










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 n.4 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i4.6
ARTICLES
Effect of additives on fermentation characteristics, nutrient values and in vitro fermentation profile of Neolamarckia cadamba leaf silage
L. HeI; C. WangII; W. ZhouI; Q. ZhangI; X. ChenI, #
IGuangdong Key Laboratory for Innovative Development and Utilization of Forest Plant Germplasm, Guangdong Province Research Centre of Woody Forage Engineering Technology, College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou, China
IICollege of Animal Science, South China Agricultural University, Guangzhou, China
ABSTRACT
This study was designed to investigate the effects of additives on the ensiling characteristics of Neolamarckia cadamba leaf (NCL) silage. The NCL were allocated to six ensiling treatments, namely control (C-S), uncompacted (UC-S) (negative control), with the addition of sodium sulphite (S-S), with propionate (P-s), with citric acid plus sodium acetate (CA-S), and with glucose (G-S). Fermentation parameters, chemical compositions and in vitro fermentation profile of the silages were investigated after 60 days' storage. Compared with the control, the application of additives improved dry matter recovery (DMR), while UC-S had a poorer DMR. All the silages increased in pH value and acetate content, except for P-S. Lactate concentration was higher in S-S and G-S, and lower in UC-S, P-S, and CA-S. P-S, CA-S, and G-S had increased water soluble carbohydrate (WSC) concentrations. Soluble crude protein was higher in UC-S and CA-S. All the treated silage had greater gas production at slower rates, except for S-S. The fermentation of S-S, P-S and CA-S resulted in higher content of short-chain fatty acids and microbial crude protein relative to UC-S. S-S (38.82%) and CA-S (37.89%) had higher organic matter digestibility relative to the control (36.43%). S-S (42.80%) and P-S (43.99%) had higher crude protein digestibility, which was lower in UC-S (30.73%) when compared with the control (37.74%). There was no difference in the predicted available energy of these silages. The low ammonium nitrogen ratio and the high proportion of true protein inferred that protein was well preserved in these silages, even the uncompacted silage. These results suggest that sulphite and organic acids, such as propionate and citrate, could be an option to improve the feed quality of NCL silage, and its great protein conservation deserves more attention.
Keywords: feed quality, in vitro technique, Neolamarckia cadamba, ruminant nutrition, silage processing
Introduction
Neolamarckia cadamba, an evergreen tropical tree that is native to South and South East Asia, has been exploited as woody forage in the past few years, because of its rapid growth, biomass yield and high protein content, and abundant components such as plant polyphenols (He et al., 2018). A previous study showed that harvesting N. cadamba at intervals of two months for silage processing would obtain the largest biomass yield over the year (Zhang, 2018). Moreover, N. cadamba could be high-quality forage that is comparable with the traditional Leucaena leucocephala and Medicago sativa for ruminant animals (Zayed et al., 2014). Partially replacing corn silage with N. cadamba silage in a fattening diet could improve the growth performance and meat quality of Lezhi black goats (Wang et al., 2017). Making full use of this unconventional feed resource could be an important measure to confront the challenge of feed supply.
Ensiling seems to be the most practical way of keeping forage supplied all year round. It is based on lactic acid bacteria (LAB), which convert WSC anaerobically into organic acids to decrease the pH and inhibit the activity of spoilage organisms (McDonald et al., 1991). However, the water soluble carbohydrate (WSC) content of N. cadamba leaf (NCL) is typically lower than 5% (He et al., 2018; Wang et al., 2018), which is thought to be insufficient to support the pH decrease (Ni et al., 2017). Generally, there are several potential strategies to improve the ensiling process, that is by increasing the LAB number by inoculating, lowering pH by acid addition, clearing oxygen residue by antioxidant application, and supplying a fermentable substrate such as sugar (Kung et al., 2003). An LAB inoculant that is prepared by incubating the extract anaerobically is usually applied to improve silage quality because of a lack of epiphytic LAB, but the inoculant seems to show inconsistent effects on different materials (Ellis et al., 2016). However, the authors' previous research showed that the LAB number of inoculated NCL silage was even lower than the control silage, and improved effects were rarely found in the silage fermentation (He et al., 2018). Therefore, to exploit suitable additives for NCL silage, several additives were selected in this study rather than LAB inoculant.
It was commonly supposed that chemical additives would improve the ensiling process of NCL silage, but few studies have been reported in the literature. Therefore, this study was conducted to investigate the effects of silage additives, namely sodium sulphite, propionic acid, citrate plus sodium acetate and glucose, on the fermentation characteristics, carbohydrate constituents, protein fractions, in vitro gas production and digestibility of NCL silage.
