










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 n.2 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i2.14
ARTICLES
Effect of varying levels of dietary inclusion of sweet lupin on the growth production characteristics of ostriches (Struthio camelus var. domesticus)
T.S. BrandI, II, #; J.A. EngelbrechtII; J. van der MerweII; L.C. HoffmanII
IDirectorate: Animal Sciences, Department of Agriculture, Western Cape Government, Private Bag X1, Elsenburg, 7607, South Africa
IIDepartment of Animal Sciences, University of Stellenbosch, Private Bag X1, Matieland, 7602, South Africa
ABSTRACT
Nutrition contributes approximately 75% of the total input costs to an intensive ostrich production unit. An increase in the price of traditional protein sources thus necessitates finding cheaper alternatives. Sweet lupins were identified as a possible alternative; therefore, the effect of feeding various sweet lupin inclusion levels was evaluated throughout the different feeding phases. One hundred and forty ostrich chicks were randomly divided into five dietary treatments with three replications each. The chicks all received a standard commercial pre-starter ostrich diet. Five iso-nutritional diets were formulated for each feeding phase according to specifications set out for each of the different feeding phases using Mixit2+ software. For each feeding phase the diets contained either soybean oilcake meal (control diet, 0LD) or sweet lupins (alternative protein source). The 100 lupin diet (LD) was formulated to include the maximum amount of sweet lupins according to the specifications for the specific species and the feeding phase, and therefore differs between the three feeding phases. Soybean oilcake meal was thus gradually replaced by sweet lupins in the following ratios: 100:0 (0LD), 75:25 (25LD), 50:50 (50LD), 25:75 (75LD), and 0:100 (100LD). Feed and water were supplied ad libitum. Feed intakes and live weights were measured every fortnight until slaughter at ca. 11 months of age. No differences were found between the treatment diets for the live weight, dry matter intake (DMI), average daily gain (ADG) or feed conversion ratio (FCR) at the end of each feeding phase. However, the birds on the 75LD tended to have the highest end weight and ADG, and those on the 100LD the lowest at the end of the starter phase. No differences were found for slaughter weight, dressing percentage and big drum muscle weight of the birds. Birds on the 50LD and 75LD tended to have the heaviest cold carcasses; this differed from the 100LD birds which had the lightest. Birds fed the 50LD had heavier thigh weights than those on the other diets. The results of this study indicate that soybean oilcake meal can be replaced in the diets of slaughter ostriches with sweet lupins up to 15% (75LD) in starter diets and 30% (100LD) in grower and finisher diets without any significant effect on any of the production and slaughter traits.
Keywords: alternative protein, average daily gain, dry matter intake, feed conversion ratio, lupins, ostrich nutrition, production
Introduction
Although the ostrich industry in South Africa is still relatively new, as ostriches have not been domesticated for as long as traditional livestock species (Brand & Olivier, 2011; Cloete et al., 2012), the industry is continuously growing and intensifying since there is still a lot to discover regarding these species. Any increase in reliable scientific information will enhance the scope of opportunities and the profitability of the industry.
On the farm level, high running costs remain a problem, with nutrition contributing ca. 75% of the total input costs of an intensive ostrich production unit. A large part of these costs is represented by the protein source, with soybean oilcake meal being the primary protein source in animal feeds (Dalle Zotte et al., 2013). The increase in the demand for traditional protein sources at a reasonable cost requires of producers to find cheaper alternatives to ensure the cost-efficient production of slaughter ostriches without affecting their growth performance. The nutrient requirements of an ostrich changes as it grows, since in a space of 70 - 80 days, its gastrointestinal tract develops from that of a typical bird neonate to that of a hindgut fermenter (Brand & Gous, 2006). Slaughter ostriches therefore generally require the following diets: pre-starter, starter, grower, finisher and maintenance for the different feeding phases when reared intensively (Brand & Gous, 2006; Brand & Olivier, 2011).
Information on the nutritive value of alternative protein sources in ostrich diets is limited (Cilliers, 1994). This necessitates the quantification of the nutrient composition of raw materials in order to allow for the formulation of diets that fit the needs of the bird more closely and ensure the continued economic viability of the industry. Thus, to improve the feeding regimens of ostriches the inclusion of alternative protein sources such as lupins (Lupinus angustifolius) need to be investigated. Three species of lupin are distinguished according to the colour of the flowers, namely L. angustifolius (blue), L. albus (white) and L. luteus (yellow) (McDonald et al., 2011). There are also sweet (low in alkaloids, < 0.1%) and bitter (alkaloid-rich, 0.1% - 4%) varieties within these species (Breytenbach, 2005). Sweet lupin varieties with low alkaloid content have successfully been cultivated by plant breeders; therefore, the use of these varieties in diets with prolonged intakes should be of little concern regarding toxicity to the animal. However, lupins have not yet been fed to ostriches as part of a balanced feedlot diet. Consequently, the inherent characteristics of lupins, such as the anti-nutritional factors (e.g. alkaloids and non-starch polysaccharides) and high fibre levels that could influence the production and slaughter traits of the birds need to be evaluated to determine the optimum lupin inclusion level per feeding phase.
This study is therefore aimed at determining whether and to what extent soybean oilcake meal can be replaced by locally produced sweet lupin seed in the diets of ostriches (Struthio camelus var. domesticus).
Materials and Methods
Ethical approval (R14/108) for this study was obtained from the Western Cape Department of Agriculture. The trial was carried out from November 2014 until September 2015 at the Kromme Rhee Research Farm near Stellenbosch in the Western Cape Province of South Africa. The farm is situated at longitude 18°50' E and latitude 33°51' S with an altitude of 177 m above sea level.
The ostrich chicks (of both sexes) used in this trial were obtained from the resource flock at the Oudtshoorn Research Farm in the Klein Karoo region of South Africa (situated at longitude 22°15' E and latitude 33°37' S at an altitude of 300 m above sea level) and transported after hatching to the experimental site. On arrival, the chicks were randomly divided into five dietary treatment groups with three replications each, resulting in 15 groups of chicks with 15 - 17 chicks per group. The chicks were initially reared indoors in 15 pens of 2.85 m x 2.05 m per pen with free access to outdoor runs (14.6 m x 1.8 m) during the day. In the late afternoon the indoor pens were closed to protect them from the cold, wind and rain. At nine weeks of age the chicks were moved into outdoor paddocks of approximately 25 m x 20 m to provide sufficient space to avoid unnecessary skin damage and leg abnormalities. The outdoor paddocks were equipped with adequate shelter and feed troughs (67 cm x 60 cm x 78 cm) for the ostriches. The shelter provided, prevented the feed from getting wet from dew or rainfall. Mortalities were recorded and dead birds were sent for post-mortem investigation throughout the experimental period to ensure that the dietary treatments were not responsible for these deaths. Most of the mortalities (37.3%) occurred during the pre-starter phase and were mainly due to fractured legs, tibio-tarsal deformities, haemorrhagic enteritis, yolk sac infection, torsion of the intestine and gastric stasis and dilatation; none was linked specifically to diet.
