










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.49 no.2 Pretoria 2019
http://dx.doi.org/10.4314/sajas.v49i2.11
ARTICLES
Fatty acids profile and oxidative stability of eggs from laying hens fed diets containing hemp seed or hempseed cake
D. Mierliță#
University of Oradea, Department of Animal Science, Oradea City, 410087, Romania
ABSTRACT
High levels of polyunsaturated fatty acids (PUFAs) are desirable in eggs for its nutritional quality, but render them vulnerable to oxidation. The aim of this trial was to assess the effects of dietary intake of hemp (seeds or cake) on the fatty acid (FA) profile and oxidative stability of eggs. The control diet (C), which was composed of corn, soybean meal and sunflower oil (2.5%), was compared with two experimental diets that were designed to replace sunflower oil with fat from hemp seed (HS diet) or hempseed cake (HC diet). One hundred and twenty Tetra-SL LL laying hens (24-week old) were used in a 10-week trial. Each treatment was replicated five times with eight birds each. Average hen-day egg production was not affected by feeding either the HS or the HC diet. The α-linolenic acid (ALA) concentration in eggs was increased by substituting the HS- or HC-based diets fed to the hens with dietary ALA. Similar deposition profiles were exhibited by eicosapentaenoic acid (EPA) and docosahexaenoic acids (DHA) in yolks in response to increasing the dietary ALA supply. The HS group showed a greater concentration of egg yolk ALA and EPA than the HC group, which had a higher concentration of linoleic acid (LA). These alterations in yolk composition resulted in n-6 : n-3 FA ratio values as low as 2.98 and 4.15 for HS and HC, respectively, compared to 11.07 for the control diet. The atherogenicity index and cholesterol level were not affected by hemp (seed or cake) inclusion, while the thrombogenicity index decreased when compared to the control diet. On days 0, 15 and 30 of storage (4 °C), two eggs were selected randomly from each replicate (totalling 10 eggs per treatment) and analyzed. The PUFAs were not affected by storage. An exception occurred in the HC group, in which eggs had lower n-6 FA content. Egg storage for 30 d led to a reduction in egg α-tocopherol and an increase of malondialdehyde (MDA) concentration, an indicator of lipid peroxidation. The HS treatment resulted in the lowest MDA (0.22 mg MDA/kg yolk for fresh eggs and 0.35 mg for eggs in 30-day storage). The study demonstrates that the level and type of PUFAs, level of α-tocopherol and duration of egg storage significantly affected the oxidative stability of eggs. The results obtained suggest that the inclusion of hemp seed appears to be more effective in maintaining the oxidative stability of egg lipids than hempseed cake.
Keywords: egg enrichment, α-tocopherol, malondialdehyde, omega-3, storage time
Introduction
Eggs are a food source for human consumption and contain most of the nutrients needed by humans. However, egg consumption should be limited due to the high content of saturated fatty acids (SFA) and cholesterol that increase the risk of developing many diseases (Attia et al., 2015). By contrast, the omega-3 fatty acids, especially α-linolenic acid (ALA, C18:3n-3), eicosapentaenoic acid (EPA, C20:5n-3) and docosahexaenoic acid (DHA, C22:6n-3), have positive effects on human health (Simopoulos, 2002). Therefore, maximizing the three fatty acid (FA) levels in eggs would be beneficial to human health.
A large number of studies have concluded that the FA composition of eggs is dependent on the FA composition of the feed of the laying hens, hence enriching eggs is done by enriching the feed of the laying hens with a source of n-3 PUFAs that will be assimilated in the eggs (Hammersh0j & Johansen, 2016; Nyberg, 2017). To increase the concentration of polyunsaturated fatty acids (PUFAs) in eggs, sources such as fish oil, linseed oil, corn oil, rapeseed oil, soybean oil and sunflower oil can be included in the diet of laying hens.
Hemp (Cannabis sativa L.) was initially grown for its fibre content and seed production in the Nordic countries, but was prohibited because of its high narcotic element (delta-9-tetrahydrocannabinol - THC) content. In 2003, hemp varieties with low concentrations of THC were allowed to be grown in the European Union (EU Council Directive, 2003) for fibre and oil production. Currently in the EU, only cultivars with less than 0.2% THC, as described in Council Regulation (EC) No 1420/98, are permitted. In 2012, the hemp cultivar, Armanca, was found in Romania to be a variety that could be utilized for oil seed production.