Materials and Methods
The ensiling procedures of this study were in accordance with He et al. (2018). In brief, NCL were collected manually from Yuejin North Experimental Plot of South China Agricultural University, and then chopped into a particle length of 2 - 3 cm with a hand hay cutter. The prepared NCL was ensiled with the following treatments in six replicates: control silage (C-S), uncompacted silage (UC-S) (negative control), silage with the addition of sodium sulphite (S-S), silage with propionic acid (P-S), silage with citric acid plus sodium acetate (1:1, w/w) (CA-S) and silage with D-glucose (G-S). All the chemical agents (analytical reagents) were purchased from Guangzhou Chengshuo Reagent Company (Guangzhou, China), and their inclusion levels in the silages were 1% (w/w) of fresh matter. The additives were dissolved in 10 mL distilled water and sprayed with a mini sprayer onto 1.0 kg leaves, while the control was sprayed with 10 mL distilled water. All the pre-mixed forage was compacted in laboratory polyethylene bags (20 cm by 30 cm) and sealed with a food vacuum sealing machine, except that the UC-S was sealed without extraction of air. Following 60 days' fermentation at room temperature (around 28 °C), each bag was unsealed and sampled twice, so that one part was used to determine silage fermentation, and the other was oven-dried at 65 °C for 48 hours and ground in a hammer mill to pass a 1-mm sieve for chemical analysis and in vitro culture.
The extract fluid was prepared by stirring fresh silage (25 g) in distilled water (225 mL) with a domestic fruit juicer for three minutes, and then filtered with four layers of cheesecloth to attain the supernatant. With the fluid, pH value was measured using a pH meter (PHS-3C, INESA INSTRUMENT, Shanghai, China). Organic acids, including lactic acid, acetic acid, propionic acid and butyric acid, were analysed by high performance liquid chromatography (HPLC), which was equipped and set as follows: Shodex RS Pak KC-811 column (Showa Denko K.K., Kawasaki, Japan), SPD-20A detector (Shimadzu Co., Ltd, Kyoto, Japan); eluent 3 mmol/L HClO4 with a running rate of 1.0 mL/min, temperature of column oven 50 °C. Ammonium-nitrogen (NH3-N) was measured according to the method of Broderick & Kang (1980). Dry matter recovery (DMR) was measured as the difference in dry matter (DM) content before and after 60 days' silage fermentation.
Samples were analysed in duplicate for DM, crude protein (CP), neutral detergent fibre (NDF), acid detergent fibre (ADF), acid detergent lignin (ADL) and ash according to AOAC (2000) procedures. Specifically, CP was analysed by an automatic Kjeldahl apparatus (K-9860, Hanon, Jinan, China). NDF, ADF and ADL were analysed using an A220 fibre analyser (ANKOM Technology Corp., Macedon, NY, USA) without sodium sulphite and heat stable amylase, and the results were expressed inclusive of ash. Ash was measured by combustion in a muffle furnace at 550 °C for four hours following calcination. Protein fractions, including non-protein nitrogen (NPN), true protein (TP) and soluble crude protein (SCP), were analysed according to the method of Licitra et al. (1996). WSC was measured with anthrone-sulphuric colorimetry according to the instructions of the test kit (Comin Biotechnology Ltd, Suzhou, China). Total phenols and hydrolysable tannins were determined with Folin-Ciocalteu colorimetry (Min et al. (2015). Condensed tannins were measured according to Coblentz & Grabber (2013).
In vitro incubation was carried out with an ANKOM RFS gas production meter (ANKOM Technology Corp., Macedon, NY, USA) according to the instrument's specifications. Before the morning feeding, rumen fluid was collected from three Holstein cows (approximately 500 kg bodyweight) that were fitted with permanent rumen cannulae and fed twice a day a total mixed ration consisted of 50% corn silage and 50% commercial concentrate, and then strained through four layers of cheesecloth into a vacuum bottle and transported to the laboratory within 20 min. The rumen fluid was mixed with the buffer solution in a 1:2 (v/v) proportion under a continuous flow of CO2. The buffer solution and incubation fluid were prepared according to Menke & Steingass (1988). Each sample was weighed (1.0 g) in triplicate into 250 mL glass bottles in advance. All the bottles (with three blank controls), which were injected with 150 mL incubation fluid, were incubated at 39 °C for 48 hours, and gas production (GP) (mL) was recorded automatically at 0.5-hour intervals. At the end, the fermentation fluid was measured for pH value (PHS-3C, INESA Scientific Instrument Co, Shanghai, China) and then centrifuged to obtain the supernatant that was designated for NH3-N analysis. The residue was collected and dried at 65 °C for 48 hours to calculate in vitro dry matter digestibility (DMD). The residue of the same sample was mixed to determine the digestibility of NDF and ADF (NDFD and ADFD).
