










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 n.4 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i4.1
ARTICLES
Investigating novelty traits to improve cow-calf efficiency in South African Afrikaner, Angus and Charolais for climate-smart production
M.C. MokolobateI, II, #; M.M. ScholtzI, II; F.W.C NeserII
IARC-Animal Production Institute, Private Bag X2, Irene, 0062, South Africa
IIUniversity of the Free State, Bloemfontein, 9300, South Africa
ABSTRACT
The aim is of the study was to identify novelty traits that could be used as breeding objectives to improve cow-calf efficiency and describe cow efficiency in extensive systems in support of climate-smart production in beef cattle. The traits that were investigated were 'weaning weight of the calf as a trait of the dam' (K205) and 'kilogram calf weaned per large stock unit' (KgC/LSU. The latter trait is a value that expresses performance (calf weaning weight) per constant unit, namely per LSU. This may be a useful breeding objective or goal to increase production efficiency, which may reduce the carbon footprint of extensive beef cow-calf production systems. The investigation of the novel traits was conducted on three diverse breeds, namely Afrikaner, Angus and Charolais, with 6104, 7581 and 2291 complete cow-calf records, respectively. Only cows with all three first parities recorded were used to investigate KgC/LSU and K205, as breeding objectives to improve cow-calf efficiency. The heritabilities for KgC/LSU were 0.52, 0.24 and 0.21 for the Afrikaner, Angus and Charolais, respectively, and for K205 were 0.40, 0.17 and 0.13 respectively. The genetic relationship between KgC/LSU and K205 for Angus and Charolais varied substantially. In Charolais cows a strong negative correlation (-0.75) was found, while a strong positive correlation (+0.84) was estimated in Angus cows. These results indicate that a 'cow efficiency index' in which several traits (production, fertility and efficiency) are included may be a more effective alternative breeding strategy. Breeding strategies and production systems to improve the production efficiency of beef cattle could play a significant role in reducing the carbon footprint and would enhance climate-smart beef production.
Keywords: breeding objective, calf weight, cow weight
Introduction
An effective way to reduce the carbon footprint of beef production and to support climate-smart production is to reduce cattle numbers and increase the production per animal. Increased productivity generates less greenhouse gas emission per unit of product (Scholtz et. al., 2010). It therefore becomes increasingly important to define breeding objectives and develop appropriate selection criteria and crossbreeding strategies to ensure that beef production is effective and aimed at sustainable production (climate-smart production) in changing environments. A successful and efficient cow-calf production system would be one that produces bigger weaner calves without wasting resources (feed). Efficiency of a production system essentially measures the resources that are required to produce a desired output (Maddock & Lamb, 2010). Therefore, efficiency estimations should be expressed in relation to the output attained relative to the resources utilized (Greiner, 2009). Improving cow-calf efficiency is important as it represents about 72% of maintenance requirements to produce a market-ready slaughter animal, the bulk of which are utilized by the dam (Ferrell & Jenkins, 1982). Quantifying production efficiency of cow-calf production to date is still a challenge, as most performance or improvement measurements are still per individual trait (milk production, weaning weight, calving interval, growth rate, etc.). Selection for these traits may increase production, but not necessarily efficiency of production.
A measurement is thus required that expresses output per constant (standardized) unit, such as Livestock Unit (LSU), which might be a useful breeding objective or goal to increase production efficiency. It may also reduce the carbon footprint of extensive cow-calf production systems. Since an LSU is linked to specific metabolizable energy (ME) requirements, it should be possible to eventually 'link' this breeding objective with the carbon footprint of a weaner calf production system. Selection for productivity and efficiency for instance would have a permanent mitigating effect on the production of greenhouse gases (GHG), as higher productivity would lead to higher gross efficiency as a result of reducing the maintenance cost of animals (Wall et al., 2010; Scholtz et al., 2011). Proper trait definition is therefore imperative.
