










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 no.4 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i4.3
ARTICLES
Aluminosilicates and yeast-based mycotoxin binders: Their ameliorated effects on growth, immunity and serum chemistry in broilers fed aflatoxin and ochratoxin
M.Y. FarooquiI; A. KhaliqueI, #; M.A. RashidI; S. MehmoodII; M.I. MalikI
IDepartment of Animal Nutrition, University of Veterinary and Animal Sciences, Faculty of Animal Production and Technology, 54000 Lahore, Pakistan
IIDepartment of Poultry Production, University of Veterinary and Animal Sciences, Faculty of Animal Production and Technology, 54000 Lahore, Pakistan
ABSTRACT
The aim of this study was to evaluate the effects of commercial toxin binders on growth performance, organ morphology, immunity and serum biochemistry in broilers. Dietary treatments consisted of the negative control (NC): experimental diet with aflatoxin B1 <20 parts per billion (ppb), ochratoxin A (OTA) <5 ppb; control (C) experimental diet without toxin binder; Z1: toxin binder 1 g/kg of zeta plus; Z2: toxin binder 2 g/kg of zeta plus; TX1: toxin binder 1 g/kg of Toxfin Dry; and TX2: toxin binder 2 g/kg of Toxfin Dry). Except for NC, all diets contained 57 ppb aflatoxin B1 and 23 ppb ochratoxin A. Feed intake was higher in the TX1, TX2, NC, Z2 and Z1 treatments than in the control. Weight gain was higher in Z2, TX2, Z1, TX1 and NC than in C. Feed conversion ratio (FCR) was poor in C. The control had the highest liver weight, though the weights of the spleen, kidneys and hearts of the birds were similar in all treatments. Gizzard weight, thymus weight, and bursa of Fabricius were lowest in C. The weight of the pancreas was similar among treatments. The antibody titres against new castle disease were higher in treatments Z2, Z1, TX2, TX1 and NC than in C. Urea and creatinine concentrations, and aspartate aminotransferase activity in serum were similar among treatments, whereas the serum alanine transaminase activity was higher in C than in Z1, TX1, TX2, Z2 and NC. It was concluded that growth rate, FCR and immunity indices were improved in broilers fed toxin binder. At lower levels of mycotoxin in feed, 1 g/kg of toxin binder (clay based or yeast based) was sufficient to ameliorate the adverse effects of aflatoxin B1 and OTA, whereas at higher levels of mycotoxins, supplementation of toxin binder should be increased.
Keywords: alanine aminotransferase, aspartate aminotransferase, carcass characteristics, growth performance, toxin binders, urea, creatinine
Introduction
Mycotoxins are secondary metabolites of several fungal species that contaminate almost all kinds of food before and post-harvest (Bhatti et al., 2016). These mycotoxins (T-2 toxin, deoxynivalenol, aflatoxins, fumonisins, ochratoxin A (OTA), patulin and zearalenone) are usually stable under varying environmental conditions, and exert toxic effects in poultry (Chen et al., 2008) by depressing immunity and decreasing productivity (Yegani et al., 2006; Manafi, 2012). Numerous researchers have shown that OTA decreases animal growth and production (Huff et al., 1988; Aravind et al., 2003). Furthermore, OTA has immunosuppressive properties in broiler chicks (Singh et al., 1990). Similarly, aflatoxin has adverse effects on broiler growth. Aflatoxin B1 is considered the most potent hepatotoxin, and is well known owing to its carcinogenicity (Wilson & Payne, 1994).
