










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 no.4 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i4.11
ARTICLES
PCR-RFLP-based identification of polymorphisms in BMPR1B, GDF9 and BMP15 genes associated with litter size in Beetal and Teddy goats
M. IslamI; A. BasheerI, #; K. JavedI; A.A. AnjumII; I. ZahoorI
IAnimal Breeding and Genetics, Department of Livestock Production, University of Veterinary and Animal Sciences, Lahore, Pakistan
IIDepartment of Microbiology, University of Veterinary and Animal Sciences, Lahore, Pakistan
ABSTRACT
The genes BMPR1B, GDF9 and BMP15 are oocyte-derived members of the transforming growth factor-β superfamily, which is essential for follicular growth and ovulation. The aim of the current study was to determine the incidence of reported mutations in exon 3 of BMPR1B gene, exon 1 of GDF9 gene, and exon 2 of BMP15 gene and their association with litter size in three parities of Beetal and Teddy goats. Based on the known mutations in these genes, PCR primers were designed to find out the polymorphism through the PCR-RFLP technique in 120 randomly selected animals of the two breeds, which vary in their prolificacy. The current results show that there were significant differences in litter size [1.708 ± 0.060 in Beetal (B) and 2.167 ± 0.056 in Teddy (T) goats] in both breeds. The results of breed x parity interaction showed significant differences in litter size in first [1.158 ± 0.061 (B); 1.861 ± 0.057 (T)], third [2.062 ± 0.078 (B); 2.583 ± 0.073 (T)] and average parity [708 ± 0.060 (B); 2.167 ± 0.056 (T)] in both breeds. All three loci in both breeds were found to be polymorphic and the results of breed x parity x genotype analysis showed that genotypes of all the three genes were significantly associated with litter size in all parities in both breeds. For the BMPR1B gene, heterozygous (CT) animals of Beetal and Teddy goats had the largest litter size in all three parities and the average (parity) litter size, showing the overdominance of the heterozygotes. Similarly, for GDF9 gene, heterozygous animals (AG) had the largest litter size in both breeds. For BMP15 gene homozygous carrier animals (CC) of both breeds had the highest number of kids, followed by the heterozygous (AC) genotypes. The current results show the importance of BMPR1B, GDF9 and BMP15 as the major genes that influence prolificacy in both Beetal and Teddy goats. It is suggested that these polymorphisms can be used as molecular markers to select the prolific animals in these breeds.
Keywords: Beetal goats, Teddy goats, molecular markers, prolificacy, polymorphism
Introduction
Goats are considered 'future animals' for rural and urban prosperity (Ahlawat et al., 2016) owing to their ability to tolerate hot climatic conditions (Khan et al., 2013), their better productivity (Kumar et al., 2016), their small size, their ability to utilize poor quality roughages (Ahlawat et al., 2015b) and their non-competitiveness with human beings for food (Shi et al., 2010). The goat population is highest in Asia (55%), followed by Africa (39%) (FAOSTAT, 2016), which is known to have the lowest per capita GDP in the world (FAOSTAT, 2016). Hence, it is anticipated that goat populations could be helpful in improving the livelihoods of livestock-keeping rural household on these continents. According to Food and Agricultural Organization (FAO), about 73% more meat and 58% more dairy products need to be produced by 2050 to feed the growing human population, which is expected to increase to 9.6 billion from 7.2 billion (FAOSTAT, 2016). This increase in population would result in further shortages of grazing and water resources, which are already at risk owing to the threat of global warming (Ahlawat et al., 2015b; Rojas-Downing et al., 2017). The gap between demand and production of quality animal protein could be bridged by selecting goat genetic resources for prolificacy by employing modern genetics tools.
Animals that produce twins participate more than 1.5 times more in meat production than those that produce a single offspring per kidding (Ahlawat et al., 2016). Litter size is a sex-limited and polygenic trait and is controlled by a plethora of endocrine and paracrine mediators (Polley et al., 2009a). Ovulation rate and eventually the litter size are the main factors that determine the reproductive efficiency in small ruminants (Bradford et al., 1989; Rosatia et al., 2002). A large number of studies have been performed on various sheep breeds to identify the candidate genes that are involved in controlling the ovulation rate and prolificacy (Kumari et al., 2016). Three genes of the ovary-derived transforming growth factor β (TGFß) superfamily (Shabir M et al., 2013), including bone morphogenetic protein receptor-type 1B (BMPR1B/FecB), also called booroola gene (Chu et al., 2007); growth differentiation factor 9 (GDF9/FecG) (Feng et al., 2011); and bone morphogenetic protein 15 (BMP15/FecX) (Wang et al., 2011) have been reported to be the major candidate genes for prolificacy in sheep (Polley et al., 2009b).
BMPR1B was the first key gene that was identified for its role in controlling litter size in sheep (Montgomery et al., 1993) and a (FecBB) mutation in this gene was reported in the Boorola Merino (Wilson et al., 2001) and Javanese sheep (Davis et al., 2002). It is a substitution mutation (Q249R) in the intracellular kinase signalling domain of the BMPR1B gene which was found to be associated with hyper-prolific phenotypes of Booroola sheep (Fogarty, 2009; Hua et al., 2009). This co-dominant autosomal locus is involved mainly in the growth and development of ovarian follicles (Polley et al., 2009b). BMP15 protein is the product of an X-linked gene that is expressed in oocytes in mammals, and it helps to stimulate follicular development (Hanrahan et al., 2004). Heterozygous ewes that carry the naturally occurring BMP15 point mutations have enhanced rates of ovulation (Polley et al., 2009a). Several mutations in this gene have been identified, such as FecXI and FecXH from Romney sheep, FecXB and FecXG from Cambridge and Belclare (Davis, 2005; Mullen MP, 2013), FecXL from Laucane sheep (Bodin et al., 2007), FecXR from Rasa Aragonesa, FecXGr from Grivette (Demars et al., 2013) and FecXO from Olkuska sheep (Demars et al., 2013).