In sheep, an interesting measurement of 'mother-offspring' efficiency was reported by Olivier et al. (2001), in which the reproductive performance of ewes was defined as 'total weight of lamb weaned per ewe joined' (TWW), which also demonstrated that TWW can be genetically improved by direct or indirect selection. Following a study on Dorper sheep, Van Wyk et al. (2003) recommended that the total weight of lamb weaned should be regarded as a selection criteria, since it is a composite trait that incorporates elements of the lamb and the ewe. These results, which were obtained on sheep, propose a similar initiative in beef cattle, which would be selection for 'weight of calf weaned as a trait of the dam' and this possibility should be explored as a means of improving cow efficiency.
In this study, qualities such as kilogram calf weaned per large stock unit (KgC/LSU) and weaning weight (K205), both as traits of the dam, were investigated as measures of cow-calf efficiency. No reference to such traits could be found in the literature, hence the novelty. It is also important to understand the correlations between these new traits and conventional traits, such as calf weaning weight and dam weight, which would be included in this study.
It was relevant that the results on the Bonsmara of the novel cow-calf efficiency traits from Mokolobate (2015) should be compared with those of other breeds. It was decided to use diverse breed types for this comparison. Those selected were the Afrikaner, which can be regarded as a tropical adapted taurine breed (indigenous Bos taurus africanus), Angus (British Bos taurus) and Charolais (European Bos taurus).
The Afrikaner is one of the oldest indigenous breeds in South Africa. Its history dates back to the end of the fourteenth century. The breed is closely associated with the history of the country and its people. The Afrikaner Cattle Breeder's Society, which was founded in 1912, was one of the first societies in South Africa. Over the last two decades Afrikaner breeders have focused on economically important traits, and performance recording was made compulsory, although it is not enforced (Scholtz et al., 2010). The small to medium frame size of the Afrikaner cow makes it the ideal dam line in crossbreeding for the production of heavy weaner calves (Theunissen, 2011; Mokolobate et al., 2014).
The Angus breed originated in Scotland in the 16th century and the first Angus cattle were introduced in South Africa in 1895. The Angus Society of South Africa is also one of the older breeders' societies and was founded in 1917. The Angus is the most numerous British beef breed in South Africa. Performance recording is not compulsory (Scholtz, 2010).
The Charolais has its origin in the Bresse-Plateau region in the Jura mountains of eastern France. The first Charolais cattle were imported into South Africa in 1955 and 1956, followed by a substantial import in 1962 (Bosman, 1994). In 1966 the Charolais Cattle Breeders' Society of South Africa was founded. Performance recording is not compulsory (Scholtz, 2010).
Materials and Methods
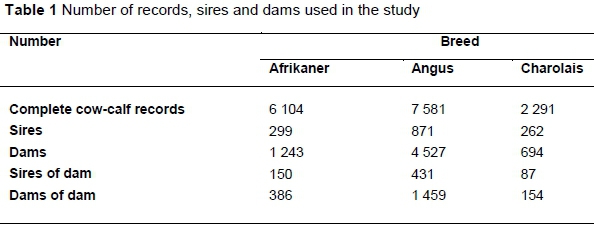
The edited dataset, as used by the Agricultural Research Council (ARC) for routine genetic evaluations, was used for this study. Herds with less than three years of recording, contemporary groups with fewer than 10 cow records and two service sires in the first parity, and animals with unknown parents were removed from the final dataset used for analysis. Only cows that had completed three parities were included in the analysis. The final datasets comprised 6104, 7581 and 2291 complete calf-cow records for the Afrikaner, Angus and Charolais breeds, respectively. The descriptions of the datasets are presented in Table 1. The relatively low number of records per breed could be contributed to insufficient data capturing on the national database or discontinued recording.