To eliminate, inactivate and minimize the adverse effects of mycotoxins certain strategies have been developed, for example, irradiation, physical separation, ozonization and chemical treatments of contaminated food and feed (Goldblatt & Dollear, 1979). However, one of the best strategies in terms of cost and effectiveness on a large scale is the adsorption of mycotoxin molecules with toxin binder. The bound mycotoxin molecule is then excreted in the faeces (Bintvihok et al., 2002). Amelioration of mycotoxin with toxin binders is common practice in the poultry industry. Commercially, two types of toxin binders are available, namely, clay-based and yeast-based. These toxin binders form a complex with a mycotoxin so that absorption of toxin from the intestine is restricted to the maximum extent (Dos Anjos et al., 2015). Researchers have found that smectite clay and bentonite-montmorillonite efficiently avert digestive absorption of mycotoxins of feed origin, ensuring food safety and animal health (Saleemi et al., 2015). Yeast-based toxin binders harbour polysaccharides, lipids and protein fractions. These fractions of the yeast cell wall are capable of adsorbing mycotoxin owing to the availability of adsorption sites (Kolosova & Stroka, 2012). These substances are non-digestible and are claimed to be efficient in binding with mycotoxins in the small intestines when mixed with feed. The current experiment was therefore designed to evaluate the effectiveness of commercial toxin binders, Toxfin Dry (clay) and zeta plus (clay + yeast), in broilers fed mycotoxin contaminated feed.
Materials and Methods
The experiment was conducted at Poultry Research and Training Centre (PRTC) Ravi Campus UVAS, Pattoki. The experimental procedures were in accordance with the Animal Care and Use Committee University of Veterinary and Animal Sciences, Lahore. Four hundred (400) day-old chicks were procured from a commercial hatchery. Up to day 7, the chicks were reared in a tunnel-ventilated broiler house under standardized farm practices without any supplementation. At the start of day 8, 360 birds were randomly assigned to six treatments. Each treatment had six replicates with 10 birds per replicate (see Table 1). Old corn that was contaminated with a mycotoxin and uncontaminated fresh corn were selected. The concentration of aflatoxin B1 and OTA were analyzed in the corn samples.

After confirmation of the laboratory results, six isonitrogenous and isoenergetic diets (Table 2) were formulated and analyzed for aflatoxin B1 and OTA. These treatments consisted of negative control (NC), an experimental diet with aflatoxin B1 at <20 ppb, OTA at <5 ppb; the control (C) diet with aflatoxin B1 at 57 ppb, and OTA at 23 ppb; Z1: aflatoxin B1 at 57 ppb and OTA at 23 ppb + toxin binder 1 g/kg of zeta plus; Z2: aflatoxin B1 at 57 ppb and OTA ppb 23 + toxin binder 2 g/kg of zeta plus; TX1: aflatoxin B at 57 ppb and OTA at 23 ppb + toxin binder 1 g/kg of Toxfin Dry, and TX2 aflatoxin B1 at 57 ppb and OTA at 23 ppb + toxin binder 2 g/kg of Toxfin Dry. All experimental diets were fed to broiler from days 8 to 35 (21% crude protein and 2900 kcal metabolizable energy/kg). The birds were kept on a floor that was covered with litter at a stocking density of 0.65 ft2 per bird. The light was provided for 23 hours. The birds had open access to fresh water and were offered with feed ad libitum. Temperature and humidity in the house were provided according to the Ross management guides. The broilers were vaccinated according to the breed management guide. At the end of every week, feed intake and bodyweight were recorded and FCR was calculated.
At day 35, three birds from each replicate were randomly selected and blood was collected via cardiac puncture in vacutainers without any anticoagulant. Immediately after blood collection birds were slaughtered humanely and organs were weighed. To evaluate the adverse effects of mycotoxins on the visceral organs of the birds, the hearts, liver, bursa of Fabricius, gizzards, kidney, spleen and thymus were weighed with a digital weighing balance. Serum was harvested with a centrifuge (3000 g for 15 min at 23 °C). The aliquots of the serum were labelled and stored at -20 °C. Serum samples were sent to University Diagnostic Lab, UVAS, Lahore, to determine the concentrations of creatinine and urea, activities of alanine amino transferase (ALT, EC 2.6.1.2) and aspartate aminotransferase (AST, EC 2.6.1.1), and antibody titres against new castle disease (ND) were also analyzed from the collected serum.
The experimental design was completely randomized. Data for feed intake, body weight and FCR, organ weight, immunity and blood biochemistry were analyzed with one-way ANOVA using statistical package, SAS 2002. Means were compared with Duncan's multiple range (DMR) test (Duncan, 1955) at P <0.05 significance levels.