However, GDF9 is another major gene that is responsible for the highly prolific phenotype in sheep. It has a major role in growth and differentiation of antral follicles in female reproduction in mammals (Chu et al., 2011a). A mutation (FecGH) in GDF9 gene in heterozygous form was seen to be responsible for a high ovulation rate, but in homozygous form it causes infertility in CambridTge and BelEclare sheep (Hanrahan et al., 2004). Likewise, other mutations in this gene, which include FecGT and FecGE, had been observed in Icelandic (Davis, 2005) and Santa Inês sheep (Demars et al., 2013), respectively,.
Some recent studies have reported the association of genetic variation in BMP15 (Ahlawat et al., 2016; Wang et al., 2011) and GDF9 (Chu et al., 2011a; Chu et al., 2011b; Feng et al., 2011) with litter size in some goat breeds. Beetal is a heritage goat, and the pride of Pakistan and Indian (eastern) Punjab, and is used mainly for milk and meat (Khan et al., 2008), while the Teddy is known for its ability to survive in severe environmental conditions, its fast growth rate, and high prolificacy (Khan et al., 2016), and is found in Pakistan and Bangladesh. The twinning rate in Beetal goats is 50% - 55%, whereas in Teddy it is >70% (Nawaz et al., 2013; Muhammad et al., 2015). Therefore, the present study was planned to identify the genetic variations in BMPR1B, BMP15 and GDF9 genes, which are associated with litter size, in these two goat breeds of the Indo-Pak subcontinent.
Materials and Methods
Ethical permission (number DR/152) for the collection of blood samples was obtained from the Ethical Review Committee for Animal Research of University of Veterinary and Animal Sciences, Lahore, Pakistan.
The present research was conducted on 120 animals, 60 of each of the Teddy and Beetal (60 + 60) goat breeds. A 5 mL blood sample was collected from the jugular vein of each animal with a sterilized 5 mL syringe fitted with a 22-gauge needle. Blood samples were transferred to a 50 mL falcon tube containing 160 μL EDTA, and samples were immediately stored on ice. Later, samples were stored at -20 °C for subsequent DNA extraction. The phenotypic (litter size) data for first three parities of each Beetal goat were collected from Small Ruminant Training and Research Centre (SRTC), Ravi Campus, University of Veterinary and Animal Sciences, Lahore, Pakistan, and from private breeders. Similarly, Teddy goat blood samples and phenotypic data for the first three parities were collected from Livestock Production Research Institute (LPRI), Bahadarnagar, Okara, Pakistan, and private breeders in the surrounding areas.
A volume of 100 μL of each blood sample was transferred into Eppendorf tubes. To this, 1000 μL TEB buffer (Tris HCL 10 mM, EDTA 2 mM) was added, and samples were vortexed for 5 minutes and then centrifuged at 1000 rpm for 5 minutes. A pellet was made at the bottom of the Eppendorf tube. After discarding the remaining solution, 20 μL proteinase K enzyme was added, along with 60 μL SDS (10%) solution and 100 μL TNE buffer (Tris HCL 10 mM, NaCL 400 mM, EDTA 2 mM). Samples were vortexed for 5 minutes and placed in a water bath at 58 °C for overnight digestion. PCL (phenol 25: chloroform 24: isoamyle 1) was added to the fully digested samples. Three layers were formed after centrifugation at 1000 rpm for 10 minutes. Samples were washed with 70% ethanol and again centrifuged at 1000 rpm for 10 minutes. A solid pellet of DNA was formed and dissolved in 100 μL double distilled water. DNA was stored at 4 °C or -20 °C. The concentration of DNA was measured in a Nanodrop spectrophotometer (ThermoFisher).
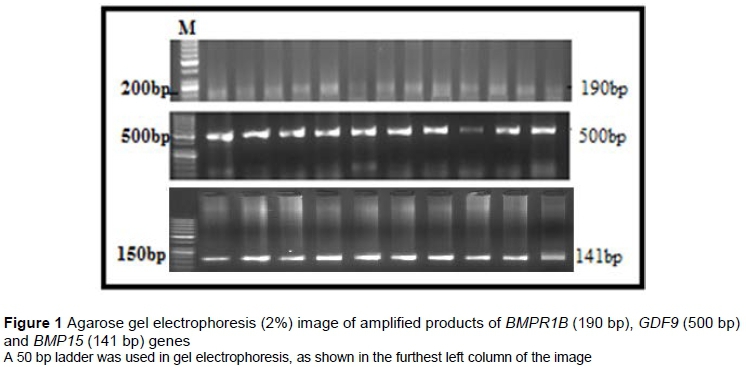
Polymerase chain reaction (PCR) was carried out in a final reaction volume of 20 μL in C-1000 Touch™ thermal cycler (Bio-Rad, USA). The primers for the amplification of BMP15, GDF9 and BMPR1B are given in Table 1. The reaction mixture consisted of 150 μΜ dNTPs, 1.2 mM MgCl2, 2.0 mM 10x buffer, 30 ng of each of forward and reverse primer and 1 unit of Taq DNA polymerase. The PCR reaction cycle protocol was 5 min at 95 °C, 30 cycles at 94 °C for 45 seconds, annealing at various temperatures (Table 1) for 30 seconds, extension at 72 °C for 45 seconds. The PCR products were visualized in Gel DocTM EZ imager (Bio-Rad, USA), following electrophoresis through 2% agarose gel (Figure 1).