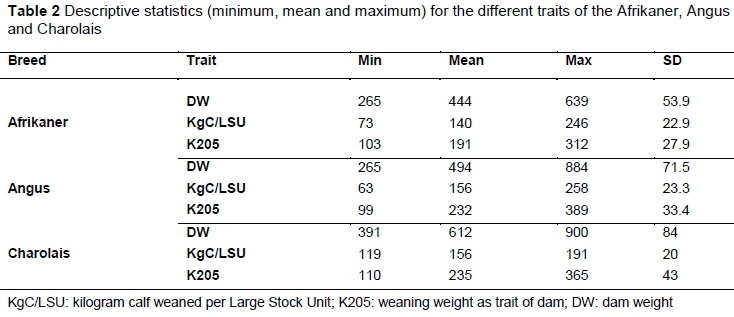
The descriptive statistics for traits that were investigated, namely dam weight (DW), K205, and KgC/LSU of the Afrikaner, Angus and Charolais are given in Table 2. In South Africa, an LSU is defined as the equivalent of an ox with a weight of 450 kg and a weight gain of 500 g per day on grass pasture with mean digestible energy concentration of 55%. To maintain this, 75 MJ metabolizable energy is required (Meissner et al., 1983). This is similar to the animal unit that is used in North America (Thorne & Stevenson, 2007). As expected, Charolais had the highest mean DW. The Angus had the highest mean KgC/LSU, followed by the Afrikaner. If KgC/LSU is taken as a definition of cow efficiency, then the cow efficiency of the Angus is 31% higher than that of the Charolais.
A number of models were tested to estimate the (co)variance components for the traits DW, KgC/LSU and K205, the latter two as traits of the dam, using repeatability models in ASReml 3.0 (Gilmour et al., 2009). The most suitable model for each trait was selected based on the log likelihood values following the same approach as Neser et al. (1996) and Mohammadi et al. (2013). The simplest model with improved (smaller) log likelihood values was chosen, since fitting other models did not change the log likelihood values favourably. Three models that were tested, including one with service sire, are presented below.
Model 1: Service sire included as additional random effect
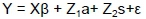
where: Y = vector of observations (DW, KgC/LSU and K205)
β = vector of fixed effects (HYS, sex) influencing KgC/LSU, K205, and DW
a = vector of direct additive effects
s = vector of additive effects related to service sire
ε = vector of residuals and where
X, Z1 and Z2 were incidence matrices relating observations to their fixed and random effects
Model 2: Single trait analysis
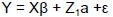
where: Y = vector of observations KgC/LSU, K205, and DW
β = vector of fixed effects (HYS, sex, damage, parity) influencing KgC/LSU, K205, and DW
a = vector of direct additive effects
ε = vector of residuals and where
X and Z1 were incidence matrices relating observations to their fixed and random effects (animal)
Model 3: Bivariate analysis
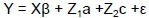
where: Y = vector of observations KgC/LSU, K205, and DW
β = vector of fixed effects (HYS and sex) influencing KgC/LSU, K205, and DW
a = vector of direct additive effects
c = vector of additional random permanent environmental effects (animal)
ε = vector of residuals and where
X and Zi were incidence matrices relating observations to their fixed and random effects (animal)
Results and Discussion
Model i, with service sire included, did not produce significantly better log likelihood values and was therefore not used. In Dorper sheep, Van Wyk et al. (2003) reported that the inclusion of service sire only had a marginal additional effect on birth and weaning weight. Model 2 was used for the estimation of variance components and heritabilities in a single trait analysis, whereas Model 3 was used to estimate the genetic correlations between the traits in a bivariate analysis.
