that were fed diets supplemented with selenomethionine, sodium selenite and vitamin E. Czech. J. Anim. Sci. 55, 388-397. [ Links ]
Suchy, P., Strakova, E. & Herzig, I., 2014. Selenium in poultry nutrition: A review. Czech. J. Anim. Sci. 59, 495-503. [ Links ]
Surai, P.F. & Fisinin, V.L., 2012. Feeding breeders to avoid oxidative stress in embryos. XXIV World Poultry Congress Proceedings, WPSA, Salvador, Brazil. Chicken Breeder and Broiler Production 1-12. [ Links ]
Surai, P.F. & Fisinin, V.I., 2014. Selenium in poultry breeder nutrition: An update. Anim. Feed. Sci. Technol. 191, 1-15. [ Links ]
Surai, P.F. 2016. Antioxidant system in poultry biology: Superoxide dismutase - Review article. J. Anim. Res. Nut. 1, 117. [ Links ]
Tufarelli, V., Ceci, E. & Laudadio, V., 2015. 2-Hydroxy-4-methylselenobutanoic acid as new organic selenium dietary supplement to produce selenium-enriched eggs. Biol. Trace. Elem. Res. 171,453-458. [ Links ]
Verma, J., Johri, T.S., Swain, D.B.K. & Ameena, S., 2004. Effect of graded levels of aflatoxin and their combination on the performance and immune response of broilers. Br. Poult. 45, 512-518. [ Links ]
Wang, Z.G., Pan, X.J., Zhang, W.Q. & Peng, Z.Q., 2010. Methionine and selenium yeast supplementation of the maternal diets affects antioxidant activity of breeding eggs. Poult. Sci. 89, 931-937. [ Links ]
Wawrzykowski, J. & Kankofer, M. 2011. Changes in activity during storage and characteristics of superoxide dismutase from hen eggs (Gallus gallus domesticus). Eur. Food. Res. Technol. 232, 479-484. [ Links ]
Zambiazi, R.C., Przybylski, R. & Zambiazi, M.W., 2007. Fatty acid composition of vegetable oils and fats - B. CEPPA, Curitiba 25, 111-120. [ Links ]
Zdunczyk, Z., Drazbo, A., Jankowski, J., Juskiewicz, J., Antoszkiewicz, Z. & Troszynska, A., 2013. The effect of dietary vitamin E and selenium supplements on the fatty acid profile and quality traits of eggs. Archiv. Tierzuch. 56, 719-732. [ Links ]
Ziaei, N., Moradi-Kor, N. & Esfandiar Pour, E., 2013. The effects of different levels of vitamin E and organic selenium on performance and immune response of laying hens. Afr. J. Biotechnol. 12, 3884-3890. [ Links ]
Received 12 November 2018
Accepted 18 August 2019;
First published online 17 September 2019
# Corresponding author: bnavidshad@uma.ac.ir


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}