The amplified regions of undigested PCR product were sequenced using an ABI3130 automated DNA sequencer (Applied Biosystems, USA). Multiple sequence alignments were performed with MEGA 7 (Sudhir et al., 2016) software using the CLUSTALW (Thompson et al., 1994) algorithm to identify polymorphisms.
PCR products of all the genes were digested with restriction enzymes (Table 1) at 37 °C for 12 hours. After digestion, the samples were stored at 4 °C. The digested products were visualized in Gel DocTM EZ imager (BIO-RAD, USA) following electrophoresis through 2% agarose gel (Adhikari et al., 2012).
The following fixed effects model was used to analyse litter size in Beetal and Teddy goats and least squares mean was used for multiple comparisons with litter size among genotypes.
where: Yük is the phenotypic value of litter size
μ is the population mean
B is the fixed effect of ith breed (Beetal and Teddy)
P is the fixed effect of jth parity
G is the fixed effect of kth SNP genotype
GB is the interaction of kth genotype and ith breed
eyk| is the random residual effect of each observation
Statistical analysis was performed with the GLM procedure of SAS program (version 8.2) (SAS Institute Inc., Cary, NC, USA). In the current study, the data were taken of only those births occurring in late winter to early spring.
Results
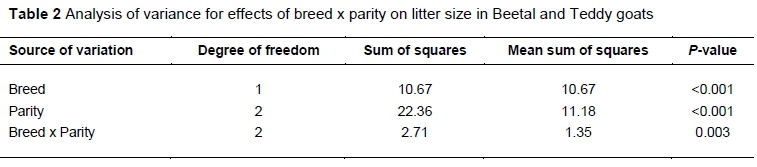
The results of the analysis of variance on litter size revealed a highly significant effect of breed (P <0.001) and parity (P <0.001) on litter size, separately. The average litter size for Beetal and Teddy was 1.708 ± 0.060 and 2.167 ± 0.056, respectively. Likewise, breed x parity showed significant differences (P <0.01 or P = 0.003) in the litter size in Parity 1, 3 and average parity in both goat breeds (Table 2). The mean litter size in Parity 1 and Parity 3 was 1.158 ± 0.06 1 and 2.062 ± 0.078 for Beetal and 1.861 ± 0.057 and 2.583 ± 0.073 for Teddy goat, respectively (Table 3).
The amplified fragments of BMPR1B (190 bp), GDF9 (500 bp) and BMP15 (141 bp) genes showed a size of 190 bp, 500 bp and 141 bp, respectively, indicating the primer specific amplification (Figure 1).
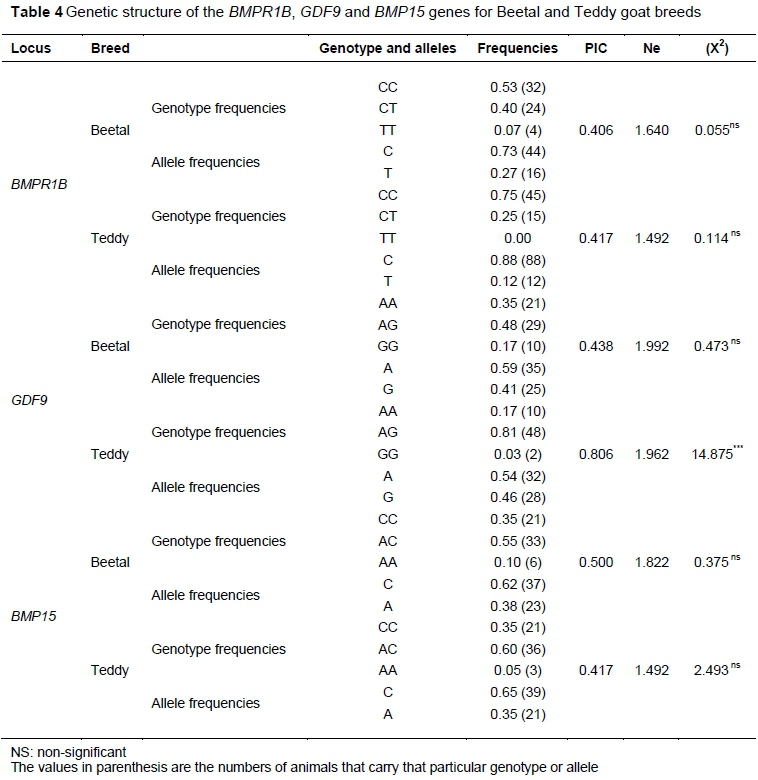
The PCR-RFLP genotyping of 120 goats, 60 of each of Beetal and Teddy breeds, showed CC, CT, and TT genotypes (Yacoub et al.). The CC genotype had both bands at 190 bp, while the TT genotype showed two fragments at 140 bp and 50 bp positions. However, the heterozygote CT showed fragments at 190 bp, 140 bp and 50 bp positions (Figure 2A). A pictorial view of changed allele from C to T is given in the chromatogram (Figures 2B and 2C). The frequencies of allele C and T in Beetal goats were 0.73 and 0.27, while in Teddy goats their frequencies were 0.88 and 0.12, respectively. Moreover, the frequencies of the three observed genotypes CC, CT and TT in Beetal goats were 0.53, 0.40 and 0.07, while in Teddy goats their values were 0.75, 0.25 and 0.00, respectively (Table 4).