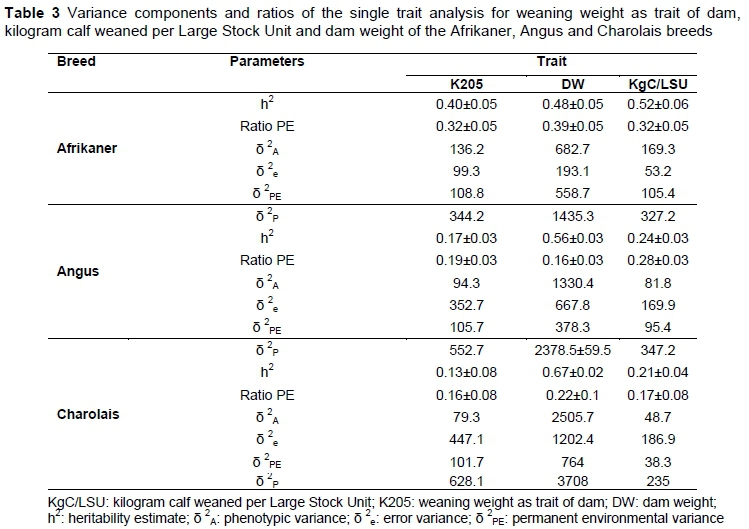
The variance components for K205, DW and KgC/LSU of the breeds are presented in Table 3. There was a vast difference in the heritability estimates between the breeds for the same trait. Heritability estimates of K205 for the three breeds ranged from 0.13 to 0.40. Since no similar trait could be found in the literature, these values had to be compared with the heritabilities of related traits. Wright et al. (1987) reported heritabilities of 0.12 and 0.09 for direct and maternal weaning weight respectively, while Van der Westhuizen et al. (2010) reported heritabilities of 0.22 and 0.12 and Neser et al. (1996) reported heritabilities of 0.13 - 0.28 and 0.14 - 0.29 for the same traits, respectively.
The heritability estimate for KgC/LSU was much higher for the Afrikaner (0.52) compared with the Angus (0.24), Charolais (0.21) and the Bonsmara (0.26), the latter from Mokolobate et al. (2015), indicating that it might be possible to drastically increase the cow efficiency in the Afrikaner. Jordaan ("2015) found that cow efficiency in the Afrikaner had increased by 18.3% over 25 years, whereas it had increased by only 10.0% in the Bonsmara. This can possibly at least in part be attributed to the higher heritability for the trait in the Afrikaner. While heritability of K205 varied across the breeds, it was the highest for the Afrikaner at 0.40. This indicates that it is possible to include K205 in a selection programme as a means of improving efficiency.
The results in this study support the findings of Mokolobate (2015) of the complexities when selecting for a ratio such as KgC/LSU, which demonstrated a high negative correlation and a high positive correlation between K205 and KgC/LSU of -0.75 and 0.84 respectively. These complexities seem to be similar to the use of other ratio traits such as calf-to-cow weight ratio, which demonstrate discrepancies when using such a trait (MacNeil, 2007).
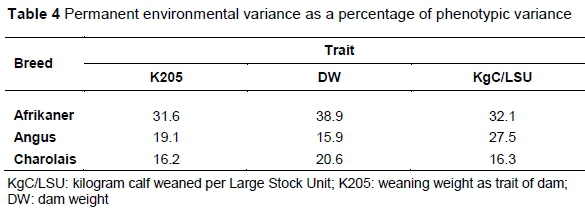
The permanent environmental variance as percentage of the phenotypic variance is summarized and presented in Table 4, since this may be an indication of the carry-over effect from one parity to the next in the repeatability model.
Table 4 shows that the permanent animal effect as percentage of phenotypic variance is much larger in the Afrikaner breed. It can be speculated that since the Afrikaner is kept under more extensive (severe) conditions than the other two breeds, this might be the cause of the larger effect.
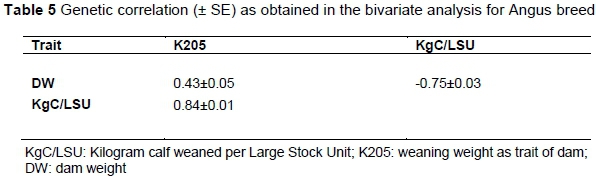
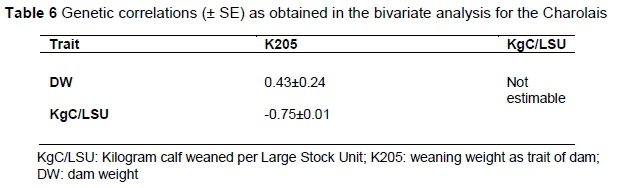
An attempt was also made to estimate the genetic correlations between the traits in question for the three breeds using the bivariate model. In the Afrikaner the analyses did not converge and therefore no genetic correlations could be estimated. The genetic correlations for the Angus and Charolais are presented in Tables 5 and 6, respectively. The genetic correlations between DW and KgC/LSU could also not be estimated in the case of the Charolais. This is possibly due to the high interdependency between the two traits and the low number of records available.