The trial was initiated at the onset of the starter phase, when the 141 surviving ostrich chicks were 83 days old. Although the chicks were divided into their experimental groups on arrival at the farm, they all received the same standard commercial pre-starter ostrich diet (Table 1). Five iso-nutritional diets were formulated using Mixit2+ software (Agricultural Software Consultants Inc., San Diego, USA) for each feeding phase (i.e., starter, grower and finisher) according to specifications prescribed by a model developed by Gous & Brand (2008). Within each feeding phase these diets contained either soybean oilcake meal (control diet; 0LD) or sweet lupins (Lupinus angustifolius, Eureka cultivar) (alternative protein source; diets 25LD, 50LD, 75LD and 100LD) as the primary protein source.
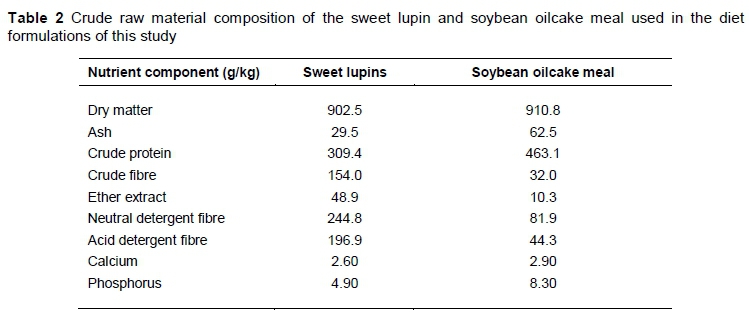
Table 2 shows the crude raw material composition of the sweet lupin and soybean oilcake meal used in the diet formulations of this study. These values were determined by grinding the raw material samples using a RetschTM ZM200 sample mill (Haan, Germany) with a 1.5 mm screen to create a meal with a consistent particle size. Thereafter, the raw materials were analyzed using the methods of the Association of Official Analytical Chemists (AOAC, 2002) for dry matter (DM) (method 934.01), ash (method 942.05), crude protein (CP) (method 976.05), crude fibre (CF) (method 962.09), ether extract (EE) (method 920.39), acid detergent fibre (ADF) (Goering & Van Soest, 1970), neutral detergent fibre (NDF) (Robertson & Van Soest, 1981). The calcium (Ca) and phosphorus (P) values were analyzed using method 6.1.1 (dry ashing) of the Agri Laboratory Association of Southern Africa guidelines (ALASA) (ALASA, 1998).
The 100-lupin diet (LD) was formulated to include the maximum amount of sweet lupins according to the specifications for the specific species and the feeding phase. The maximum number of lupins included in the 100LD therefore differs between the three feeding phases. The remaining three diets were formulated by mixing the diets to determine the gradual increase in lupins in the diets from 0LD up to 100LD.
Soybean oilcake meal was thus gradually replaced by sweet lupins (Lupinus angustifolius, Eureka cultivar) at the following ratios: 100:0 (0LD), 75:25 (25LD), 50:50 (50LD), 25:75 (75LD), and 0:100 (100LD) to make up the five dietary treatments for each feeding phase (Tables 3 - 5). The ostriches in this trial were fed their respective diets ad libitum throughout the different feeding phases and according to standard practices. They also had free access to clean, fresh water throughout the trial period. In the results and discussion sections, diets 1 - 5 will be referred to as 0LD, 25LD, 50LD, 75LD and 100LD for ease of identification. LD represents lupin diet and the number indicates the ratio of the lupin inclusion level to the amount of soybean oilcake meal in the diet. The results obtained for each of the three feeding phases were statistically analyzed separately as the lupin inclusion levels in the diets differed between the phases.
The total alkaloid content of the finely ground pooled feed samples of the diets containing sweet lupins (Lupinus angustifolius, Eureka cultivar) was determined by the method described by Boschin et al. (2008), with minor modifications. The sample preparation method was modified to extract the total alkaloid content directly by using a 50:50 methylene dichloride:methanol (MDC:MeOH) mixture. The total alkaloid content was then analyzed using GC-MS analysis with a 30 m x 0.25 mm, internal diameter 0.25 μηι, AT-Wax capillary column. The following temperature settings were used for the gas chromatography: 150 °C for five minutes, which was then increased by 5 °C per minute up to 300 °C for 15 minutes. Analysis was performed in split mode, with a split ratio of 1 : 25. The injection volume was 1 μΙ_, injection temperature 250 °C, interface temperature 300 °C, and the acquisition was from m/z 50 to 450. The source operated in EI mode at eV. The total alkaloid content was determined using Mass library (Agilent) and the detection limit used to quantify the total alkaloids was 100 ng/mL. However, no alkaloids were found in the respective feed samples at this detection limit.
The spectrophotometry method described by Von Baer et al. (1978) was used to determine the total alkaloid content of the sweet lupin cultivar used by Brand & Smith (2016). This method is a quantitative determination of total alkaloids with bromocresol purple at 405 nm. The total alkaloid content (49.1 mg/kg) of the sweet lupin cultivar found in the study by Brand & Smith (2016) was therefore used to calculate the estimated amount of total alkaloids of the five dietary treatments of the current study (Tables 3 - 5).
Fortnightly feed intake per paddock was measured by weighing back the refusals for the two weeks and subtracting this value from the quantity of feed offered during that period. The weight of each bird was also recorded every second week on the same day for the duration of the trial for the determination of the average daily gain (ADG). The feed conversion ratios (FCR) per treatment diet were expressed as DMI per kg gain per feeding phase. The birds received their compulsory inoculation against Newcastle disease at approximately five months of age, and their feathers were not clipped or plucked at six months of age as these were still blood feathers.
The birds were slaughtered at approximately 11 months of age at a registered abattoir, Mosstrich (situated at longitude 22°00' E and latitude 34°09' S) in Mossel Bay. They were moved to quarantine paddocks (tick-free environment, rodent control, no vegetation and a 3 m cleared area around each paddock) 14 days prior to slaughter as required by the European Union (EU) meat quality standards highlighted in the report by the Department of Agriculture, Forestry and Fisheries (Maja, 2012; DAFF, 2014). Transport of the birds to the abattoir was performed by private contractors and on arrival the ostriches were offloaded at the specifically designed lairage area. The birds were kept in lairage for about 16 hours before slaughter and received fresh clean water during lairage.