Results
The least square means of feed intake, weight gain and FCR are presented in Table 3. The results revealed that feed intake was higher (P <0.05) in TX1, TX2, NC, Z2 and Z1 than in C. Weight gain was lower (P <0.05) in C than in Z2, TX2, Z1, TX1 and NC. Similarly, the poorest (P <0.05) FCR was observed in C.
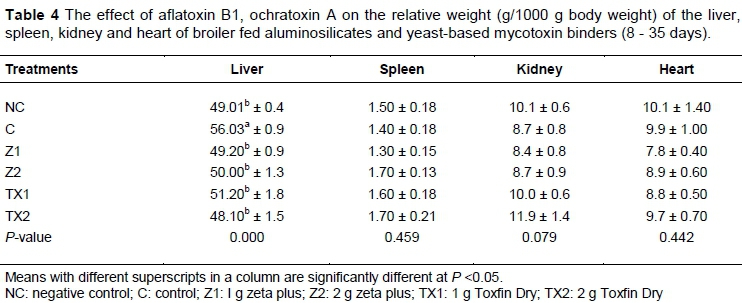
The least mean squares of organ weight are presented in Table 4. The results showed that the C treatment had highest (P <0.05) liver weight, followed by TX1, Z2, Z1, NC and TX2. The weights of spleen, kidneys and heart of birds were not affected (P >0.05) by dietary treatments.
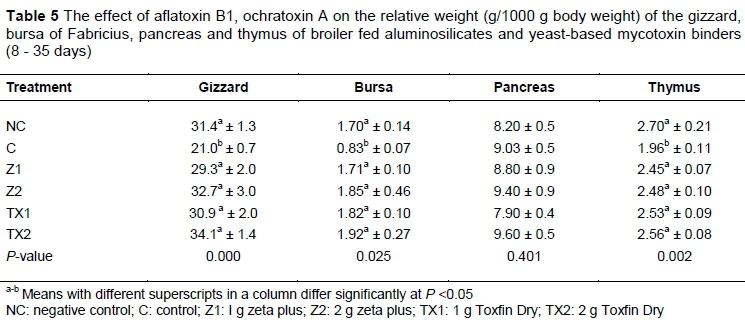
The least square means of gizzard, bursa, pancreas and thymus are presented in Table 5. The gizzard weight of the C was lower (P >0.05) than that of the Z1, TX1, NC, Z2 and TX2 treatments. The weight of the bursa of Fabricius was lowest (P <0.05) in C compared with TX2, Z2, TX1, Z1 and NC. The weight of the pancreas was similar (P >0.05) among all treatments. The thymus weight was lowest (P <0.05) in C compared with all treatments. However, the rest of the treatments showed similar thymus weights.
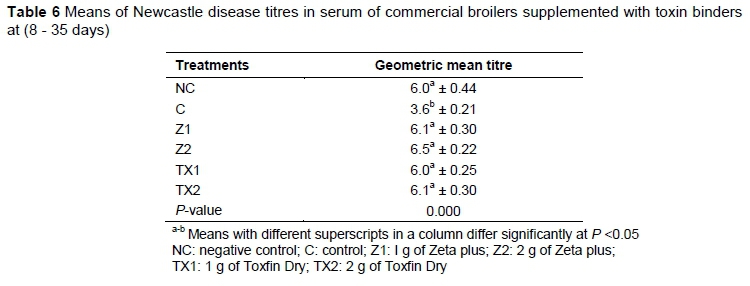
Titres against the ND virus are shown in Table 6. The antibody titre against ND was higher (P <0.05) in Z2, Z1, TX2, TX1 and NC compared with C.