The increased production of hemp and the availability of hemp seed and hempseed products have created opportunities for use in livestock rations. Whole hemp seeds contain approximately 25% crude protein (CP), 33% to 35% oil and 34% carbohydrate, in addition to a broad range of vitamins and minerals (Gakhar et al., 2012). In a recent study, the inclusion of hemp seed and hempseed oil as a source of PUFAs, especially ALA, in the diet of laying hens was effective in improving the egg yolk FA profile by increasing the n-3 fatty acid concentration of eggs (Gakhar et al., 2012).
Chain length and the number of double bonds compromise n-3 FA oxidative stability. The hazard to lipid oxidation is higher in eggs enriched in EPA and DHA rather than ALA (González-Esquerra & Leeson, 2000). The oxidative damage of yolk lipids and overall reduction in egg quality is aggravated during storage (Hayat et al., 2010). Therefore, the control of lipid oxidation in such products is required to prevent the loss of nutritional value and to prevent the production of potentially toxic compounds. To maintain a high quality, the concentration of antioxidants should therefore be elevated. In egg yolks the concentration of α-tocopherol and carotenoids as antioxidants is believed to be important for oxidative stability (Criste et al., 2018). Dietary vitamin E supplementation has been shown to alleviate this problem with eggs (Hayat et al., 2010).
One potential opportunity for the use of hemp seed as a feed ingredient relates to the relatively high level of ALA (17% - 19%) (Parker et al., 2003) compared to other vegetable oils (<9%), though flax being a notable exception. Because flaxseed has been used successfully in the diets of laying hens to produce n-3 enriched eggs (Basmacioglu et al., 2003; Souza et al., 2008; Hayat et al., 2010; Petrovic et al., 2012) the level of ALA in hemp seeds or hempcake might provide additional options to improving egg quality. Hitherto, only a limited number of scientific reports can be found on hemp and its derived products in poultry diets