The slaughter weight was determined by weighing each bird on an electronic walk-on scale before they entered the stunning area. The slaughtering procedures used by the abattoir staff were similar to those described by Hoffman (2012). After exsanguination, the feathers of each bird were plucked by hand and the skin was carefully flayed from the carcass. Following evisceration, the abdominal fat, better known as the fat pad, was removed, collected, and weighed for each bird to determine whether the increasing sweet lupin inclusion levels of the five dietary treatments had any effect on the yield.
The carcasses, consisting of the neck, wings, chest and thighs were washed and moved into the cold storage facilities (0 - 2 °C). The carcasses were left in the cold-storage facilities for 18 hours and before deboning commenced the next morning the cold carcass weights were recorded. These weights were used to calculate the dressing percentage. The right thigh weight of each ostrich was also recorded and used to calculate the contribution of the thigh weight to the carcass. The weight of the right thigh was multiplied by two and divided by the weight of the cold carcass to calculate the percentage contribution. The weight of the big drum muscle (muscularis gastrocnemius) of the right thigh of each ostrich was also recorded and divided by the weight of the right thigh to give an indication of its contribution to the total thigh weight.
Analysis of variance (ANOVA) was done using the GLM (General linear model) procedure of SAS statistical software (Version 9.2, SAS Institute Inc., Cary, NC, USA). An experimental unit consisted of a group of ostriches in a paddock fed a specific diet and thus paddocks were random replicates for treatment diets. The Shapiro-Wilk test was performed on all data to test for deviation from normality (Shapiro & Wilk, 1965). Fisher's protected least significant differences were calculated at the 5% level to compare treatment means (Ott, 1998).
To illustrate and describe the growth patterns over the entire trial period for each treatment diet Gompertz growth models were fitted to the data and the slopes compared to assess any differences in weight gain using analysis of variance (ANOVA):
Bird weight (BW) = a*exp(-exp(-b*(age-c)))
where: a = mature weight (kg),
b = rate of maturing parameter, and
c = age of maximum growth (days) (Brand & Olivier, 2011).
As the trial only commenced at the starter phase (± 80 days of age) and the ostriches were slaughtered before mature weight equilibrium was reached, estimated Gompertz regression parameters were not of practical interpretation value and were thus not compared statistically to illustrate and describe the growth patterns.
For the ANOVA and regression models on live weight and DMI, fitted over the whole trial period, the weights of birds at the start of the trial were used as a covariate. To determine the effect of the treatment diet within each phase, the end weight of the previous phase was used as a covariate in the ANOVA and regression models, i.e., initial weight, end weight and DMI within each phase were adjusted for weight differences at the beginning of that phase.
The DMI per bird per day per paddock was calculated for the entire growth period, as well as per phase, as the total feed intake per paddock for a given period divided by the "bird x days." The number of ostriches per paddock did not necessarily remain stable over a given period as a result of mortalities. To account for the actual number of ostriches that consumed the feed during a given period, bird x days were calculated per paddock as the sum of the product of the number of birds and days over observation times. The ADG per paddock was determined from the slope of the linear regression of growth on age (in days) within each phase, as well as for the entire period. The FCR was calculated by dividing the DMI by the ADG. Completely random ANOVAs were conducted on the production traits, ADG and FCR.
Supplementary to the ANOVAs contrasts were specified to determine which (if any) polynomial trend over protein levels in the diet was significant. Production traits were further analyzed by fitting the relevant regression functions to the data to describe the observed trends. A probability level of 5% was considered significant and these polynomials functions were reported. When trends were not significant, linear polynomials were used in graphs to elucidate results.
One-way ANOVAs was also performed to analyze the slaughter traits according to the experimental design where each treatment diet was completely randomly repeated in three paddocks.
Results
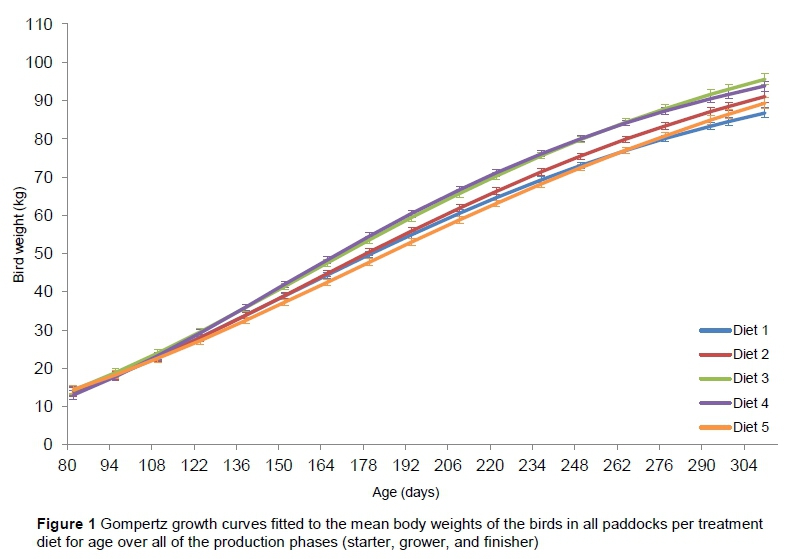
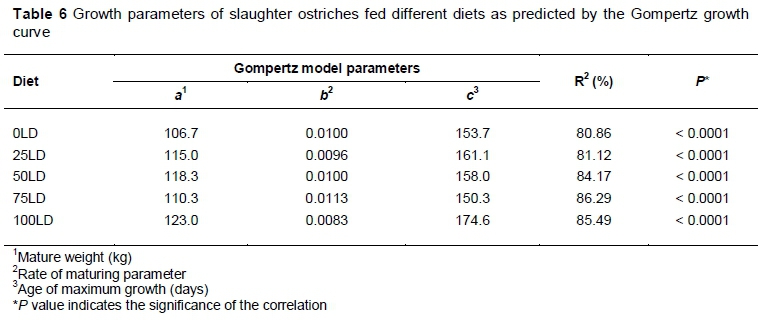
Gompertz growth curves for weight change in ostriches on each diet over the experimental period are presented in Figure 1. The Gompertz equation fits the data well within the boundaries of the data (starter, grower and finisher phases) and reveals the sigmoidal growth curve for all the diets. The estimated values for the growth parameters of the Gompertz model for the different diets are presented in Table 6. There were no differences for the production trait means between the different diets.