For the BMPR1B gene, the CC, CT and TT genotypes had mean values of 1.60 ± 0.08, 1.84 ± 0.09 and 1.66 ± 0.24 litter size, respectively, in Beetal goats, showing the overdominance of the heterozygous genotype (CT). However, in Teddy goats CC and CT genotypes had mean values of 2.01 ± 0.07 and 2.37 ± 0.09 litter size, respectively (Table 5). The litter size of CT genotype was significantly (P <0.05) higher than that of other genotypes in Parity 1 in Beetal and Parity 2 and Parity 3, and average litter size in Teddy goats. The results of the current study showed the overdominant effect of heterozygous genotype in both breeds. However, no TT genotype was observed in Teddy goats in the present study.
The result of agarose gel electrophoresis showed three fragments of the lengths of 500bp, 300bp and 200bp (Figure 3A). Polymorphism was found in this gene in tested animals of both breeds and the three genotypes AA, AG and GG were observed. The AA genotype had both bands at 500 bp, while the GG genotype showed two fragments at 300bp and 200bp position. However, the heterozygote AG showed three fragments at 500bp, 300bp and 200bp positions (Figure 3A). Sequencing results (Figure 3C) show the substitution of nucleotide A to G at 300bp. Genotypic frequencies of GDF9 polymorphism in Beetal goat were 0.35, 0.48 and 0.17 for AA, AG and GG, respectively, whereas the frequencies of A and G alleles was 0.59 and 0.41 (Table 4). In Teddy goats, genotypic frequencies for AA, AG and GG were 0.21, 0.65 and 0.14, respectively, and allelic frequencies were 0.56 and 0.44 for A and G alleles, respectively (Table 4).
For GDF9, in both Beetal and Teddy goats, AG genotype was responsible for the highest litter size in all the parities, whereas the AA genotype was associated with the lowest litter size in all the parities in both breeds. In the Teddy goat, the AG genotype was associated with 1.90 ± 0.07, 2.17 ± 0.10, 2.72 ± 0.08 and 2.26 ± 0.06 litter size in first, second, third parity and average litter size of all three parities, respectively (Table 5), which was significantly (P <0.05) greater than values of litter size for other genotypes. However, in Beetal goats, the AG genotype had significantly greater litter size in second (2.07 ± 0.14), third (2.29 ± 0.12) and average parity (1.85 ± 0.09) than the other genotypes.
The PCR-RFLP genotyping of BMP15 gene in tested animals of both breeds showed AA, AC and CC genotypes (Figure 4A). In AA genotype, both bands were at 141bp, while CC genotype showed two fragments of 111bp and 40bp length, whereas the heterozygote genotype (AC) showed three fragments at 141bp, 111bp and 40bp (Figure 3A). Nucleotide change from A to C is shown in the chromatogram in Figure 4 (B and C). The frequencies of allele C and A in Beetal goats were 0.62 and 0.38, respectively, while in Teddy goats their frequencies were 0.68 and 0.32. Moreover, the frequency of the three genotypes, CC, AC and AA, in Beetal goats were 0.35, 0.55 and 0.10, while in Teddy goats their values were 0.35, 0.65 and 0.00, respectively (Table 4).

Least square means and standard errors (mean ± SE) of the three genotypes CC, AC and AA of BMP15 gene with litter size in Beetal and Teddy goat are presented in Table 5. In both Beetal and Teddy goats the CC genotype had the highest values of litter size in all the parities. In Teddy goats, the CC genotype was associated with significantly larger litter size in second, third, and average parities, which was 2.43 ± 0.09, 3.00 ± 0.05 and 2.44 ± 0.05, respectively. However, AA genotype was not observed in Teddy goats.
In Beetal goats, CC genotypes had the significantly (P <0.05) higher values of litter size (1.39 ± 0.09) in first parity compared with the CA and AA genotypes, whereas in second, third and average parity (litter size) it had significantly (P <0.05) greater value of litter size than that of AA genotype.
Discussion
The results of this study showed highly significant effects of breed (P <0.001), parity (P <0.001) and breed x parity interaction (P <0.01) on the litter size. The possible reason for the significant difference between breeds is attributable to breed difference (Hyder et ai., 2002), as the Teddy is highly prolific whereas the Beetal is considered medium prolific. Secondly, the parity is known to have positive effects on litter size in goats (Pan et ai., 2015). Consistent with the current results, Ahlawat et ai. (2015a) reported significant effects of breed and parity on litter size in six Indian goat breeds.
In genotypes, the mutations in BMPR1B, GDF9 and BMP15 genes were tested, all of which proved polymorphic in both breeds. Moreover, the results of breed x parity x genotype interaction showed significant differences in litter size in both breeds in all three parities.