Enzyme degradation of crude protein (CPD) was conducted with cellulase (400 U/mg, Trichoderma viride) and pepsin (3000 U/g, porcine mucosa) that were purchased from Beizhuo Biotech Company (Shanghai, China), according to the method of Faithfull (2002). First, Erlenmeyer flasks (150 mL) containing 0.5 g feed sample and 40 mL cellulase citrate-phosphate buffer (6 g/L, pH = 4.6) were incubated in the shaker setting at 150 rpm/min and 39 °C for 24 hours. After that, the enzymes were inactivated in boiling water for 10 min. Subsequently, the flasks were filled with 10 mL pepsin-0.1 M HCl solution (35 g/L) and incubated for another 48 hours. Finally, the residue was collected by filtering and analysed for nitrogen content.
To estimate the kinetic parameters of gas production, gas production records were fitted using the nonlinear option of SAS (version 9.0), with the model described by France etal. (2000):
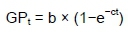
where: GPt (mL/g DM) is the volume of GP at time t
b (mL/g DM) is the asymptotic GP
c (/h) is the rate of gas production
The partitioning factor at 48 hours of incubation (PF48), a measure of fermentation efficiency, was calculated as the ratio of DMD (mg/g) to the volume (mL/g) of GP at 48 hours (i.e., DMD/GP48) according to Blümmel et al. (1997). Gas yield (GY24) was calculated as the volume of gas (mL/g DM) produced after 24 hours of incubation divided by the amount of DMD (g) (GP24 /DMD). Short-chain fatty acid (SCFA) concentration was calculated according to Getachew et al. (2002) as:
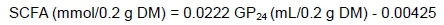
Microbial crude protein (MCP) biomass production was calculated according to Blümmel et al. (1997) as:
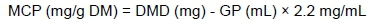
where: 2.2 mg/mL is a stoichiometric factor, which expresses mg of C, H and O required for the production of SCFA gas associated with the production of 1 mL of gas.
Organic matter digestibility (OMD) and metabolizable energy (ME) were calculated according to Menke & Steingass (1988), and the net energy for maintenance (NEm) and for gain (NEg) were calculated according to NRC (2000):

All data were subjected to the GLM procedure of SAS (version 9.0, SAS Institute Inc., Cary, NC, USA), considering additive treatments as fixed factors in the linear model:
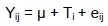
where: γ is every observation of silage treatment i (T,)
μ is the general mean; T is the effect of treatment i
ey is experimental random residual error
Duncan's test was used to do multiple comparisons with differences being declared significant at P <0.05.
Results and Discussion
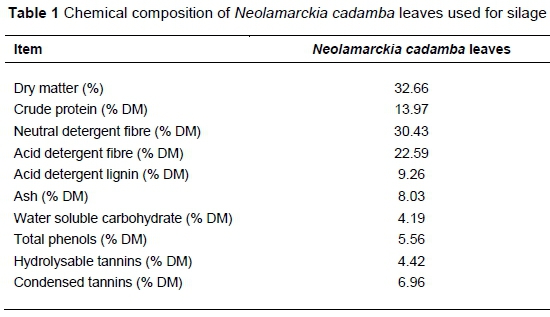
The chemical composition of raw NCL is presented in Table 1. The results showed that NCL contained about 14% CP and 30% NDF, in which the protein level was high enough for beef cattle and the fibre content fell into the dietary range (NRC, 2000), thus promoting its high inclusion in the diet. These data were in line with the results of Wang et al. (2018) and Zhang (2018). WSC is critical for silage fermentation, acting as the fermentation substrate, in which 5% is thought to be the threshold for acceptable fermentation (Ni et al., 2017), inferring that ensiling NCL (4.42% WSC) without the application of additives may not work well. Moreover, NCL contains high contents of phenols and tannins, which might affect palatability and feed utilization because of their protein binding and precipitating properties (Estell, 2010; Huang et al., 2016). It has also been reported that forage conserved as silage greatly diminished the extractability of protein-protecting polyphenols (Terrill et al., 1990; Minnee et al., 2002), most of which were transformed into compounds during ensiling (unpublished data), suggesting that NCL silage may be superior in palatability relative to its raw material.