The average initial body weight of the birds at 83 days of age was 13.78 ± 0.55 kg (LSM ± SE) and no differences were observed between treatment groups in the initial weight of the birds for each feeding phase. The mean weight of the birds at the end of the entire experimental period (all phases) was 91.7 ± 1.52 kg and this did not differ (P = 0.34) between the respective dietary treatments. Furthermore, no differences were found between the treatment diets for the weights of the birds at the end of each feeding phase. However, there was a tendency (P = 0.07) for the live weights at the end of the starter phase to differ between the treatments. The mean end weight of the birds on the 75LD tended to be 26.2% heavier than that of the birds fed the 100LD, which were the lightest (35.9 ± 1.75 kg) at the end of the starter phase. Regarding the production traits for the starter, grower, finisher and overall growth phases no differences were found between the treatment diets for the DMI, ADG or FCR. The only exception to this was the tendency (P = 0.09) observed in the ADG during the starter phase. The growth rate of the birds on the 75LD was 33.0% greater than that of the birds on the 100LD, which had the lowest gain during this period. Mean DMI and ADG of 2020.5 ± 61.9 g/bird/day and 351.5 ± 8.3 g/bird/day respectively were observed over all the treatments, with a mean FCR of 5.75 ± 0.09 kg feed/kg weight gain found for the whole trial period (all phases).
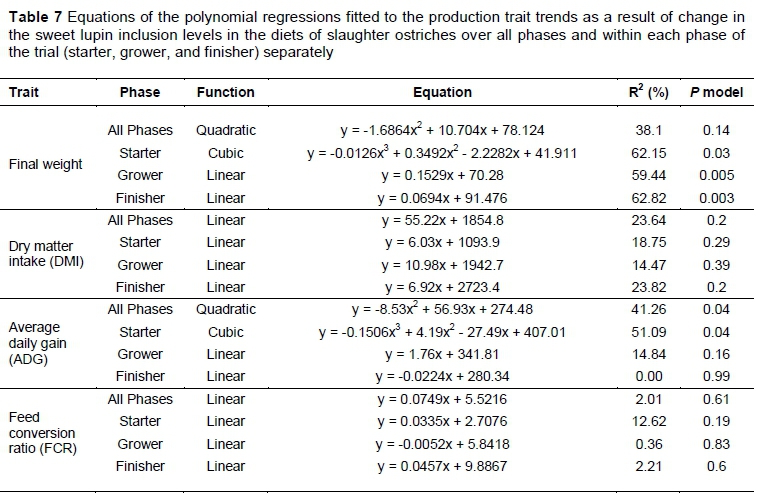
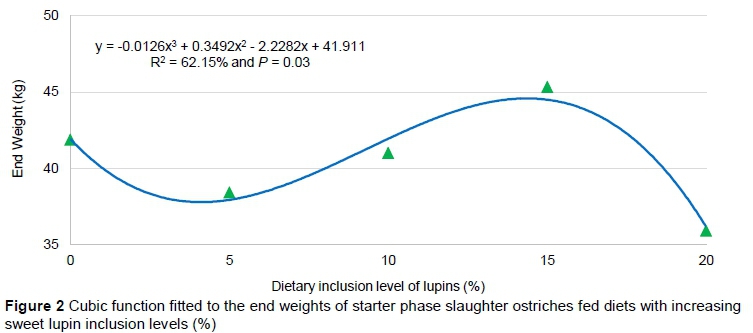
Production traits were further analyzed using various regression models to describe trends resulting from changes in the sweet lupin inclusion levels in the diets over all phases and within each phase of the trial (starter, grower, and finisher) separately. A probability level of 5% was considered significant and the regression models corresponding to the most significant, polynomial contrasts are presented in Table 7. Where none of the polynomials was significant, the linear polynomial was used to indicate that there was no trend and is also presented in Table 7. The only significant trends (P <0.05) in the regression models were for end weights in the starter, grower and finisher phases. A cubic function (R2 = 62.2% and P = 0.03) best fitted the end weights of the starter phase (Figure 2), whereas for the grower (R2 = 59.4% and P = 0.005; Figure 3) and finisher (R2 = 62.8% and P = 0.003; Figure 4) phases, a linear function fitted the end weight over diet relationship.
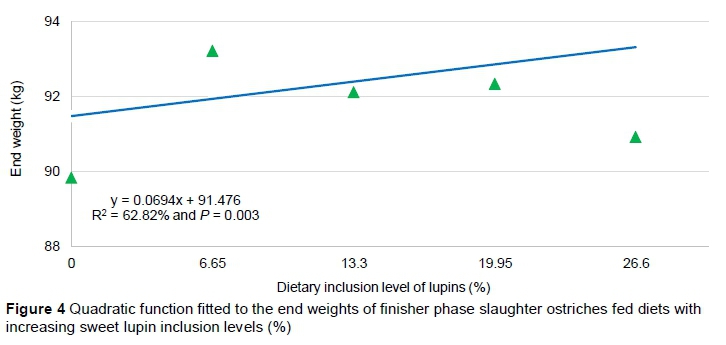
Regression analysis of the remaining three production traits (DMI, ADG and FCR) per phase and over the entire experimental period revealed no significant trends. However, the regression model for the ADG over the whole growth period and during the starter phase was significant (P <0.05). Regression analysis of data from the whole period indicated that the ADG showed a quadratic relationship (R = 41.3% and P = 0.04) between the diets, with a tendency for a higher growth rate with the intermediate diets combining soybean oilcake and sweet lupins (Figure 5). Regression analysis of the ADG during the starter phase revealed that a cubic function fitted the data best (R2 = 51.1% and P = 0.04), with 75LD having the highest ADG and 100LD the lowest.
It must be noted that although some of the regression models are significant (P <0.05), the R2 values of the regression models presented in Table 7 give an indication of how close the data lie to the fitted regression line. As the R2 values were not very high, the regression models did not explain a high percentage of the variation in the data. The highest R2 value found was 62.8%. Thus, the regression functions do not describe the effect that the treatments have on the production traits very accurately.
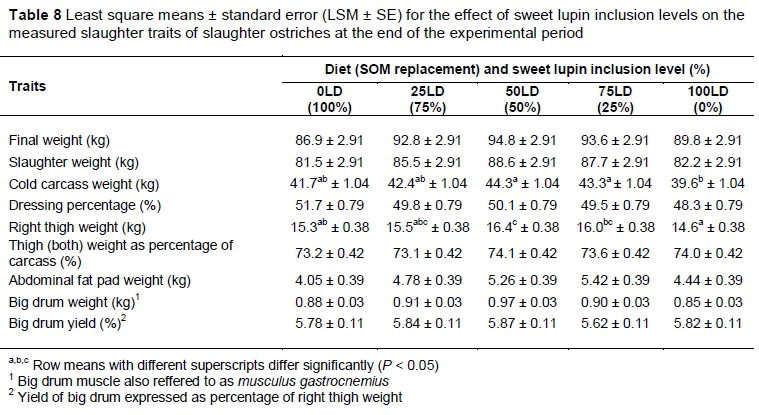
No differences were found between the treatment diets in terms of the final weights (P = 0.36) and the slaughter weights of the ostriches at the abattoir (P = 0.32) (Table 8). The cold carcass weights differed (P = 0.08) between the treatment diets. The birds that were fed on the 50LD and 75LD did not differ and had the heaviest carcasses of 44.3 ± 1.04 kg. Birds fed on the 0LD and 25LD diets had lighter carcasses (41.7 ± 1.04 kg and 42.4 ± 1.04 kg, respectively) and did not differ from the cold carcass weights of the group that were fed on the 100LD diet (39.6 ± 1.04 kg). However, the cold carcass weights of birds fed the 50LD and 75LD differed (P <0.05) from those fed the 100LD, weighing 10.6% heavier. With regards to the dressing percentage, no differences (P >0.05) were observed between the treatments (Table 8). Some variation (P <0.05) was, however, found in the right thigh weights, with the thighs of birds receiving the 50LD weighing 12.3% more (P <0.05) than that of those receiving the 100LD. The right thigh weights of the birds on the 0LD (15.3 ± 0.38 kg), 25LD (15.5 ± 0.38 kg) and 75LD (16.0 ± 0.38 kg) had intermediate values. Furthermore, no differences were found in the combined thigh weights as a percentage of the carcass (P = 0.37), the weights of the abdominal fat pads (P = 0.15) or the big drum muscles (P = 0.16), or for the big drum muscles when expressed as a percentage of the right thigh (P = 0.59).