Tables 5 and 6 demonstrate a moderate but positive relationship between DW and K205 of 0.43 in both breeds. However, in the Charolais it is not significant owing to the large SE. Since K205 is a novel trait, it could not be compared with estimates from the literature, but it is similar to those between cow weight and weaning weight that were reported by Koots et al. (1994) and the 0.44 reported by Van der Westhuizen et al. (2010) between mature weight and weaning weight direct.
However, there is a major difference in the nature of the relationship between KgC/LSU and K205 in the Angus and the Charolais; the Charolais having a strong negative correlation (-0.75) and the Angus a strong positive correlation(+0.84), compared with that of the Bonsmara (+0.39) (Mokolobate, 2015). At this stage it is not clear to what this difference could be attributed to, but one possibility is the small number of records. The genetic correlation between KgC/LSU and DW was not estimable in the Charolais and was -0.75 in the Angus, which is of the same magnitude as the -0.83 of the Bonsmara.
Conclusion
Cow efficiency can be improved dramatically through climate-smart breeding strategies. It is becoming more evident that the inclusion and combination of contributing traits in some kind of a selection index is the more feasible and most appropriate option. Alternative breeding objectives should be developed that would optimize genetic improvement in cow efficiency that include calf weight and cow weight. This could result in the development and evaluation of alternative selection indexes that would facilitate maximum genetic improvement in cow efficiency. Normally, the traits in a selection index are weighed with their economic value. However, the traits could be assigned weights that could be linked to carbon footprints or credits (sequestration) and not only economic weights. It would be possible to link the annual carbon footprint with an LSU, as done by Du Toit et al., 2013).
Breeding strategies and systems to improve the production efficiency of beef cattle could play a significant role in reducing the carbon footprint from beef production. This would enhance climate-smart beef production and support the South African government's commitment to reducing GHG emissions.
Acknowledgements
The financial contributions of Red Meat Research and Development South Africa (RMRD SA) and the National Research Foundation of South Africa (NRF), under grant UID 90099, are acknowledged. The grant holder acknowledges that opinions, findings and conclusions or recommendations expressed in any publication generated by NRF-supported research are those of the authors, and the NRF accepts no liability whatsoever in this regard.
Authors' Contributions
The idea for the study was conceived by MMS, who secured the funding and supervised MCM. MCM was recruited as the student to work on the project and received her MSc. cum laude. FWCN was the supervisor at the university and gave guidance on the study. All authors read and approved the final manuscript.
References
Bosman, D.J., 1994. National Beef Cattle Performance and Progeny Testing Scheme1980-1992. Results. Department of Agriculture and Water Supply, Pretoria, South Africa. [ Links ]
Du Toit, C.J.L., Meissner, H.H. & Van Niekerk, W.A., 2013. Direct greenhouse gas emissions of the game industry in South Africa. S. Afr. J. Anim. Sci. 43, 376-393. [ Links ]
Ferrell, C.L. & Jenkins, T.G., 1982. Efficiency of cows of different size and milk production potential. RLHUSMARC Germ Plasm Evaluation Report 10, 12. [ Links ]
Gilmour, A.R., Cullis, B.R. & Welham, S.J., 2009. ASREML reference manual. NSW Agriculture Biometric Bulletin no. 3. [ Links ]
Greiner, S.P., 2009. Beef cow size, efficiency, and profit. http://pubs.ext.vt.edu/news/livestock/old/aps-200904_Greiner.html [ Links ]