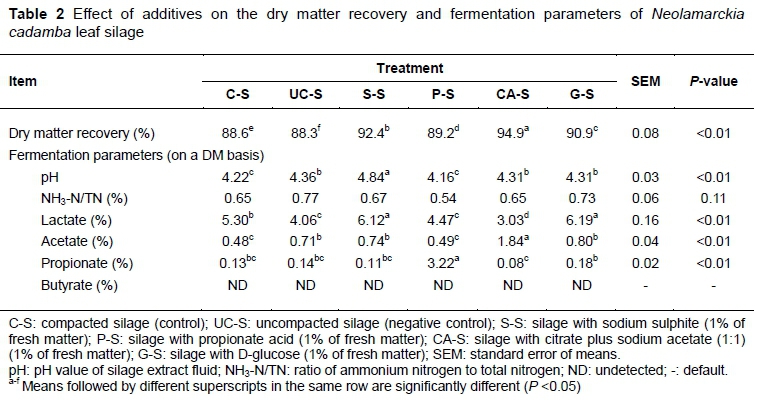
The data that reflect the fermentation quality of NCL ensiled with various additives (C-S, UC-S, S-S, PS, CA-S and G-S) are summarized in Table 2. There were significant (P <0.01) differences in DMR (88.31% -94.93%), pH value (4.16 - 4.84), and the contents of lactate (3.03% - 6.19%), acetate (0.48% - 1.84%), and propionate (0.08% - 3.22%) of NCL ensiled with various additives. Dry matter loss occurred inevitably during ensiling owing to the respiration of plant cell and aerobic fermentation in the initial phase (McDonald et al., 1991; Dos Santos et al., 2015), but it could be minimized. Any strategies that could inhibit these activities may benefit nutrient preservation. In the present study, additives improved (P <0.05) DMR, as expected, and the uncompacted treatment resulted in a poorer (P <0.05) DMR. The additives would alter the ensiling process in alternative pathways by promoting pH decrease, inhibiting microbial activity and accelerating the dominance establishment of LAB. The study of Zhao et al. (2018), which investigated dynamic variations in pH and DM loss of rice straw silage, showed that additives (hemicellulase and Lactobacillus plantarum) resulted in a rapid pH reduction at the early stage of ensiling and significantly decreased DM loss. pH is a simple parameter to evaluate the extent of silage fermentation and silage quality (Wang et al., 2009). A lower pH generally inferred further fermentation and better aerobic stability. Compared with the control, all the silages had higher (P <0.01) pH values and acetate contents, except for P-S, in which lactate concentration was higher (P <0.01) in S-S and G-S, and lower (P <0.01) in UC-S, P-S and CA-S. Not compacting resulted in decreased lactate content and thus increased the pH value (P <0.01) because more fermentable substrate was consumed in aerobic activity. In view of the changes of lactate content, the addition of sodium sulphite and glucose promoted the activity of acid-producing bacteria, while the addition of acids inhibited LAB fermentation. The chemical nature of the additives would also make a difference. The addition of sodium sulphite led to a higher pH value (4.84) because of its alkalinity, which might impair the aerobic stability of the silage. It is reported that moulds are usually inhibited at pH < 4.5 (Kung et al., 2003). Adding acetate and propionate led to a greatly increased concentration of the corresponding acids, but did not decrease the pH of the silages, which may be attributed partly to the differences in their acidity and buffering capacity. Similarly, Li et al. (2016) showed that citrate inhibited the production of lactic acid and acetic acid of alfalfa silage, resulting in a relatively higher final pH value. Additionally, all these silages had low NH3-N proportions, with butyric acid being undetected, indicating good preservation of protein. Given that the silage pH values did not seem low enough to achieve such high true protein recovery, which was not expected for high-protein silage, it is speculated that this is correlated with their high tannin content and other functional components. The naturally occurring condensed tannins were verified for reducing proteolysis of alfalfa during conservation (Grabber & Coblentz, 2009; Grabber et al., 2011).