Discussion
According to Dalle Zotte et al. (2013), all of the nutrient requirements of an animal must be met in order to successfully rear any livestock species. Up until 1995, ostrich diet formulations were largely based on energy values for poultry ingredients, as information on metabolizable energy (ME) values derived for ostriches was scarce (Angel, 1996). However, computer-based mathematical simulation models have since been developed by Gous & Brand (2008), which is a useful tool in determining least-cost diet formulations that can help feed manufacturers improve the competitiveness of the ostrich industry. These models have been updated to take into account the effects of different energy and protein levels on ostrich production (Carstens, 2013).
Lupins are cheaper than other protein sources, making them widely used in livestock feeds (Edwards & Van Barneveld, 1998). Only soybeans, which are a more popular protein source, can compete with lupins in this respect. However, lupins have a fibrous seed coat which contributes to their higher NDF content (201.1% more NDF than soybean oilcake meal) and affects the digestibility of the meal (McDonald et al., 2011). This is a major concern for young monogastric animals (Ensminger, 1992), but the ostrich- a hindgut fermenter-has the ability to digest fibrous material (Brand & Olivier, 2011). This allows ostriches to utilize a greater proportion of the available energy in the feed than poultry and pigs (Brand et al., 2000c; Kruger, 2007). Swart (1988) found that ostriches could possibly absorb between 12% - 76% of their energy in the form of volatile fatty acids, which are the end products of fibre digestion in the large intestine. It is, however, important to remember that ostrich chicks are not able to utilise fibre efficiently before approximately three months of age, as their ability to digest crude fibre only develops thereafter (Angel, 1996). This gives rise to the need for different feeding phases at specific growth stages when ostriches are reared intensively. Swart (1988) also described the ability of the ostrich to utilize low quality raw materials, showing that they can effectively digest up to 66% of the hemicellulose and 38% of the cellulose in a feed.
Currently, lupin seed meal serves as an economically viable alternative to soybean meal and rapeseeds thanks to the low-alkaloid sweet varieties that are available (Ewing, 1997). As mentioned, lupins contain alkaloids that can be toxic to the animal. However, the seeds of L. angustifolius cultivars grown for their low levels of alkaloids maintain a low and relatively stable alkaloid content (Brand & Brandt, 2000). In order to prevent the risk of toxicity only alkaloid levels lower than 0.06% are deemed suitable for animal feeds (McDonald et al., 2011). In this study, no alkaloids were found in the respective feed samples as analyzed using the method described by Boschin et al. (2008) at a detection limit of 100 ng/mL. The estimated total alkaloid content per diet, calculated from the total alkaloid content of the sweet lupin cultivar found by Brand & Smith (2016), was also below 0.06%. The use of the provided diet formulations (Tables 4 -6) with prolonged intakes should therefore not present any risk of toxicity for the birds and can therefore be regarded as safe. They should also not influence the feed preference of the birds, which can be a problem with high alkaloid levels as they decrease the palatability of a feed (Smith, 2005).
Long-term feed intake is primarily influenced by the energy content of the diet, as an animal will eat in order to meet its energy requirements (Forbes & Shariatmadari, 1994). Throughout the trial period, the experimental diets were formulated to meet the nutrient requirements of the birds and had equal ME values; any differences found in the nutrient components were deemed too small to have had a significant effect on the DMI of the birds. Lupins contain a relatively high protein content (32%), although they have limited biological value due to their low methionine and proportionally low lysine content in comparison with other legumes (Ewing, 1997; Edwards & Van Barneveld, 1998). These shortfalls can be supplemented by including other protein sources in the diet or by using synthetic amino acids. Synthetic methionine and lysine were thus included in the respective trial diets where necessary.
The Gompertz equations fitted to the mean weights of the birds per paddock for each diet, over the course of the experimental period, revealed sigmoidal growth curves for all the diets. As mentioned, the growth stage of the bird will determine its nutrient requirements, because as the bird grows, its body composition in terms of the protein : fat ratio changes (Brand & Olivier, 2011). There were in fact no differences in the general growth patterns of the birds (three different phases: initial self-accelerating, intermediate linear and self-decelerating) between the five dietary treatments. It can therefore be concluded that the composition of the diets do not impede the growth of the ostriches.
Brand & Gous (2006) stated that the nutrient composition of the feed will determine the production rate of growing ostriches. In terms of the production traits (end weight, DMI, ADG and FCR) of these slaughter ostriches, results were within the bounds determined by previous intensive slaughter ostrich production studies (Brand et al., 2000a; 2004b; 2014; Dalle Zotte et al., 2013). This indicates that the production traits for all the diets resembled that of the production norms for slaughter ostriches.
In this study, a tendency (P = 0.09) was found for the ADG of the starter phase to vary between the diets, with the birds fed the 75LD tending to have a higher ADG than the birds fed the 100LD, although no differences were observed for the DMI and FCR. The DMI values were similar to those of Brand et al. (2000b) who evaluated the effects of three different crude fibre levels on the production performance of slaughter ostriches. In contrast, Brand et al. (2000b) found differences in the ADG and FCR. The greater number of treatment diets used in the sweet lupin trial compared to Brand et al. (2000b), as well as the knowledge gained over the years in terms of understanding the nutritional requirements of the ostrich since the work by Brand et al. (2000b), could well be responsible for the difference in the trends. However, Brand et al. (2014) found differences in all three production traits when evaluating low, medium and high energy diets.