and its nutritive and antioxidant efficacy in poultry (Gakhar et al., 2012; Konca et al., 2014). Consequently, the current study was designed to assess the effect of dietary intake of hemp seeds or hempcake on laying hen performance and egg yolk FA profile. As stated, high levels of PUFAs in eggs are desirable for its nutritional quality but this renders it vulnerable to oxidation. Accordingly, the secondary objective of the study was to examine the oxidative stability of yolk lipids in relation to the FA profile and tocopherols concentration of eggs throughout a 30-day refrigerated storage period, when the diet included a source rich in PUFAs.
Materials and Methods
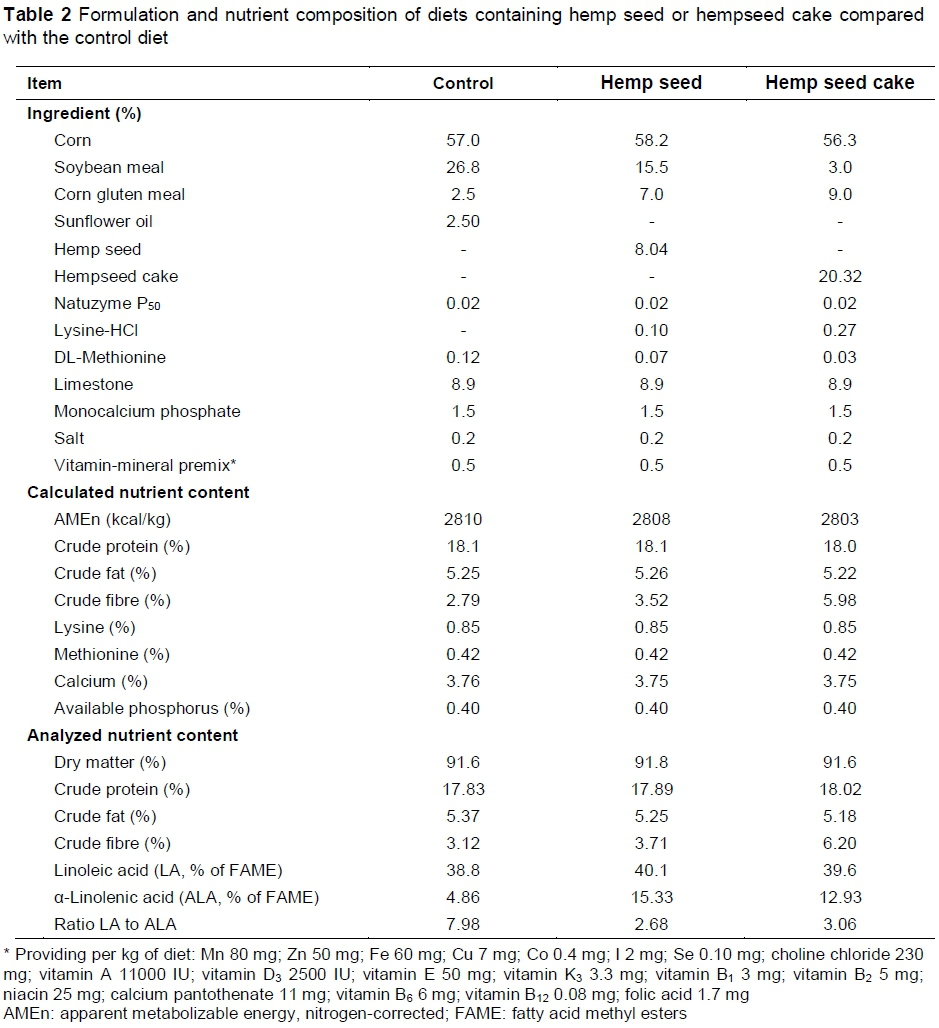
One hundred and twenty, 24-week-old Tetra-SL LL laying hens were assigned to three dietary treatments with five replicates of eight hens each (40 laying hens/treatment), in a completely randomized design. The control diet (C), which contained corn, soybean meal and sunflower oil (2.5%) as basic ingredients, was compared with two experimental diets that were designed to replace sunflower oil and part of the soybean meal in the control diet with fat from hemp seed (HS diet) or hempseed cake (HC diet).
The hens were fed a balanced commercial diet as their daily requirement for two weeks prior to commencement of the study to allow them to adapt and reach a standard level of egg production. Before starting the trial, the egg production of the hens was measured individually. Hens with equal egg production were blocked and within block were placed in each replicate. Each replicate of eight birds was housed in a cage of 1.0 mx1.5 m. A bird occupied an area of approximately 1875 cm2/hen. Feed and water were offered ad libitum during the experiment. Environmental temperature was set at 22 °C. A regimen of 14 h constant lighting (15 lux) and continuous ventilation were provided. All the birds were kept under uniform management conditions throughout the experimental period. The experiment lasted 10 weeks, with the hens reaching the age of 34 weeks.