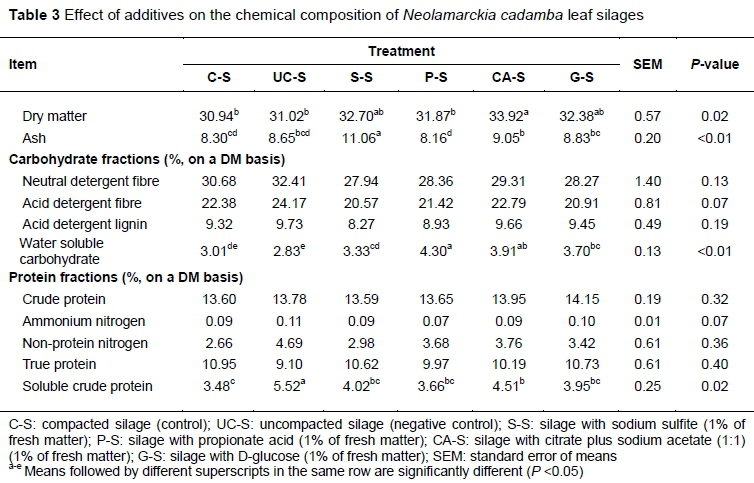
The nutrient values, including DM, ash, carbohydrate constituents and protein fractions, of various NCL silages are shown in Table 3. In the present study, uncompacted silage showed a similar chemical profile to the control, mainly because the residual air was not sufficient to make a difference, suggesting that there should be an optimum compacting density for practical silage production. CA-S increased (P <0.05) in the contents of DM and ash, and S-S presented a higher (P <0.01) ash content, which should be attributed to the addition of sodium sulphite and sodium acetate. P-S, CA-S and G-S had increased (P <0.01) WSC concentrations. that is, the application of propionate, citrate plus sodium acetate and glucose resulted in higher concentrations of WSC. WSC in the silage generally derived from the residual WSC of feedstuff and the outcome of polysaccharide hydrolysis. The higher WSC content in the silages with the addition of acid should be attributed to their fermentation inhibition and the acidolysis of cellulose and hemicellulose (McDonald et al., 1991; Zhang et al., 2016). Owing to the effect of plant protease and microbial activity during ensiling, part of the true protein is degraded to non-protein nitrogen (NPN), including mainly peptides, free amino acids, and NH3-N (McDonald et al., 1991). To evaluate the extent of protein conservation, there is a need to analyse their protein fractions, which would determine the bioavailability of CP (Licitra et al., 1996). Protein solubility is an important indicator for protein feed as it largely determines the ratio of rumen by-pass protein (Hvelplund & Madsen, 1990). In the present study, all the silages had relative low protein solubility in the borate-phosphate buffer, where UC-S and CA-S had increased (P <0.05) soluble crude protein, inferring that uncompacted silage suffered more severe proteolysis, given its high NPN concentration, and the addition of citrate caused more protein acidolysis. The low protein solubility may be because of the high tannin content (condensed tannin>7%), because tannin extracts are able to bind with proteins to form complexes and prevent protein hydrolysis in silage and in the rumen (Piluzza et al., 2014; Herremans et al., 2019). It was reported that naturally occurring condensed tannins contributed to the reduced proteolysis of alfalfa during conservation (Grabber & Coblentz, 2009). NPN often accounts for more than half of the total N in alfalfa silage and can result in inefficient N utilization by ruminants (Li et al., 2016). The present results inferred that the protein was well preserved in the silages in view of the low NH3-N concentration and the high ratio of true protein, which should be taken advantage of in practical production to modify dietary bypass protein, improving the efficiency of nitrogen utilization in ruminants and decreasing nitrogen excretion.
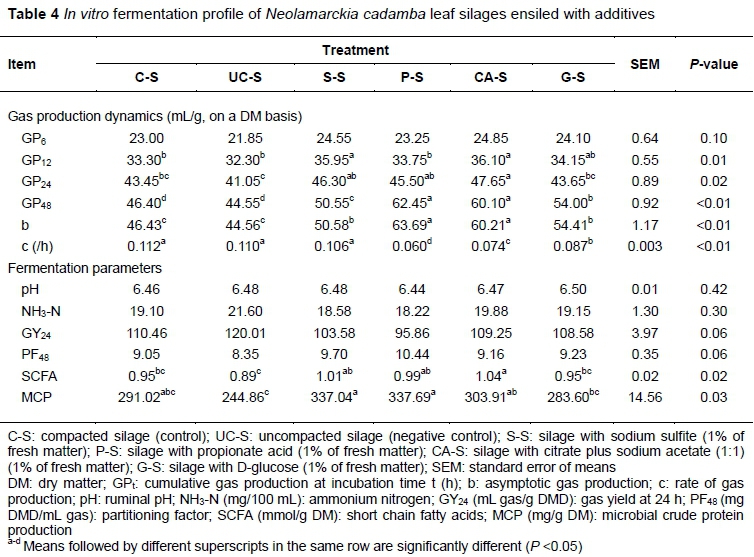
Because in vivo measurement is time consuming and varies widely, in vitro techniques such as in vitro rumen culture and enzymatic digestion have been developed as common methods to evaluate the nutritional value of ruminant feeds (Wang, 2016). As one of the most widespread techniques, in vitro gas production is a simple way of evaluating feed quality that reflects the extent of feed fermentation and digestibility (He et al., 2015). In the present study, the gas production curves of various NCL silages showed apparent gaps after 30 hours' incubation (Figure 1). Compared with the control, cumulative gas production was higher (P <0.01) in S-S and CA-S at 12 hours' fermentation and higher (P <0.05) in CA-S at 24 hours. All the silages had higher (P < 0.01) gas production (GP48 and b) with slower (P <0.01) rates of gas production (c), except for S-S (Table 4), suggesting that additive treatments resulted in more fermentable carbohydrates in the silages, which was in line with their increased WSC contents. Thus, SCFA, the primary fermentation product of carbohydrates, was expected to increase, probably contributing to the improved energy supply of the silage. Silage S-S, P-S, and CA-S resulted in higher (P <0.05) contents of SCFA and MCP relative to UC-S (Table 4), verifying that uncompacted silage had an inferior supply of energy and protein. It is inferred that poor energy supply would decrease the efficiency of microbial protein synthesis (Fijatkowska et al., 2015). No differences (P >0.05) were found in the other fermentation parameters such as pH, NH3-N, GY24 and PF48.