As expected, the size of PCR product of BMPR1B gene was 190 bp, showing the primer specific amplification. After digestion with restriction endonuclease AvaII, both bands of the wild-type genotype (CC) were at 190 bp while the heterozygotes (CT) showed band at 190 bp, 140 bp and 50 bp and the mutant genotype (TT) showed both bands at 140 bp and 50 bp. In the current study, the results showed different patterns of bands, suggesting the presence of a mutation at the locus of BMPR1B (Figure 2A) in both breeds. The results showed that the frequency of the wild-type allele (C) was higher in animals of both breeds compared with the mutant allele (T). The goodness-of-fit of the results showed no significant difference in genotypic frequencies in either breed (Table 4). The BMPRIB gene is one of the major candidate genes that controls the ovulation rate and consequently increases the litter size in goats and sheep (Polley et ai., 2009a; Polley et ai., 2009b). The current results showed that the heterozygous carrier animals, which have the CT genotype, of both breeds had the highest litter size in all three parities, which might be because of overdominance of heterozygous individuals. The average litter size in Beetal and Teddy goats for the CT genotype was 1.84 ± 0.09 and 2.37 ± 0.09, respectively. The greater litter size in Teddy goats is probably because of breed effect, because Teddy goats are highly prolific, with greater incidence of triplets and quadruplets (Hyder et ai., 2002). In the current study, no animal of the TT genotype was observed in the tested animals of Teddy goats. However, it is likely that the mutant genotype (TT) can be observed in Teddy goats by increasing the sample size, because the CT genotype was found in this breed, and had significantly larger litter size compared with other genotypes. In agreement with the current results Polley et al. (2009a) observed the single nucleotide polymorphism in BMPR1B gene in Black Bengal goats and reported that the heterozygous carrier and homozygous carrier animals had 3.04 and 3.11 kids, respectively, compared with wild-type genotype, which had 2.7 kids/doe. However, in contrast with the current findings, no polymorphism in BMPR1B gene was found to be associated with litter size in six Indian goat breeds (Ahlawat et al., 2015a). The probable reason is that these authors focused on novel variations, which were not tested earlier. In another similar study, Ahlawat et al. (2014) found some single nucleotide polymorphisms in the promoter region of BMPR1B gene in nine Indian goat breeds, but it was not found to be associated with litter size. However, polymorphisms in BMPR1B gene have been detected in various sheep breeds (Ahlawat et al., 2016; Polley et al., 2009a). Chu et al. (2007) observed FecB mutation in Small-tailed Han sheep, and reported that heterozygous (FecB/FecB+) and homozygous mutant (FecB/FecB) genotypes had more lambs per ewe compared with ewes that had wild-type genotype. Likewise, Mahdavi et al. (2014) identified FecB mutation in Kalekoohi sheep, and observed that the (mutant) BB, and B+ genotypes had 0.52 and 0.35 more lambs per ewe compared with the wild-type (++) genotype.
The genotyping results of exon 1 of GDF9 gene in the present study showed that the investigated locus was polymorphic in both Beetal and Teddy goats. The three genotypes, AA, AG and GG, were observed in both breeds, which were found to be significantly (P <0.05) associated with litter size. In both breeds, A was the major allele and G the minor. However, the results of goodness-of-fit showed that there were significant (P <0.001) differences in genotypic frequencies in Teddy goats, whereas in Beetal goats there were none. In both Beetal and Teddy goats, the animals with AG genotype had the highest litter size in all the parities, whereas the AA (wild-type) genotype was associated with the lowest litter size in all the parities in both breeds. In Teddy goats, the AG genotype was associated with 1.90 ± 0.07, 2.17 ± 0.10, 2.72 ± 0.08 and 2.26 ± 0.06 litter size in first, second, third parity and average litter size, respectively, which was significantly (P <0.05) greater than values of litter size for other genotypes. However, in Beetal goats, the AG genotype had significantly greater litter size in second (2.07 ± 0.14), third (2.29 ± 0.12) and average parity (1.85 ± 0.09) than the other genotypes.
Consistent with the current results, Feng et al. (2011) observed three genotypes, AA, AC and CC, in Jining Grey goat and reported that CC and AC genotypes had maximum litter size compared with the wildtype genotype (AA). Likewise, Silva et al. (2011) observed a mutation (FecGE) in GDF9 gene in Santa Inês sheep, which caused a change of amino acid phenylalanine to cysteine in the conserved region. The authors reported that the ewes that were homozygous for FecGE had 82% higher rate of ovulation and 58% greater prolificacy. In the contrary, Ahlawat et al. (2016) studied polymorphism in GDF9 gene associated with litter size in seven goat breeds and found non-significant effect of genotype on litter size. Likewise, Dutta et al. (2013) investigated polymorphism in GDF9 through PCR-RFLP technique in Assam Hill goats and observed that all tested animals were monomorphic. Therefore, it was reported that litter size in this goat breed was not related to a mutated allele of GDF9. In a similar study on Black Bengal goats Polley et al. (2009a) investigated PCR-RFLP based polymorphism in the GDF9 gene and found that all the tested animals were monomorphic for these mutations. However, Zhu et al. (2013) observed a substitution mutation (A792-G) in exon 2 of GDF9 gene in Big Foot and Jintang Black goats, which was involved in change of amino acid valine to isoleucine. The AA genotype was significantly associated with greater litter size in these two Chinese goat breeds.