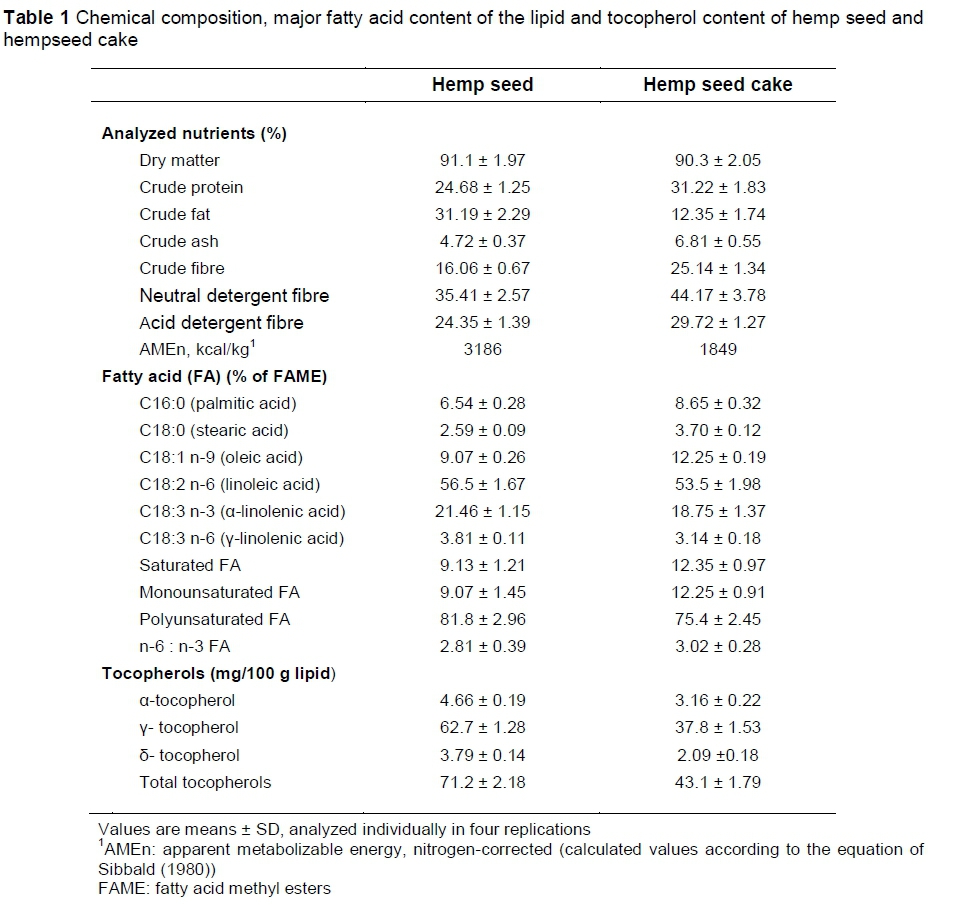
The hemp seeds (Cannabis sativa L., cv. Armanca variety for oil) were cold-pressed by a commercial oil producer (RosbroAvicom SRL), to obtain hempseed cake. The composition of the hemp seed and hempseed cake used for the formulation of diets was based on analysis realized in the laboratory. Samples were analyzed for dry matter (DM) (ISO, 1999), neutral detergent fibre (NDF) and acid detergent fibre (ADF) (Van Soest et al., 1991) on a fibre analyser (Ankom Technology, Fairport, NY), crude fat (Soxhlet method, AOAC, 1996) and crude protein (CP) (Kjeldahl technique) (Vapodest Distillation Systems - Gerhardt; AOAC, 1996) (Table 1). Hemp seed and hempseed cake were supplied in quantities calculated to provide the same amounts of fat, that is, 25 g/kg feed.
Eggs were collected twice daily at 9:30 am and 18:30. Feed intake was measured weekly for each replicate. Eggs collected over two days at the end of each week were weighed. All birds were weighed at 24 and 34 weeks of age. Eggs were collected for a period of eight consecutive days from all of the pens (two eggs per replicate) within each of the three treatments (a total of 240 eggs; 80 eggs/treatment; 16 eggs/ replicate) during weeks 9 and 10. Samples of the hemp seed, hempseed cake and experimental diet samples (n = 3) were collected and stored at -20 °C until analyzed for fatty acids.
On days 0, 15 and 30 of storage, two eggs were selected randomly from each replicate (totalling 10 eggs per treatment). The eggs were stored in a cold room at 4 °C. Lengths of storage times of 15 and 30 days were considered similar to the duration of consumer storage of shell eggs. Egg yolks were separated, and an aliquot was taken for FA profile, tocopherols concentration and lipid oxidation products, measured as TBA reactive substances (TBARS - thiobarbituric acid reactive substances).
The FA composition of the hemp seed, hempseed cake and dietary and yolk samples was determined using standard gas chromatographic techniques of fatty acid methyl esters (FAME) (AOAC, 1996). Fatty acids were extracted with a chloroform/methanol mixture according to Folch et al. (1957). At the first stage fat was saponified, using a 0.5N KOH methanol solution at 70 °C. Esterification with methanol was conducted in the presence of sulphuric acid as a catalyst.