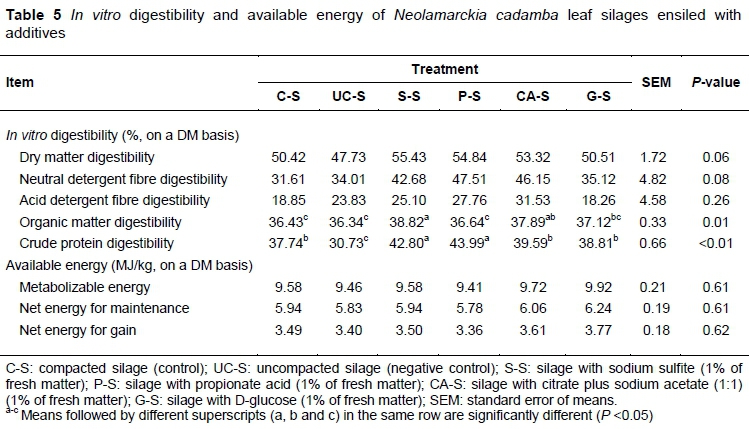
Digestibility is one of the most important factors that affect forage intake and depends largely on the chemical composition (Huhtanen et al., 2007; He et al., 2018). Measurement of in vitro DM digestibility has been used widely to assess the nutritional quality of feeds because of its high correlation with in vivo digestibility (He et al., 2015). Typically, in vitro gas production at 24 hours of ruminant feed is highly correlated with its digestibility and available energy content (He et al., 2015). In the present study, silage S-S and CA-S had higher (P <0.01) OMD, and CPD was higher in S-S and P-S, and lower in UC-S (P <0.01) (Table 5). Applying additives at ensiling tended to improve the digestibility of the silages, especially the cell wall components, coinciding with the comparison of their gas production. The additives may destroy the complex structure of the fibre matrix and then improve its accessibility. Moreover, the CPD of S-S and P-S increased significantly, where the changes of tannin-protein complex should be to blame, at least in part. Unfortunately tannin content had not been monitored during silage processing in the present study. Tannins could combine with protein or bind digestive enzyme, exerting an intricate effect on protein digestibility (Tabacco et al., 2006). The exact causes need further study, but these findings provide a possible method to use tannin-rich forages. The energy content of feedstuff is highly correlated with the digestibility of DM or OM (Wang, 2016). However, the differences in the predicted available energy (ME, NEm and NEg) of the silages were not large enough to be significant (P >0.05). The endpoint of in vitro incubation could also bias the comparison because different silages underwent various degradation rates. That is why one does not see consistent results with different evaluation methods.
Conclusions
The results of the present study showed that applying sodium sulphite or glucose to N. cadamba leaf silage at ensiling promoted lactate production, while the additions of propionate and citric acid plus sodium acetate decreased lactate concentration. The application of additives resulted in higher WSC content, and higher in vitro gas production with improved digestibility. The low NH3-N ratio and the high proportion of true protein inferred that the protein was well preserved in these silages, even the uncompacted silage. These findings suggest that sulphite and organic acids such as propionate and citrate acid could be an option to improve the feed quality of N. cadamba leaf silage, which is high in true protein proportion.
Acknowledgements
This work was financially supported by the Guangdong Young Innovative Talents Project (218220), and Guangzhou Science Technology & Innovation Commission (201707010462).
Authors' Contributions
LH and XC conceived and designed the experiments. LH, CW and WZ conducted the field trial, and collected and analysed the samples. LH wrote the paper. QZ and LH discussed and reviewed the paper.