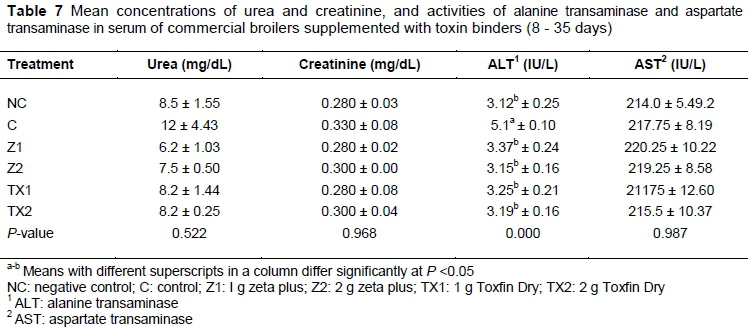
The means of urea, creatinine concentrations, and ALT and AST activities are presented in Table 7. The results of the present study show that urea, creatinine and AST were similar (P >0.05) among treatments, whereas serum ALT activity was higher in C (P <0.05) compared with the Z1, TX1, TX2, Z2, and NC.
Discussion
The results of the current experiment shown that feed intake was lower in control (C) compared with other treatments. Lower intake may be associated with a higher level of mycotoxins in C. Similar findings were reported by Verma et al. (2002), that mycotoxin in feed resulted in reduced feed intake in broilers. Huff et al. (1975) reported that ochratoxin at >20 ppb compromised feed intake owing to an adverse effect on the gizzard. A synergistic effect of aflatoxin and ochratoxin may aggravate feed intake depression (Appleton & Campbell, 1983; Huff, 1991). Most of the synergistic toxicity of two toxins exert serious adverse effects on feed intake compared with a single toxin (Huff et al., 1988). These findings were in accordance with those of Denli et al. (2009), who observed lower feed intake when aflatoxin was given in the feed. Similarly, Doerr et al. (1974) reported that the feed intake of birds exposed to ochratoxin was decreased owing to its nephrotoxic effect. Kubena et al. (1994) also reported that aflatoxin and ochratoxin both depress feed intake by compromising the enzyme and excretory systems of birds. Although Z1, Z2, TX1 and TX2 were also fed higher levels of mycotoxins, feed intake was higher compared with C. This higher intake may be associated with neutralization of mycotoxins by toxin binders. Neutralization of mycotoxins by toxin binder was also reported by (Ibrahim et al., 2000). Gowda et al. (2008) recorded that the addition of sodium calcium aluminosilicates to the aflatoxin B1 diet significantly enhanced the feed intake.
The results of the current study show that Z2, TX2, Z1, TX1 and NC had higher body weight than C. Higher body weight is associated with higher feed intake in these treatments. Similar findings were reported by Huff et al. (1988), in which they studied combined toxins that reduced weight gain, which might be because of impairment of protein synthesis. A large reduction in body weight because of a diet that was contaminated with OTA at levels of 400 and 800 ppb was also reported by Elaroussi et al. (2006). Treatments that contain toxin binder have higher body weight at day 35 compared with those fed only toxins. Similar findings were reported by Santin et al. (2002), in that broiler chicks exposed to OTA at a level of 250 or 500 ppb during the initial stage (1 to 21 days), the final stage (22 to 42 days), or in the total experimental period (1 to 42 days) had lower average body weight compared with the control. Significantly, greater body weight gain was observed in the birds that were fed the clay-based toxin binder in the present study. A similar response was observed in weight gain, FCR, organ weight and immunity parameter in all treatments that were fed toxin binder as feed supplement, which may be associated with lower level of mycotoxin (aflatoxin 57 ppb and ochratoxin 23 ppb) in feed compared with Elaroussi et al. (2006). The current results are in line with the findings of Kubena et al. (1994), who showed reduced weight gain owing to toxins in feed. Clay-based toxin binder reduced the effect of toxin in the feed by binding the toxin from the intestine and providing stability to the intestinal wall (Manafi, 2012). Aluminosilicates bind with aflatoxin molecules, which cannot be absorbed from the animal's digestive tract. The bound aflatoxins are then excreted in the faeces (Bintvihok et al., 2002).
The current results showed that FCR was poor in C compared with Z2, TX2, TX1, Z1 and NC. Poor FCR is associated with lower feed intake and lower weight gain in C. Santin et al. (2002) reported that ochratoxin influenced the FCR negatively in the broiler chicks. Poor FCR in a diet with mycotoxins could be attributed to hepatic cell damage and decreased pancreatic enzymes. The groups in which toxin binder was added, showed better FCR compared with the contaminated group. This was similar to the results recorded by Harvey et al. (1992). The combination of 1 mg/kg aflatoxin and 2 mg/kg ochratoxin showed a significant decrease in FCR compared with the control (Verma et al., 2004).