During the grower phase of the sweet lupin trial, no differences were found for the DMI, ADG or FCR. These results are supported by Brand et al. (2004b) who also found no differences in the production traits of ostriches when feeding different protein levels. However, studies on the effects of different dietary energy levels by Brand et al. (2004b) showed differences in each trait between the different treatment diets. Brand et al. (2014) found no differences in the DMI of the birds, which supports the results found in the sweet lupin trial, but the results are inconsistent in terms of the ADG and FCR, where differences were observed. However, Brand et al. (2000b) found no differences in the FCR of the birds, but differences were observed for the DMI and ADG (about 1569.0 and 358.3 g/bird/day, respectively). Relative to the sweet lupin trial, Brand et al. (2000b) observed lower intakes but still achieved similar growth rates and thus better FCRs, ranging between 3.80 - 5.08 kg feed/kg weight gain.
Regarding the finisher phase of the sweet lupin trial, once again no differences were observed between the treatment diets. The same trend was followed for the production traits of ostriches in the finisher phase of Brand et al. (2000b) as for the grower phase. The DMI and ADG revealed differences between the three treatment diets, but no differences were observed for the FCR, supporting the results found in the sweet lupin trial. The results of the protein trial by Brand et al. (2004b) partly support the findings of the sweet lupin trial, with no differences observed in the ADG or FCR of the birds. However, the total DMI of the birds was higher than that found in the current trial (3306.0 versus 2815.1 g/bird/day). This was unexpected as the birds in the sweet lupin trial were slaughtered at 11 months of age and not 12 as in the protein trial of Brand et al. (2004b).
Brand et al. (2000a) found no differences in the DMI, ADG or FCR when feeding ostriches diets differing in protein content (13%, 15% and 17% protein) from four to 11 months of age. These findings support the findings of the sweet lupin trial, where no differences were observed in the production traits with regard to the whole trial period. Brand et al. (2000a) also fed different energy levels (9, 10.5 and 11 MJ/kg ME), which also did not result in any differences in the ADG (approximately 302.3 g/bird/day) of the birds, but differences were observed for the DMI (2647 g/bird/day) and FCR (7.68 kg fee/kg weight gain) (Brand et al., 2000a). Contradictory to these findings, Dalle Zotte et al. (2013) found differences in the ADG of ostriches fed 0%, 3%, 6%, 9% and 12% dietary levels of cottonseed oilcake meal, but no differences for the DMI and FCR. However, guidelines for feeding and managing ostriches between six and 13 months of age recommend a DMI ranging from 1.1 and 2.9 kg/bird/day, an ADG of 200 - 340 g/bird/day and a FCR of between 5 and 15 kg feed/kg weight gain (Brand & Olivier, 2011). The results found in the respective studies are thus in agreement with the guidelines provided by Brand (2014). However, it is important to remember that as the bird ages, the ADG decreases after the inflection point is reached (Brand & Gous, 2006), a trend also observed in these studies.
Regarding the slaughter traits of the birds, the dietary treatments did not influence growth or production. This can somehow be expected since the ostrich has the ability to digest fibrous material and so the higher dietary sweet lupin inclusion could also be utilised. The values obtained in the sweet lupin trial resembled the findings of Brand et al. (2004a), Hoffman et al. (2007) and Viviers (2015) who also evaluated the slaughter traits of ostriches in growth studies. The birds in the sweet lupin trial were slaughtered at 11 months of age rather than 12 - 14 months, as was done in the cited studies. The birds in this study therefore attained a lower mean final live weight. The cold carcass weight and right thigh weight in the current study followed the same trend as the mean final weight (Kritzinger, 2011), with the birds slaughtered at an early age (11 months) having a lighter cold carcass and consequently lighter right thighs. Therefore, the big drum muscles of the birds in the sweet lupin trial were also lighter than those found by Viviers (2015). However, the mean dressing percentage (49.9%) fell within the range reported for previous studies, with 45.1% for the energy trial by Viviers (2015) and 51.1% for Hoffman et al. (2007). The yield of both thighs expressed as a percentage of the carcass was similar to that reported by Viviers (2015).
Brand et al. (2004a) found a mean abdominal fat pad weight of about 6.4 kg, while Viviers (2015) reported a lower value of 5.4 kg. However, in both aforementioned studies the birds were slaughtered at 12 months of age, so the lower fat pad weight (mean value of 4.8 kg) of the birds in the sweet lupin trial is to be expected. Differences in the abdominal fat pad weight were reported in the energy trial by Brand et al. (2004a), but Viviers (2015) and Brand et al. (2004a) (protein trial) did not observe any differences, which is in agreement with the results of this study. Hoffman (2005) postulated that any extra energy in the diet is stored in the abdominal fat pad and not in the intra-muscular fat reserves.
Overall it can be concluded that sweet lupins can successfully be included in the diets of slaughter ostriches up to 15% (75LD) in the starter phase and 30% (100LD) in the grower and finisher phases. Similar results were achieved for the production and slaughter traits measured as reported for previously performed studies, suggesting that the sweet lupin inclusion in the diets did not have any negative impact on the birds. The fact that, overall, there were no differences in the production traits or measured slaughter traits between the treatments during the different feeding phases is satisfactory. However, some variation was found in the end weight and ADG of the starter phase and for the cold carcass and right thigh weights. Nonetheless, it appears that sweet lupins can replace soybean oilcake meal in the diets of slaughter ostriches up to the dietary sweet lupin inclusion levels evaluated in this study without any detrimental effect on any of the production and slaughter traits, as in this study the birds showed acceptable growth performance.
When these findings are compared to results obtained and recommendations made by other researchers, it is clear that the ostrich is able to consume and utilise this raw material at higher inclusion levels than other monogastric animals. In "The Feeds Directory" by Ewing (1997), the recommended inclusion rates of lupin flakes/meal per species are as follows: 0% in creep, weaner, and grower diets of pigs, but for finisher and sow diets, a 7.5% inclusion level is allowed; in poultry diets 0% inclusion is advisable in chick diets, 5% in broiler diets and 7.5% in breeder and layer diets.
According to McDonald et al. (2011), the maximum lupin inclusion levels in the diets of adult poultry and pigs is 10% and for growing pigs and broilers, 5%. Brand et al. (1995) noted that lupins can be included up to 20% in the diets of growing-finishing pigs as a replacement for fishmeal. However, a lower intake and growth rate can be expected due to the higher fibre content of lupins. Kim et al. (2012) reported that the inclusion level of lupins should be limited to 5% - 10% in commercial piglet weaner diets as lupins consist of approximately 25% seed coat (mostly insoluble fibre), while its kernel contains about 30% cell wall materials (polygalacturonans), which cause a dilution effect on the nutritional density of the balanced feed. However, there is a general perception that lupins can be included at levels higher than 15% in the diet of weaner pigs, provided that the indigestible hull is removed.