Conflict of Interest Declaration
The authors have no conflict of interest to declare.
References
AOAC, 2000. Official Methods of Analysis. 17th edition. Association of Official Analytical Chemists, Gaithersburg, MD, USA. [ Links ]
Blümmel, M., Steingass, H. & Becker, K., 1997. The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed intake of roughages. Br. J. Nutr. 77, 911-921. [ Links ]
Broderick, G.A. & Kang, J.H., 1980. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 63, 64-75. [ Links ]
Coblentz, W.K. & Grabber, J.H., 2013. In situ protein degradation of alfalfa and birdsfoot trefoil hays and silages as influenced by condensed tannin concentration. J. Dairy Sci. 96, 3120-3137. [ Links ]
Dos Santos, W.C.C., Nascimento, W.G.D., Magalhães, A.L.R., Silva, D.K.A., Silva, W.J.C.S., Santana, A.V.S. & Soares, G.S.C., 2015. Nutritive value, total losses of dry matter and aerobic stability of the silage from three varieties of sugarcane treated with commercial microbial additives. Anim. Feed Sci. Technol. 204, 1-8. [ Links ]
Ellis, J.L., Bannink, A., Hindrichsen, I.K., Kinley, R.D., Pellikaan, W.F., Milora, N. & Dijkstra, J., 2016. The effect of lactic acid bacteria included as a probiotic or silage inoculant on in vitro rumen digestibility, total gas and methane production. Anim. Feed Sci. Technol. 211, 61-74. [ Links ]
Estell, R.E., 2010. Coping with shrub secondary metabolites by ruminants. Small Rumin. Res. 94, 1-9. [ Links ]
Faithfull, N., 2002. Methods in Agricultural Chemical Analysis: A practical handbook. CABI, Wallingford, USA. [ Links ]
Fijalkowska, M., Pysera, B., Lipinski, K. & Strusinska, D., 2015. Changes of nitrogen compounds during ensiling of high protein herbages-a review. Annals Anim. Sci. 15, 289-305. [ Links ]
France, J., Dijkstra, J.M., Lopez, S. & Bannink, A., 2000. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: Derivation of models and other mathematical considerations. Br. J. Nutr. 83, 131-139. [ Links ]
Getachew, G., Crovetto, G.M., Fondevila, M., Krishnamoorthy, U., Singh, B., Spanghero, M., Steingass, H., Robinson, P.H. & Kailas, M.M., 2002. Laboratory variation of 24 h in vitro gas production and estimated metabolizable energy values of ruminant feeds. Anim. Feed Sci. Technol. 102, 169-180. [ Links ]
Grabber, J.H. & Coblentz, W.K., 2009. Polyphenol, conditioning, and conservation effects on protein fractions and degradability in forage legumes. Crop Sci. 49, 1511-1522. [ Links ]
He, L., Zhou, W., Wang, Y., Wang, C., Chen, X. & Zhang, Q., 2018. Effect of applying lactic acid bacteria and cellulase on the fermentation quality, nutritive value, tannins profile and in vitro digestibility of Neolamarckia cadamba leaves silage. J. Anim. Physiol. Anim. Nutr. 00, 1-8. [ Links ]
He, L.W., Meng, Q.X., Li, D.Y., Wang, F. & Ren, L.P., 2015. Effect of steam explosion on in vitro gas production kinetics and rumen fermentation profiles of three common straws. Italian J. Anim. Sci. 14, 4076-4082. [ Links ]
Herremans, S., Decruyenaere, V., Beckers, Y. & Froidmont, E., 2019. Silage additives to reduce protein degradation during ensiling and evaluation of in vitro ruminal nitrogen degradability. Grass Forage Sci. 74, 86-96. [ Links ]
Huang, Q.Q, 2016. Characterization of condensed tannins in purple prairie and determination of its effects on digestive characteristics of lambs. Northwest Agricultural and Forestry University, Yangling, China (doctoral dissertation). [ Links ]
Huhtanen, P., Rinne, M. & Nousiainen, J., 2007. Evaluation of the factors affecting silage intake of dairy cows: a revision of the relative silage dry-matter intake index. Animal 1, 758-770. [ Links ]
Hvelplund, T. & Madsen, J., 1990. The amount of absorbed nutrients from the gastro-intestinal tract of dairy cows. Beretning Fra Statens Husdyrbrugsfors0g. pp. 12-17. [ Links ]
Kung, L., Stokes, M.R. & Lin, C.J., 2003. Silage additives. In: D.R. Buxton, R.E. Muck & J.H. Harrison. Silage Science and Technology. Agronomy Series 42, 305-360. American Society of Agronomy. Doi:10.2134/agronmonogr42.c7 [ Links ]