Aflatoxin and ochratoxin have adverse effects on the weight of the liver. The highest weight of liver in C indicated that it was associated with mycotoxins in feed, namely, those fed 1 g/kg or 2 g/kg of toxin binder recovered better compared with C (without toxin binder). Similar findings were reported by Kermanshahi et al. (2007) that enlargement and discoloration of the liver were associated with aflatoxin. The concentration of aflatoxin at 1 mg/kg or more caused a significant reduction in immune status and growth in broilers, whereas 2 mg/kg of aflatoxin may increase liver weight (Verma et al., 2004). Che et al. (2011) reported that diet contaminated with toxins results in an adverse effect on liver morphology. Contamination of feed with mycotoxins increases lipid deposition in the liver and weight also increases (Sharlin et al., 1980). Clay-based toxin binders are used to decrease the effects of toxins, particularly ochratoxin, in broilers. Similarly, they help to improve diet absorption and decreases the toxin effects on the liver (Santin et al., 2003), whereas spleen, kidney and heart were not affected by the toxin binder.
The weights of gizzard and bursa of Fabricius were lower in C than in NC and in those fed toxin binders. Similar findings by Verma et al. (2002) stated that combining aflatoxin and OTA 2 mg/kg and 4 mg/kg, respectively, led to a decrease in gizzard weight. The reduction in the sizes of these organs might have been due to necrosis and cellular depletion by the mycotoxins (Kubena et al.,1990; Hoerr, 2010).
Elaroussi et al. (2006) argued that the reduced size of the bursa was due to mycotoxin. Immunosuppression was the major alteration that was associated with the mycotoxin, which might occur because of the reduced size of immunological organs such as the bursa and thymus.
Group C had the lowest antibody titre against ND compared with NC and other treatments. This was associated with the immune suppression effects of mycotoxins. Similar results were demonstrated by Sharma et al. (2008). A decrease in antibody production in the current experiment was linked to regression of bursa of Fabricius (Ibrahim et al., 1998) in that mycotoxin causes bursal regression and suppression of the primary immune response. Antibody titre was higher in Z2, TX2, Z1, TX1 and NC. The result of the present study is in the line with those of Ibrahim et al. (2000), who reported that significant addition of toxin binder could ameliorate the adverse effects of mycotoxins, especially aflatoxin. Toxin binders in the diet help to reduce the absorption of toxin from the gut (Magnoli et al., 2011). Higher antibody titre is due to the binding ability and lower absorption of toxin from gut with mycotoxins (Weiner, 1997). Clay-based toxin binder also stimulates the indirect activation of B cells, T cells and macrophages (Lyons & Jacques, 1994).
These results were in agreement with those of Azzam & Gabal (1998), who reported a decrease in the antibody titre because of the mycotoxin. Antibody titre against Newcastle disease virus is normal in chickens exposed to low levels of dietary aflatoxin (10 - 15 ppb), but higher levels of mycotoxins (50 - 60 ppb) without toxin binder can suppress immunity (Corrier, 1991). Hence aflatoxin and ochratoxin individually or in combination are growth depressant and immunosuppressant. In agreement with the conclusion drawn by Mohiuddin & Reddy (1993), it might be suggested that the immunosuppressive effects of mycotoxin in feeds could be ascribed mostly to vaccine failure, leading to high morbidity, mortality and heavy economic losses.
Hepatic function and activity are measured by serum biochemical parameters, which include AST and ALT, creatinine, and urea. The results of the current study showed that urea, creatinine and AST were similar among all treatments. The highest activity of ALT was observed in C. Similarly, Sharma et al., (2008) reported that serum ALT activity was higher in broilers fed a diet that was contaminated with aflatoxin. Similar results also reported by Gowda et al. (2008), which suggested that mycotoxin exerts a direct effect on the animal's liver, which alters enzyme secretion from the body. Elevated serum ALT activity in the groups fed OTA showed degenerative changes, which lead to the seepage of enzymes from the liver. These findings are in accord with those of Sathyanarayana et al. (2004). Santin et al. (2002) and Kumar et al. (2003) fed broiler chicks from hatching through five weeks old a diet supplemented with 2 ppm OTA to study its toxic effect on broiler performance. OTA negatively influenced the serum activities of liver enzymes.