Jordaan, F., 2015. Genetic and environmental trends in landrace beef breeds and the effect on cow productivity. M.Sc. thesis, University of the Free State, Bloemfontein, South Africa. [ Links ]
Koots, K.R., Gibson, J.P., Smith, C. & Wilton, J.W., 1994. Analyses of published genetic parameter estimates for beef production traits. 1. Heritability. Anim. Breed. Abstr. 62, 309-338. [ Links ]
MacNeil, M.D., 2007. Retrospective analysis of selection applied to a ratio. Am. Soc. Anim. Sci. 58, 85-88. [ Links ]
Maddock, T.D. & Lamb, G.C., 2010. Managing beef cow efficiency. University of Florida. IFAS, Florida Coop. Ext. Serv., Animal Science Dept., EDIS Publication AN233. [ Links ]
Meissner, H.H., Scholtz, M.M. & Schönfeldt, H.C., 2012. The status, socio-economic and environmental impact, and challenges of livestock agriculture in South Africa. www.rmrdsa.co.za [ Links ]
Mohammadi, H., Shahrebabak, M.M., Vatankhah, M. & Shahrebabak, H.M., 2013. Direct and maternal (co)variance components, genetic parameters, and annual trends for growth traits of Makooei sheep in Iran. Trop. Anim. Health Prod. 45, 185-191. [ Links ]
Mokolobate, M. C., 2015. Novelty traits to improve cow-calf efficiency in climate smart beef production systems. MSc thesis, University of the Free State, Bloemfontein, South Africa. [ Links ]
Mokolobate, M.C., Theunissen, A., Scholtz, M.M., & Neser, F.W.C., 2014. Sustainable crossbreeding systems of beef cattle in the era of climate change. S. Afr. J. Anim. Sci. 44, S8-S11. [ Links ]
Neser, F.W.C., Konstantinov, K.V. & Erasmus, G.J., 1996. The inclusion of herd-year-season by sire interaction in the estimation of genetic parameters in Bonsmara cattle. S. Afr. J. Anim. Sci. 26, 75-78. [ Links ]
Olivier, W.J., Snyman, M.A., Olivier, J.J., Van Wyk, J.B. & Erasmus, G.J., 2001. Direct and correlated responses to selection for total weight of lamb weaned in Merino sheep. S. Afr. J. Anim. Sci. 31,115-121. [ Links ]
Scholtz M.M., Furstenburg, D., Maiwashe, A., Makgahlela, M.L., Theron, H.E. & Van der Westhuizen, J., 2010. Environmental-genotype responses in livestock to global warming. S. Afr. J. Anim. Sci. 40, 408-413. [ Links ]
Scholtz, M.M., McManus, C., Okeyo, A.M. & Theunissen, A., 2011. Opportunities for beef production in developing countries of the southern hemisphere. Livest. Sci. 142, 195-202. [ Links ]
Theunissen, A., 2011. Characterization of breed additive and heterosis effects in beef cattle using experimental results. M.Sc. dissertation, University of the Free State, Bloemfontein, South Africa. [ Links ]
Thorne, M.S. & Stevenson, M.H., 2007. Stocking rate: The most important tool in the toolbox. College of Tropical Agriculture and Human Resources. University of Hawaii, Manoa. http://www.ctahr.hawaii.edu/oc/freepubs/pdf/PRM-4.pdf. [ Links ]
Van der Westhuizen, R.R., Theron, H.E., Van der Westhuizen, J., Maiwaishe, A. & Jordaan, F., 2010. Newsletter Nat. Beef Record. Impr. Scheme 99: 39. [ Links ]
Van Wyk, J.B., Fair, M.D. & Cloete, S.W.P., 2003. Revised models and parameter estimates for production and reproduction traits in the Elsenburg Dorper sheep stud. J. Anim. Sci. 33, 213-220. [ Links ]
Wall, E., Simm, G. & Moran, D., 2010. Developing breeding schemes to assist mitigation of greenhouse gas emissions. Animal 4, 3 66-376. [ Links ]
Wright, H.B., Pollak, E.J. & Quaas, R.L., 1987. Estimation of variance and covariance components to determine heritabilities and repeatability of weaning weight in American Simmental cattle. J. Anim. Sci. 65, 975-981. [ Links ]
Received 18 October 2017
Accepted 26 March 2019
First published online 3 July 2019
# Corresponding author: MokolobateM@arc.agric.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}