In this study three genotypes, AA, AC and CC, were observed in exon 2 of BMP15 gene, which showed the polymorphic nature of the investigated locus in both breeds, with A as major allele and C as minor. However, the results of goodness-of-fit did not show any significant differences between genotypic frequencies in both breeds. The results of association analysis showed that allele C was involved in increasing the litter size in both Beetal and Teddy goats, and homozygous carrier animals (CC) had the highest number of kids followed by the heterozygous (AC) genotypes. In agreement with the current results, Abdel-Rahman et al. (2013) observed that homozygous carrier animals of Anglo-Nubian goat (BB) had a significantly greater litter size compared with those with heterozygous and wild-type genotypes. The current results were consistent with the findings of Wang et al. (2011), who observed that homozygous mutant (BB) genotype had higher numbers of kids (0.91 kids) than those of the AB and AA genotypes in Funiu white goats. Although the biology of mutation was not explored in the current study, it is likely that altered mutation increases the ovulation rate in both breeds, which increases the litter size. This hypothesis was supported by the findings of Pramod et al. (2013), who reported that antral follicles of Black Bengal goats expressed 2.78-fold more upregulation of BMP15 gene than those of Sirohi goats. It was also suggested that the greater number of antral follicles per ovary and higher expression of BMP15 in Black Bengal goats could be the underlying reason for greater prolificacy in this breed.
However, according to Monteagudo et al. (2009), rates of ovulation in BMP15 mutants were greater in the heterozygotes, whereas the homozygous mutants showed a primary ovarian failure, leading to complete sterility in Rasa Aragonesa sheep. This is in contrast with the current results, in which homozygous mutant animals showed higher numbers of kids in Beetal goats. However, some studies reported that no association of polymorphism in BMP15 gene was observed in several Indian goat breeds (Ahlawat et al., 2016; Polley et al., 2009a). The association of the polymorphism in exon 2 of BMP15 with litter size of Beetal and Teddy goats suggests that it could be used in marker-assisted selection to enhance prolificacy in these two breeds.
Conclusion
These results indicate that the mutations in BMPRIB, GDF9 and BMP15 genes were significantly associated (P <0.05) with litter size in Beetal and Teddy goats. In BMPR1B and GDF9, the heterozygous individuals showed the highest litter size compared with other genotypes. However, in BMP15, the homozygous mutant genotype showed the largest litter size in both breeds. It is likely that tested mutations in all three genes could be used as molecular markers to select the most prolific animals of Beetal and Teddy goats. Further research on a large number of animals is required to confirm the link of these genes with increased prolificacy in both goat breeds.
Authors' Contributions
AB and IZ designed the study and did all the analysis. MI, AB and IZ selected the animals and performed sampling. MI did do all the lab work and assisted in analysis. MI, AB, and IZ prepared the final draft of this manuscript. All authors read and approved the draft.
Conflict of Interest Declaration
The authors declare that they have no conflicts of interest.
References
Abdel-Rahman, S.M., Mustafa, Y.A., Abd Errasool, H.A., El-Hanafy, A.A. & Elmaghraby, A.M., 2013. Polymorphism in BMP-15 gene and its association with litter size in Anglo-Nubian goat. Biotech. Anim. Husb. 29, 675-683. [ Links ]
Adhikari, K., AlChawa, T., Ludwig, K., Mangold, E., Laird, N. & Lange, C., 2012. Is It rare or common?. Genetic. Epidemiol. 36, 419-429. [ Links ]
Ahlawat S., Sharma R., Maitra, A., Tantia, M.S., Roy, M. & Mandakmale, S., 2014. New genetic polymorphisms in Indian goat BMPR1B gene. Indian J. Anim. Sci. 84, 37-42. [ Links ]
Ahlawat, S., Sharma, R., Maitra, A., Raja, K.N., Verma, N.K. & Tantia, M.S., 2015a. Prolificacy in Indian goat breeds is independent of FecB mutation. Indian J. Anim. Sci. 85, 617-620. [ Links ]
Ahlawat, S., Sharma, R., Maitra, A. & Tantia, M.S., 2015b. Current status of molecular genetics research of goat fecundity. Small Rumin. Res. 125, 34-42. [ Links ]
Ahlawat, S., Sharma, R., Roy, M., Mandakmale, S., Prakash, V. & Tantia, M., 2016. Genotyping of novel SNPs in BMPR1B, BMP15, and GDF9 genes for association with prolificacy in seven Indian goat breeds. Anim. Biotech. 27, 199-207. [ Links ]