Conclusion
By keeping the results of the current experiment in mind, it can be concluded that growth rate, FCR, and immunity parameters were improved in those fed toxin binders. The similar response among treatments is associated with the level of mycotoxin, which was lower in the current experiment than those reported in the literature. Supplementation of 2 g/kg of yeast-based mycotoxin can be a successful strategy to ameliorate the adverse effects of aflatoxin B1 and OTA. So, at a lower level of mycotoxin in feed, 1 g/kg of toxin binder (clay based or yeast based) is enough to ameliorate the adverse effects of aflatoxin B1 and OTA.
Acknowledgment
The authors are thankful to the Kemin Industry, Pakistan, for provision of a research grant.
Authors' Contributions
YF: data collection; AK: experiment planning and write up; MAR: write up and Lab analysis; SM: statistical analysis; and MIM: formatting and write up.
Conflict of Interest Declaration
The authors declare there is no conflict of interest.
References
Appleton, B.S. & Campbell, T.C., 1983. Effect of high and low dietary protein on the dosing and postdosing periods of aflatoxin B1-induced hepatic preneoplastic lesion development in the rat. Cancer Res. 43, 2150-2154. [ Links ]
Aravind, K., Patil, V., Devegowda, G., Umakantha, B. & Ganpule, S., 2003. Efficacy of esterified glucomannan to counteract mycotoxicosis in naturally contaminated feed on performance and serum biochemical and hematological parameters in broilers. Poult. Sci. 82, 571-576. [ Links ]
Azzam, A. & Gabal, M., 1998. Aflatoxin and immunity in layer hens. Avian Pathol. 27, 570-577. [ Links ]
Bhatti, S.A., Khan, M.Z., Saleemi, M.K. & Saqib, M., 2016. Aflatoxicosis and ochratoxicosis in broiler chicks and their amelioration with locally available bentonite clay. Pak. Vet. J. 36, 68-72. [ Links ]
Bintvihok, A., Thiengnin, S., DOI, K. & Kumagai, S., 2002. Residues of aflatoxins in the liver, muscle and eggs of domestic fowls. J. Vet. Med. Sci. 64, 1037-1039. [ Links ]
Che, Z., Liu, Y., Wang, H., Zhu, H., Hou, Y. & Ding, B., 2011. The protective effects of different mycotoxin adsorbents against blood and liver pathological changes induced by mold-contaminated feed in broilers. Asian-Austral. J. Anim. Sci. 24, 250-257. [ Links ]
Chen, F., Ma, Y., Xue, C., Ma, J., Xie, Q., Wang, G., Bi, Y. & Cao, Y., 2008. The combination of deoxynivalenol and zearalenone at permitted feed concentrations causes serious physiological effects in young pigs. J. Vet. Sci. 9, 39-44. [ Links ]
Corrier, D., 1991. Mycotoxicosis: mechanisms of immunosuppression. Vet. Immunol. Immunopathol. 30, 73-87. [ Links ]
Denli, M., Blandon, J., Guynot, M., Salado, S. & Perez, J., 2009. Effects of dietary AflaDetox on performance, serum biochemistry, histopathological changes, and aflatoxin residues in broilers exposed to aflatoxin B1. Poult. Sci. 88, 1444-1451. [ Links ]
Doerr, J., Huff, W., Tung, H., Wyatt, R. & Hamilton, P., 1974. A survey of T-2 toxin, ochratoxin, and aflatoxin for their effects on the coagulation of blood in young broiler chickens. Poult. Sci. 53, 1728-1734. [ Links ]
Dos Anjos, F., Ledoux, D., Rottinghaus, G. & Chimonyo, M., 2015. Efficacy of adsorbents (bentonite and diatomaceous earth) and turmeric (Curcuma longa) in alleviating the toxic effects of aflatoxin in chicks. Br. Poult. Sci. 56, 459-469. [ Links ]