Whole lupins can thus be included in the diets of slaughter ostriches at higher inclusion levels than for poultry and pigs, without any concern regarding the digestibility of the feedstuff and treatment prior to feeding. This can be attributed to the hindgut fermentation ability of the ostrich which enables them to utilise fibrous materials more efficiently. This is also confirmed by Brand et al. (2000a), who found that ostriches can perform well on high fibre diets, without effecting their production negatively.
Conclusion
From this study it is concluded that sweet lupins can be included in the diets of slaughter ostriches up to 15% (75LD) in starter diets and 30% (100LD) in grower and finisher diets to replace soybean oilcake meal, without any significant detrimental effect on the production and slaughter traits. The hindgut fermentation ability of ostriches most likely enables them to utilise sweet lupins very effectively, despite their higher fibre content than soybean oilcake meal. These results will contribute to the limited knowledge on ostrich nutrition and will be incorporated in the mathematical optimisation model developed by Gous & Brand (2008) to predict feed intake and other production traits more accurately. These findings may also assist in creating a potential market for locally produced protein sources such as lupins and broaden our knowledge with regard to the potential of this raw material as a feed ingredient for animals.
Acknowledgements
Acknowledgements are hereby made to the Western Cape Department of Agriculture for joint funding of the study, as well as the personnel in their employment for their aid. In addition, the Western Cape Agricultural Research Trust for the joint funding of the study. Finally, to Mosstrich abattoir in Mossel Bay for their willingness to accommodate our needs during the slaughter process and subsequent data collection. This manuscript is summarized from the Master's thesis of Jacomina Aletta Engelbrecht.
Authors' Contributions
Concept and design: TSB; data collection and analysis: JAE; drafting of paper: JVDM; critical revision and final approval of version to be published: LCH.
Conflict of Interest Declaration
The authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest (such as honoraria, educational grants, participation in speakers' bureaus, membership, employment, consultancies, stock ownership, or other equity interest and expert testimony or patent-licensing arrangements), or non-financial interest (such as personal or professional relationships, affiliations, knowledge, or beliefs) in the subject matter or materials discussed in this manuscript.
References
ALASA., 1998. Method 6.1.1 - Dry ashing. In: Handbook of Feeds and Plant Analysis. Ed: Palic, D., Hatfield, Pretoria, South Africa. [ Links ]
Angel, C.R., 1996. A review of ratite nutrition. Anim. Feed Sci. Technol. 60, 241-246. [ Links ]
AOAC., 2002. Official Methods of Analysis (17th ed.). Association of Official Analytical Chemists, Inc., Arlington, Virginia, USA. [ Links ]
Boschin, G., Annicchiarico, P., Resta, D., D'Agostina, A. & Arnoldi, A., 2008. Quinolizidine alkaloids in seeds of lupin genotypes of different origins. J. Agric. Food Chem. 56, 3657-3663. [ Links ]
Brand, T.S. & Brandt, D.A., 2000. Alkaloid content of South African lupins (L luteus, L albus and L angustifolius species) and determination thereof by Near Infra-red Reflectance Spectroscopy. S. Afr. J. Anim. Sci., 30, 11-12 (Supplement 1). [ Links ]
Brand, T.S. & Gous, R.M., 2006. Feeding Ostriches. In: Feeding in Domestic Vertebrates: From a structure behaviour. Ed: Bels, V., Wallingford, England. CAB International. pp. 136-155. [ Links ]
Brand, T.S. & Olivier, A., 2011. Ostrich Nutrition and Welfare. In: The Welfare of Farmed Ratites. Eds: Glatz, P., Lunam, C. & Malecki, I., Heidelberg, Germany. Springer. pp. 91-109. [ Links ]
Brand, T.S. & Smith, N., 2016. Differences in the various anti-nutritional factors and the amino acid and digestible energy content (pigs) of different legumes produced in South Africa. S. Afr. J. Anim. Sci. (Submitted). [ Links ]
Brand, T.S., Olckers, R.C. & Van der Mewe, J.P., 1995. Evaluation of faba beans (Vicia faba cv. Fiord) and sweet lupins (Zupizus albus cv. Kiev) as protein sources for growing pigs. S. Afr. J. Anim. Sci. 25, 31-35. [ Links ]
Brand, T.S., Nell, C.J. & Van Schalkwyk, S.J., 2000a. The effect of dietary energy and protein level on the production of growing ostriches. S. Afr. J. Anim. Sci. 30, 15-16 (Supplement 1). [ Links ]
Brand, T.S., Brand, Z., Nel, K., Van Schalkwyk, K. & Salih, M., 2000b. Latest ostrich nutrition research leads to cheaper diets and lower production costs. Elsenburg J. pp. 8-17 (in Afrikaans). [ Links ]
Brand, T.S., Van der Merwe, J.P., Salih, M. & Brand, Z., 2000c. Comparison of estimates of feed energy obtained from ostriches with estimates obtained from pigs, poultry and ruminants. S. Afr. J. Anim. Sci. 30, 13-14 (Suppl. 1). [ Links ]
Brand, T.S., Gous, R.M., Horbanczuk, J.O., Kruger, A.C.M., Aucamp, B.B. & Brand, Z., 2004a. The effect of dietary energy and protein (amino acid) concentrations on the end products of slaughter ostriches. S. Afr. J. Anim. Sci. 34, 107-109 (Suppl. 2). [ Links ]
Brand, T.S., Gous, R.M., Kruger, A.C.M., Aucamp, B.B., Nel, C.J. & Horbanczuk, J.O., 2004b. The effect of dietary energy and protein (amino acid) concentration on the performance of slaughter ostriches. S. Afr. J. Anim. Sci. 34, 104-106 (Supplement 2). [ Links ]
Brand, T.S., Carstens, P.D. & Hoffman, L.C., 2014. The effect of dietary energy concentrations on production variables of ostrich chicks (Struthio camelus var. domesticus). Br. Poult. Sci. 55, 610-618. [ Links ]
Breytenbach, L., 2005. The influence of processing of lupins and canola on apparent metabolizable energy and broiler performance. MSc (Agric) thesis, University of Stellenbosch, South Africa. [ Links ]