Chromatographic analysis was performed using a Shimadzu GC-2010 gas chromatograph (Shimadzu Corporation, Tokyo, Japan) equipped with a DB-23 column (60 m x 0.25 mm i.d. and 0.25 μm film thickness). Column and detector temperatures were 190 and 240 °C, respectively. Carrier gas was helium at 1.0 mL/min ratio. The temperature of injection port was 230 °C with the split ratio of 1 : 80; temperature programme was 80°C/5 min, 200 °C/30 min and 230 °C/15 min (Criste et al., 2018). The FAME were identified using external standards (Supelco 37 Component FAME mix; Supelco Bellefonte, PA, USA), and the FA concentrations were calculated from the peak area of the corresponding FAs in relation to the total area of all peaks.
Egg yolks were also analyzed for total cholesterol content by the Washburn & Nix (1974) method, and calculated and expressed as milligrams per egg. Cholesterol was separated from fat after saponification with KOH and extracted with ethyl ether. The sample was subjected to chromatographic analysis under the following conditions: the length of a glass column, 1 m; internal diameter, 4 mm; film thickness, 0.25 μm; temperature of detector, 300 °C; temperature of injector, 290 °C; temperature of column, 260 °C; carrier gas, argon, flow rate, 50 cm3/min (Criste et al., 2018).
Extraction of tocopherols was done with a mixture of diethyl ether and petroleum ether (1 : 1), in aliquots of 20 mL. The ether phase, after separation, was saponified with methanolic KOH solution (10%), after which tocopherols were extracted in hexane, washed with water in a separating funnel, and evaporated to dryness. After evaporation of the hexane phase, separation of the tocopherols was by high performance liquid chromatography (HPLC) using the Parkin-Elmer LC-295 system, equipped with the Alltech C18 column (length 15 cm, internal diameter 4.6 mm and particle size 3 μm). The mobile phase for the various tocopherols was acetonitrile : methanol (85 : 15)/isopropanol 90/10. The content of tocopherol in the samples of hemp (seed and cake) and egg yolks were calculated according to external standards (Sigma-Aldrich, USA) through a linear regression from known standards (Criste et al., 2018).
Malondialdehyde (MDA) arising from the oxidation of PUFAs was chosen as a marker of secondary products of lipid oxidation. The MDA concentration was determined based the 2-thiobarbituric acid (TBA), as described by Salih et al. (1987) with some modifications by Cherian et al. (1996). Egg yolk samples (2 g) were weighed into test tubes, and perchloric acid added. The samples were homogenized for 15 s at high speed. Butylated hydroxytoluene (BHT) was added to each sample during homogenization to control lipid oxidation. The homogenate was filtered through filter paper. The filtrate (2 mL) was mixed with 2 mL of 20 mM TBA in distilled water and incubated in a boiling water bath for 30 min. Absorbance was determined at 531 nm. The oxidative stability was expressed in milligrams of MDA per kg of egg yolk.
Health lipid quality was assessed by calculating the atherogenic and thrombogenic indices as well as the ratio between the hypocholesterolemic and hypercholesterolemic, using the following equations:
• Atherogenicity index (AI) = [(4 x C14:0) + C16:0 + C18:0] /MUFA + PUFA n-6 + PUFA n-3 (Ulbricht & Southgate, 1991);
• Thrombogenicity index (TI) = (12:0 + 16:0 + 18:0)/[(0.5xMUFA)+(0.5xn-6 PUFA)+(3xn-3 PUFA)+(n-3 PUFA/n-6 PUFA)] (Ulbricht & Southgate, 1991);
• Hypocholesterolemic/hypercholesterolemic (h/H) = (C18:1 + PUFA)/(C12:0 + C14:0 + C16:0), (Fernandez et al. (2007).
The results of the analysis were compared to the analysis of variance (ANOVA) with the diet as principal effect (SAS, Software Version 8.2. SAS Institute, Inc., Cary, NC). The ANOVA for fatty acid level, α-tocopherol and MDA of yolk included the main effect of the diet, storage day and the interaction between these two main effects. In cases of significant differences, Duncan's multiple-range test was employed to compare differences among means obtained during different treatments. In addition, Pearson correlations were used to evaluate the relationships between egg yolk oxidative stability (MDA values) and α-tocopherol, PUFAs, and storage time. The level of significance to detect statistical differences was set at P <0.05 for all analyses.