Duncan, D.B., 1955. Multiple range and multiple F tests. Biometrics 11, 1-42. [ Links ]
Elaroussi, M., Mohamed, F., El Barkouky, E., Atta, A., Abdou, A. & Hatab, M., 2006. Experimental ochratoxicosis in broiler chickens. Avian Pathol. 35, 263-269. [ Links ]
Goldblatt, L. & Dollear, F., 1979. Modifying mycotoxin contamination in feeds-Use of mold inhibitors, ammoniation, roasting. Interactions of mycotoxins in animal production. National Academy of Sciences, Washington, DC, USA. pp. 167-184. [ Links ]
Gowda, N., Ledoux, D., Rottinghaus, G., Bermudez, A. & Chen, Y., 2008. Efficacy of turmeric (Curcuma longa), containing a known level of curcumin, and a hydrated sodium calcium aluminosilicate to ameliorate the adverse effects of aflatoxin in broiler chicks. Poult. Sci. 87, 1125-1130. [ Links ]
Harvey, R., Elissalde, M., Kubena, L., Weaver, E., Corrier, D. & Clement, B., 1992. Immunotoxicity of ochratoxin A to growing gilts. Am. J. Vet. Res. 53, 1966-1970. [ Links ]
Hoerr, F.J., 2010. Clinical aspects of immunosuppression in poultry. Avian Dis. 54, 2-15. [ Links ]
Huff, J., 1991. Carcinogenicity of ochratoxin A in experimental animals. IARC Sci. Publ. 115, 44. [ Links ]
Huff, W., Wyatt, R. & Hamilton, P., 1975. Effects of dietary aflatoxin on certain egg yolk parameters. Poult. Sci. 54, 2014-2018. [ Links ]
Huff, W., Kubena, L. & Harvey, R., 1988. Progression of ochratoxicosis in broiler chickens. Poult. Sci. 67, 1139-1146. [ Links ]
Ibrahim, I., Al-Joubory, K. & Shareef, A., 1998. Reducing aflatoxicosis in growing chicks by dietary sodium bentonite. IPA J. Agri. Res. 8, 130-138 [ Links ]
Ibrahim, I., Shareef, A. & Al-Joubory, K., 2000. Ameliorative effects of sodium bentonite on phagocytosis and Newcastle disease antibody formation in broiler chickens during aflatoxicosis. Res. Vet. Sci. 69, 119-122. [ Links ]
Kermanshahi, H., Akbari, M.R., Maleki, M. & Behgar, M., 2007. Effect of prolonged low level inclusion of aflatoxin B 1 into diet on performance, nutrient digestibility, histopathology and blood enzymes of broiler chickens. J. Anim. Vet. Adv. 6, 686-692. [ Links ]
Kolosova, A. & Stroka, J., 2012. Evaluation of the effect of mycotoxin binders in animal feed on the analytical performance of standardised methods for the determination of mycotoxins in feed. Food Addit. Contam: Part A 29, 1959-1971. [ Links ]
Kubena, L., Harvey, R., Huff, W., Corrier, D., Phillips, T. & Rottinghaus, G., 1990. Efficacy of a hydrated sodium calcium aluminosilicate to reduce the toxicity of aflatoxin and T-2 toxin. Poult. Sci. 69, 1078-1086. [ Links ]
Kubena, L., Harvey, R., Edrington, T. & Rottinghaus, G., 1994. Influence of ochratoxin A and diacetoxyscirpenol singly and in combination on broiler chickens. Poult. Sci. 73, 408-415. [ Links ]
Kumar, A., Jindal, N., Shukla, C., Pal, Y., Ledoux, D. & Rottinghaus, G., 2003. Effect of ochratoxin A on Escherichia coli-challenged broiler chicks. Avian Dis. 47, 415-424. [ Links ]
Lyons, T.P. & Jacques, K., 1994. Biotechnology in the Feed Industry. Proceedings of Alltech's 10th Annual Symposium, Biotechnology in the feed industry. Proceedings of Alltech's 10th Annual Symposium., Alltech UK. pp. 1-48. [ Links ]