Carstens, P.D., 2013. Studies to develop a mathematical optimisation model to describe the effect of nutrition on the growth of ostriches (Struthio camelus var. domesticus). MSc (Agric) thesis, University of Stellenbosch, South Africa. [ Links ]
Cilliers, S.C., 1994. Evaluation of feedstuffs and the metabolisable energy and amino acid requirements for maintenance and growth in ostriches (Struthio camelus). PhD thesis, University of Stellenbosch, South Africa. [ Links ]
Cloete, S.W.P., Brand, T.S., Hoffman, L., Brand, Z., Engelbrecht, A., Bonato, M., Glatz, P.C. & Malecki, I.A., 2012. The development of ratite production through continued research. World Poult. Sci. J. 68, 323-334. [ Links ]
DAFF, 2014. A profile of the South African ostrich market value chain. Directorate: Marketing of the Department of Agriculture, Forestry and Fisheries. [Internet document] Available from: URL: http://www.nda.agric.za/doaDev/sideMenu/Marketing/AnnualPublications/CommodityProfiles/Livestock/Ostrich market value chain profile2014.pdf, pp. 1-41 (Accessed 19 January 2016). [ Links ]
Dalle Zotte, A., Brand, T.S., Hoffman, L.C., Schoon, K., Cullere, M. & Swart, R., 2013. Effect of cottonseed oilcake inclusion on ostrich growth performance and meat chemical composition. Meat Sci. 93, 194-200. [ Links ]
Edwards, R.A. & Van Barneveld, R.J., 1998. Lupins for Livestock and fish. In: Lupins as crop plants: biology, production and utilization. Eds: Gladstones, J.S., Atkins, C. & Hamblin, J., Cambridge, CAB International, University Press. pp. 385-409. [ Links ]
Ensminger, M.E., 1992. Poultry Science. Danville, Illinois. Interstate Publishers. pp. 28-32. [ Links ]
Ewing, I., 1997. Lupin Flakes/Meal. In: The Feeds Directory, Vol. 1. Commodity Products. Leicestershire, England. Contex Publications. pp. 54. [ Links ]
Forbes, J.M. & Shariatmadari, F., 1994. Diet selection for protein by poultry. World Poult. Sci. J. 50, 7-24. [ Links ]
Goering, H.K. & Van Soest, P.J., 1970. Forage Fibre Analyses (Apparatus, reagents, procedures and some applications). In: Agricultural Handbook No. 379. USDA-ARS, U.S. Department of Agriculture Washington. [ Links ]
Gous, R.M. & Brand, T.S., 2008. Simulation models used for determining food intake and growth of ostriches: an overview. Aust. J. Exp. Agric. 48, 1266-1269. [ Links ]
Hoffman, L.C., 2005. A review of the research conducted on ostrich meat. Proceedings of the 3rd International Ratite Science Symposium of the World's Poultry Science Association (WPSA) and 12th World Ostrich Congress, Madrid, Spain. pp. 107-119. [ Links ]
Hoffman, L.C., 2012. Advances in the electrical stunning and bleeding of ostriches. Anim. Welfare. 21, 9-13 (Supplement 2). [ Links ]
Hoffman, L.C., Brand, M.M., Muller, M. & Cloete, S.W.P., 2007. Carcass and muscle yields of ostriches as influenced by genotype. S. Afr. J. Anim. Sci. 37, 256-260. [ Links ]
Kim, J.C., Heo, J.M., Mullan, B.P. & Pluske, J.R., 2012. Performance and intestinal responses to dehulling and inclusion level of Australian sweet lupins (Lupinus angustifolius L.) in diets for weaner pigs. Anim. Feed Sci. Technol. 172, 201-209. [ Links ]
Kritzinger, W.J., 2011. Allometric description of ostrich (Struthio camelus var. domesticus) growth and development. MSc (Agric) thesis, University of Stellenbosch, South Africa. [ Links ]
Kruger, A.C.M., 2007. The effect of different management practices on the feed intake and growth rate of ostrich chicks. M.Tech. Thesis, Nelson Mandela Metropolitan University, South Africa. [ Links ]
Maja, M., 2012. VPN/04/2012-01 (Veterinary Procedural Notice - Revision 6.0): Standard for the requirements, registration, maintenance of registration and official control of ostrich compartments in South Africa. Department of Agriculture, Forestry and Fisheries - National Directorate Animal Health, pp. 1-42. [ Links ]
McDonald, P., Edwards, R.A., Greenhalgh, J.F.D., Morgan, C.A., Sinclair, L.A. & Wilkinson, R.A., 2011. Animal Nutrition. (7th ed.), UK, Europe. Pearson. p. 6. [ Links ]
Ott, R.L., 1998. An Introduction to Statistical Methods and Data Analysis., Belmont, California. Duxbury Press. pp. 807-837. [ Links ]
Robertson, J.B. & Van Soest, P.J., 1981. The detergent system of analysis and its application to human foods. In: Basic and Clinical Nutrition, Vol 3. Eds: James, W.P.T. & Theander, O. New York, USA, Marcel Dekker Inc. pp. 158-276. [ Links ]
Shapiro, S.S. & Wilk, M.B., 1965. An analysis of Variance Test for Normality (complete samples). In: Biometrika 52. pp. 591-611. [ Links ]
Smith, N., 2005. The effect of the dietary inclusion of canola oilcake, full-fat canola and sweet lupins on the production performance and fat composition of broilers and pigs. MSc (Agric) thesis, University of Stellenbosch, South Africa. [ Links ]
Swart, D., 1988. Studies on the hatching, growth and energy metabolism of ostrich chicks: Struthio camelus var. domesticus. Ph.D. Thesis, Stellenbosch University, South Africa. [ Links ]
Viviers, S.F., 2015. Effect of dietary energy and protein on the production parameters of slaughter ostriches (Struthio camelus var. domesticus). MSc (Agric) thesis, University of Stellenbosch, South Africa. [ Links ]
Von Baer, D., Reimerder, E.H. & Felcheim, W., 1978. Methoden zur Bestimmung der Chinolizidin alkaloide in Lupinus mutabilis. Zeitschrift für Lebensmittel Untersuchung und Forschung 16q, 27-31. [ Links ]
Received 16 October 2017
Accepted 11 March 2019
First published online 25 April 2019
# Corresponding author: tersb@elsenburg.com
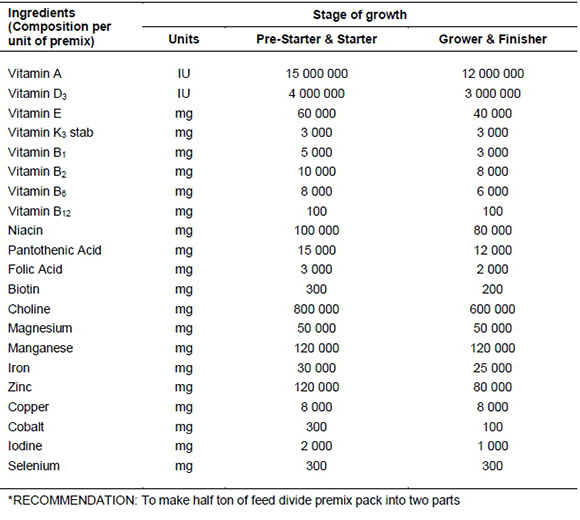
APPENDIX 1
Composition of the vitamin and mineral premix used in the four ostrich feeding phases (pre-starter, starter, grower and finisher) formulated per ton of feed


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}