Bodin, L., Pasquale, E.D., Fabre, S.P., Bontoux, M., Monget, P., Persani, L. & Mulsant, P., 2007. A novel mutation in the bone morphogenetic protein 15 gene causing defective protein secretion is associated with both increased ovulation rate and sterility in Lacaune sheep. Endocrinology 148, 393-400. [ Links ]
Bradford, G.E., Burfening, P.J. & Cartwright, T.C., 1989. Evaluation of production and reproduction of sheep, goat and alpaca genotypes in the Small Ruminant Collaborative Research Support Program. J. Anim. Sci. 67, 3058-3067. [ Links ]
Chu, M.X., Liu, Z.H., Jiao, C.L., He, Q.Y., Fang, L., Ye, S.C. & Wang, J.Y., 2007. Mutations in BMPR-IB and BMP15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 85, 598-603. [ Links ]
Chu, M.X., Wu, Z.H., Feng, T., Cao, G.L., Fang, L., Huang, W. & Li, N., 2011a. Polymorphism of GDF 9 gene and its association with litter size in goats. Vet. Res. Commun. 35, 329-336. [ Links ]
Chu, M.X., Yang, J., Feng, T., Cao, G.L., Fang, L., Di, R. & Li, N., 2011b. GDF9 as a candidate gene for prolificacy of Small Tail Han sheep. Mol. Bio. Rep. 38, 5199-5204. [ Links ]
Davis, G.H., 2005. Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. 37, 11-23. [ Links ]
Davis, G.H., Galloway, S.M., Ross, I.K., Gregan, S.M., Ward, J., Nimbkar, B.V. & Wilson, T., 2002. DNA tests in prolific sheep from eight countries provide new evidence on origin of the Booroola (FecB) mutation. Biol. Reprod. 66, 1869-1874. [ Links ]
Demars, J., Fabre, S., Sarry, J., Rossetti, R., Gilbert, H., Persani, L. & Bodin, L., 2013. Genome-wide association studies identify two Novel BMP15 mutations responsible for an atypical hyperprolificacy phenotype in sheep. PLoS Genet. 9, e1003482. [ Links ]
Dutta, R., Das, B., Laskar, S., Kalita, D.J., Borah, P., Zaman, G. & Saikia, D.P., 2013. Polymorphism, sequencing and phylogenetic characterization of growth differentiation factor 9 (GDF9) gene in Assam Hill goat. Afr. J. Biotechnol. 12, 6894-6900. [ Links ]
FAOSTAT, 2016. http://www.fao.org/faostat/en/?#data/QA. [ Links ]
Feng, T., Geng, C.X., Lang, X.Z., Chu, M.X., Cao, G.L., Di, R. & Li, N., 2011. Polymorphisms of caprine GDF9 gene and their association with litter size in Jining Grey goats. Mol. Biol. Rep. 39, 4029-4038. [ Links ]
Fogarty, N., 2009. A review of the effects of the Booroola gene (FecB) on sheep production. Small Rumin. Res. 85, 75-84. [ Links ]
Hanrahan, P.J., Gregan, S.M., Mulsant, P., Mullen, M., Davis, G.H., Powell, R. & Galloway, S.M., 2004. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol Reprod. 70, 900-909. [ Links ]
Helal, M., Mahboub, H., Hemeda, S., Ballal, S. & Heikal, H., 2014. Polymorphism of bone morphogenetic protein receptor-IB (BMPRIB) gene with litter size and kids growth of some goat breeds in Egypt. Alexandria J. Vet. Sci. 1, 28. [ Links ]
Hua, G.H., Chen, S.L., Yu, J.N., Cai, K.L., Wu, C.J., Li, Q.L. & Yang, L.G., 2009. Polymorphism of the growth hormone gene and its association with growth traits in Boer goat bucks. Meat Sci. 81, 391-395. [ Links ]
Hyder, A.U., Akhtar, P. & Gondal, K.Z., 2002. Influence of some non-genetic factors on birth weight of Teddy goat kids. Pakistan Vet. J. 22, 116-119. [ Links ]
Kasiriyan, M.M., Hafezeyan, H., Sayahzadeh, H., Jamshidi, R., Asghari, S.R., Irajeyan, G.H. & Buesagh, H., 2009. Genetic polymorphism fecb and bmp15 genes and its association with litter size in sangsari sheep breed of Iran. J. Anim. Vet. Adv. 8, 1025-1031. [ Links ]
Khan, M., Khan, M. & Mahmood, S., 2008. Genetic resources and diversity in Pakistani goats. Int. J. Agri. Biol. 10, 227-231. [ Links ]
Khan, M.S., Muhammad, M.S. & Abdullah, M., 2013. Livestock Keepers' perception of indigenous goat breeds and their contribution to livelihoods in Pakistan. Egyp. J. Sheep Goat Sci. 8, 17-28. [ Links ]
Khan, S.A., Kalhoro, B.I., Gandahi, J.A., Tunio, A.N., Vistro, W.A., Ahmed, M. & Fareed, S.K., 2016. Gross anatomical studies on testis and epididymis at pre-pubertal and pubertal stages of Teddy goat. Acad. Res. Internat. 7, 51-57. [ Links ]
Kumar, V., Kumari, N., Singh, L.B. & Kumar, S., 2016. Effect of FecB gene on body weight in Black Bengal goat. J. Anim. Res. 6, 247. [ Links ]
Kumari, R.D.S., Kumar, S., Lal, S.V., Chakraborti, A., Barari, S.K. & Amitava, D., 2016. Genetic polymorphism of bone morphogenetic protein receptor type 1 gene in Black Bengal goat and its association with litter size. Indian J. Anim. Sci. 85, 469-471. [ Links ]
Mahdavi, M., Nanekarani, S. & Hosseini, S.D., 2014. Mutation in BMPR-IB gene is associated with litter size in Iranian Kalehkoohi sheep. Anim. Reprod. Sci. 147, 93-98. [ Links ]