Magnoli, A., Texeira, M., Rosa, C., Miazzo, R., Cavaglieri, L., Magnoli, C., Dalcero, A. & Chiacchiera, S., 2011. Sodium bentonite and monensin under chronic aflatoxicosis in broiler chickens. Poult. Sci. 90, 352-357. [ Links ]
Manafi, M., 2012. Counteracting effect of high grade sodium bentonite during aflatoxicosis in broilers. J. Agri. Sci. Technol. 14, 539-547. [ Links ]
Mohiuddin, S. & Reddy, M., 1993. Immunosuppresive effects of aflatoxin in bursectomised chicks against Ranikhet disease vaccine. Indian J. Anim. Sci. (India). 63, 279-280. [ Links ]
Saleemi, M.K., Khan, M.Z., Khan, A., Ul Hassan, Z., Khan, W.A., Rafique, S., Fatima, Z. & Sultan, A., 2015. Embryotoxic and histopathological investigations of in-ovo inoculation of aflatoxigenic fungal extracts in chicken embryos. Pakistan Vet. J. 35, 403-408. [ Links ]
Santin, E., Maiorka, A., Krabbe, E., Paulillo, A. & Alessi, A., 2002. Effect of hydrated sodium calcium aluminosilicate on the prevention of the toxic effects of ochratoxin. J. Appl. Poult. Res. 11, 22-28. [ Links ]
Santin, E., Paulillo, A.C., Maiorka, A., Nakaghi, L.S.O., Macari, M., Silva, A. & Alessi, A.C., 2003. Evaluation of the efficacy of Saccharomyces cerevisiae cell wall to ameliorate the toxic effects of aflatoxin in broilers. Int. J. Poult. Sci. 2, 341-344. [ Links ]
Sathyanarayana, M., Vijayasarathi, S. & Rao, S., 2004. Pathomorphological changes in broiler chickens exposed to individual and combined toxicity of aflatoxin and ochratoxin, and assessment of efficacy of a toxin binder. Indian J. Anim. Sci. 74, 747-749. [ Links ]
Sharlin, J., Howarth Jr, B. & Wyatt, R., 1980. Effect of dietary aflatoxin on reproductive performance of mature White Leghorn males. Poult. Sci. 59, 1311-1315. [ Links ]
Sharma, D., Asrani, R., Ledoux, D., Jindal, N., Rottinghaus, G. & Gupta, V., 2008. Individual and combined effects of fumonisin B1 and moniliformin on clinicopathological and cell-mediated immune response in Japanese quail. Poult. Sci. 87, 1039-1051. [ Links ]
Singh, G., Chauhan, H., Jha, G. & Singh, K., 1990. Immunosuppression due to chronic ochratoxicosis in broiler chicks. J. Comp. Pathol. 103, 399-410. [ Links ]
Verma, J., Swain, B.K. & Johri, T.S., 2002. Effect of various levels of aflatoxin and ochratoxin A and combinations thereof on protein and energy utilisation in broilers. J. Sci. Food Agri. 82, 1412-1417. [ Links ]
Verma, J., Johri, T., Swain, B. & Ameena, S., 2004. Effect of graded levels of aflatoxin, ochratoxin and their combinations on the performance and immune response of broilers. Br. Poult. Sci. 45, 512-518. [ Links ]
Weiner, H.L., 1997. Oral tolerance: immune mechanisms and treatment of autoimmune diseases. Immunol. Today 18, 335-343. [ Links ]
Wilson, D.M. & Payne, G.A., 1994. Factors Affecting the Toxicology of Aflatoxins: Human Health, Veterinary, and Agricultural Significance. Elsevier. [ Links ]
Yegani, M., Smith, T., Leeson, S. & Boermans, H., 2006. Effects of feeding grains naturally contaminated with Fusarium mycotoxins on performance and metabolism of broiler breeders. Poult. Sci. 85, 1541-1549. [ Links ]
Received 27 August 2018
Accepted 1 April 2019
First published online 8 July 2019
# Corresponding author: akhalique@uvas.edu.pk


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}