Li, P., Ji, S., Hou, C., Tang, H., Wang, Q. & Shen, Y., 2016. Effects of chemical additives on the fermentation quality and N distribution of alfalfa silage in south of China. Anim. Sci. J. 87, 1472-1479. [ Links ]
Licitra, G., Hernandez, T.M. & Van Soest, P.J., 1996. Standardization of procedures for nitrogen. Anim. Feed Sci. Technol. 57, 347-358. [ Links ]
McDonald, P., Henderson, A.R. & Heron, S.J.E., 1991. The Biochemistry of Silage. Chalcombe, Kingston, Kent, UK. [ Links ]
Menke, K. & Steingass, H., 1988. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 28, 7-55. [ Links ]
Min, M., Zhao, W. & Li, C., 2015. The dynamic analysis of nutrient and tannin content of five kinds of available forage in Changbai Mountain region. Feed Industry 21, 12-19. [ Links ]
Minnee, E., Woodward, S., Waghorn, G. & Laboyrie, P., 2002. The effect of ensiling forage legumes on condensed tannins. Agronomy 32, 117-119. [ Links ]
Ni, K., Wang, F., Zhu, B., Yang, J., Zhou, G., Pan, Y. & Zhong, J., 2017. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresource Technol. 238, 706-714. [ Links ]
NRC, 2000. Nutrient Requirements of Beef Cattle: Update 2000. National Academy Press, Washington, DC, USA. [ Links ]
Piluzza, G., Sulas, L. & Bullitta, S., 2014. Tannins in forage plants and their role in animal husbandry and environmental sustainability: A review. Grass Forage Sci. 69, 32-48. [ Links ]
Tabacco, E., Borreani, G., Crovetto, G. M., Galassi, G., Colombo, D. & Cavallarin, L., 2006. Effect of chestnut tannin on fermentation quality, proteolysis, and protein rumen degradability of alfalfa silage. J. Dairy Sci. 89, 4736-4746. [ Links ]
Terrill, T.H., Windham, W.R., Evans, J.J. & Hoveland, C.S., 1990. Condensed tannin concentration in Sericea lespedeza as influenced by preservation method. Crop Sci. 30, 219-224. [ Links ]
Wang, F., 2016. Establishment and evaluation of effective energy prediction model in feeds of beef cattle. Doctoral dissertation, China Agricultural University, Beijing, China. [ Links ]
Wang, J., Wang, J.Q., Zhou, H. & Feng, T., 2009. Effects of addition of previously fermented juice prepared from alfalfa on fermentation quality and protein degradation of alfalfa silage. Anim. Feed Sci. Technol. 151, 280-290. [ Links ]
Wang, S., Liu, G., Li, Y., Cui, Z., Zhou, D., Liu, D. & Sun, B., 2017. Effect of different proportion silage Anthocephalus chinensis substitute silage whole plant corn on growth performance, slaughter performance and meat quality of Lezhi Black goat in fattening period. Feed Industry 12, 23-30. [ Links ]
Wang, Y., Wang, X.K., Zhou, W., Yang, F.Y., Zhang, Q. & Chen, X.Y., 2018. Effects of moisture content and additives on the quality of Neolamarckia cadamba leaves silage. J. South China Agr. Uni. 39, 80-86. [ Links ]
Zayed, M.Z., Ho, W.S., Pang, S.L. & Ahmad, F.B., 2014. Comparison of mimosine content and nutritive values of Neolamarckia cadamba and Leucaena leucocephala with Medicago sativa as forage quality index. Int. J. Sci. Technol. Res. 3, 146-150. [ Links ]
Zhao, J., Dong, Z., Li, J., Chen, L., Bai, Y., Jia, Y. & Shao, T., 2018. Ensiling as pretreatment of rice straw: The effect of hemicellulose and Lactobacillus plantarum on hemicellulose degradation and cellulose conversion. Bioresource Technol. 266, 158-165. [ Links ]
Zhang, M., 2018. Effects of different planting densities on yield and fodder quality of Neolamarckia cadamba. South China Agricultural University, Guangzhou, China (Master's thesis). [ Links ]
Zhang, Q., Yu, Z., Yang, H. & Na. R.S., 2016. The effects of stage of growth and additives with or without cellulase on fermentation and in vitro degradation characteristics of Leymus chinensis silage. Grass Forage Sci. 71, 595-606. [ Links ]
Received 5 July 2018
Accepted 23 May 2019
First published online 9 July 2019
# Corresponding author: xychen@scau.edu.cn


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}