Monteagudo, L.V., Ponz, R., Tejedor, T.M., Lavina, A. & Sierra, I., 2009. A 17 bp deletion in the bone morphogenetic protein 15 (BMP15) gene is associated to increased prolificacy in the Rasa Aragonesa sheep breed. Anim. Reprod. Sci. 110, 139-146. [ Links ]
Montgomery, G.W., Crawford, A.M., Penty, J.M., Dodds, K.G., Ede, A.J., Henry, H.M. & Hill, D.F., 1993. The ovine Booroola fecundity gene (FecB) is linked to markers from a region of human chromosome 4q. Nat. Genet. 4, 410. [ Links ]
Muhammad, M.S., Abdullah, M., Khan, M.S., Javed, K. & Jabbar, M.A., 2015. Farmers preferences for goat breeds in Punjab. Pakistan. J. Anim. Plant Sci. 25, 380-386. [ Links ]
Mullen, M.P.H.J., Howard, D.J. & Powell, R., 2013. Investigation of prolific sheep from UK and Ireland for evidence on origin of the mutations in BMP15 (FecXG, FecXB) and GDF9 (FecGH) in Belclare and Cambridge sheep. PLoS ONE. 8. [ Links ]
Nawaz, A., Babar, M.E., Hussain, T., Nadeem, A., Bilal, F., Javed, K. & Muhammad, K., 2013. Identification of molecular markers in BMP15 gene of Pakistani goat breeds. Int. J. Adv. Res. 1, 62-68. [ Links ]
Noshahr, F.A. & Rafat, A., 2014. Genetic polymorphism of GDF9 gene in Iranian Moghani sheep breed. Iran. J. Appl. Anim. Sci. 4, 887-890. [ Links ]
Pan, S., Biswas, C.K., Majumdar, D., Sengupta, D., Patra, A., Ghosh, S. & Haldar, A., 2015. Influence of age, body weight, parity and morphometric traits on litter size in prolific Black Bengal goats. J. Appl. Anim. Res. 43, 104-111. [ Links ]
Polley, S., De, S., Batabyal, S., Kaushik, R., Yadava, P., Aroraa, J.S. & Oswami, S.L., 2009a. Polymorphism of fecundity genes (BMPR1B , BMP15 and GDF9) in the Indian prolific Black Bengal goat. Small Rumin. Res. 85, 122-129. [ Links ]
Polley, S., De, S., Brahma, B., Mukherjee, A.P.V.V., Batabyal, S. & Goswami, S.L., 2009b. Polymorphism of BMPR1B, BMP15 and GDF9 fecundity genes in prolific Garole sheep. Trop. Anim. Health Prod. 42, 985-993. [ Links ]
Pramod, R.K., Sharma, S.K., Singhi, A., Pan, S. & Mitra, A., 2013. Differential ovarian morphometry and follicular expression of BMP15, GDF9 and BMPR1B influence the prolificacy in goat. Reprod. Domes. Anim. 48, 803-809. [ Links ]
Rojas-Downing, M.M., Nejadhashemi, A.P., Harrigan, T. & Woznicki, S.A., 2017. Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk Manag. 16, 145-163. [ Links ]
Rosatia, A., Mousa, E., Van Vleck, L.D. & Young, L.D., 2002. Genetic parameters of reproductive traits in sheep. Small Rumin. Res. 43, 65-74. [ Links ]
Shabir, M., Ganai, T.A.S., Misra, S.S., Shah, R. & T, A., 2013. Polymorphism study of growth differentiation factor 9B (GDF9B) gene and its association with reproductive traits in sheep. Gene 512, 432-443. [ Links ]
Shi, T., Mazumdar, T., DeVecchio, J., Duan, Z.H., Agyeman, A., Aziz, M. & Houghton, J.A., 2010. cDNA microarray gene expression profiling of Hedgehog Signaling Pathway Inhibition in human colon cancer cells. PLoS ONE. 5. [ Links ]
Shokrollahi, B., 2014. Investigation of BMP15 gene polymorphisms associated with twining in Markhoz goat. Biharean Biol. 9, 1-4. [ Links ]
Silva, B.D.M., Castro, E.A., Souza, C.J.H., Paiva, S.R., Sartori, R., Franco, M.M. & Melo, E.O., 2011. A new polymorphism in the Growth and Differentiation Factor 9 (GDF9) gene is associated with increased ovulation rate and prolificacy in homozygous sheep. Anim. Genet. 42, 89-92. [ Links ]
Sudhir, K., Glen, S. & Koichiro, T., 2016. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets Mol. Bio. Evo. 33, 1870-1874. [ Links ]
Thompson, J.D., Higgins, G. & Gibson, T.J., 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acid Res. 22, 4673-4680. [ Links ]
Wang, Y., Yuanxiao, L., Nana, Z., Zhanbin, W. & Junyan, B., 2011. Polymorphism of Exon 2 of BMP15 gene and its relationship with litter size of two Chinese goats. Asian-Austral. J. Anim. Sci. 24, 905-911. [ Links ]
Wilson, T., Wu, X-Y., Juengel, J.L., Ross, I.K., Lumsden, J.M., Lord, E.A. & Montgomery, G.W., 2001. Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of Bone Morphogenetic Protein IB Receptor (ALK-6) that is expressed in both oocytes and granulosa cells. Biol. Reprod. 64, 1225-1235. [ Links ]
Yacoub, H.A., Fathi, M.M. & Mahmoud, W.M., 2013. DNA barcode through cytochrome B gene information of mtDNA in native chicken strains. Mitoch. DNA. 24, 528-537. [ Links ]
Zhu, G.Q., Wang, Q.I., Kang, Y.G., Lv, Y.Z. & Cao, B.Y., 2013. Polymorphisms in GDF9 gene and its relationship with litter size in five breeds of Black Goats. Iran. J. Appl. Anim. Sci. 3, 625-628. [ Links ]
Received 13 August 2018
Accepted 5 June 2019
First published online 30 July 2019
# Corresponding author: atia.basheer@uvas.edu.pk


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}