Results and Discussion
Hemp seeds contained 24.7% CP and 31.2% crude fat (Table 1). The cold-pressing process removes 60.4% of the fat, and increased the CP level to 31.2%. These results are in line with those observed by Silversides & Lefrancois (2005), Halle & Schone (2013) and Neijat et al. (2014), who concluded that hemp products may serve as potentially valuable feed ingredients for laying hens.
Hempseed oil contained 21.4% linolenic acid, a lower concentration compared to flaxseed oil, which contains about 50% linolenic acid (Hayat et al., 2010). Hempseed oil also contains 56.5% linoleic acid, a higher concentration compared to flaxseed oil (15%) and rapeseed (21%). Finally, hempseed oil is richer in PUFA (81.8%) compared with the other oilseeds (camelina seed, 73%; flaxseed, 69%; sunflower, 66%; soybean, 60%; and rapeseed, 31%) (Hurtaud & Peyraud, 2007). Hemp seeds showed a high content of tocopherols (Table 1) which can ensure high oxidative stability of egg yolks.
All diets were balanced for energy, CP, crude fat, essential amino acids and minerals as required for laying hens (TETRA SL LL guidelines) (Table 2). The α-linolenic acid constituted 4.8% and 15.3% -12.9% of total FAs, and the linoleic acid (LA) : ALA ratios of the diets were 7.9 and 2.6 - 3.0 for the control and hemp diets, respectively.
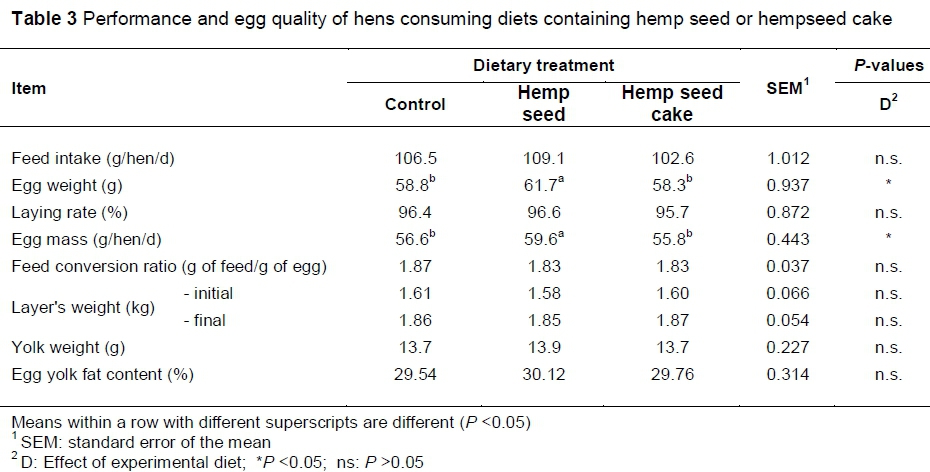
For hens consuming diets containing hemp seeds (HS group), egg weights and egg mass (g/hen/day) were significantly higher (P <0.05) than those observed in both the control and the HC diets (Table 3). With respect to other measures of production performance, the inclusion of hemp seeds or hempseed cake in laying hen diets did not affect feed intake, layer weight, feed conversion efficiency or hen-day egg production significantly. These results correspond with those of a previous study (Gakhar et al., 2012) which reported that the addition of hemp seed increased egg weight and did not lead to significant differences in feed intake, final bird weights and average hen-day egg production. On the other hand, Scheideler & Froning (1996) reported a positive effect on egg weight after feeding flaxseed at 5%, 10% and 15% levels for six weeks.
In this study no effects of the hemp diet were recorded in yolk weight or total lipid content. This agrees with the findings by Johansson (2010) where laying hens were fed diets containing hempseed cake. However, it has been reported that the increased levels of n-3 PUFAs in the diet affected yolk weight (Cabrera et al., 2006). Gonzalez-Esquerra & Leeson (2000) suggested that n-3 fatty acids in fish oil leads to a decrease in circulating triacylglycerols in blood plasma which may have a limiting effect on the availability of lipids for yolk formation
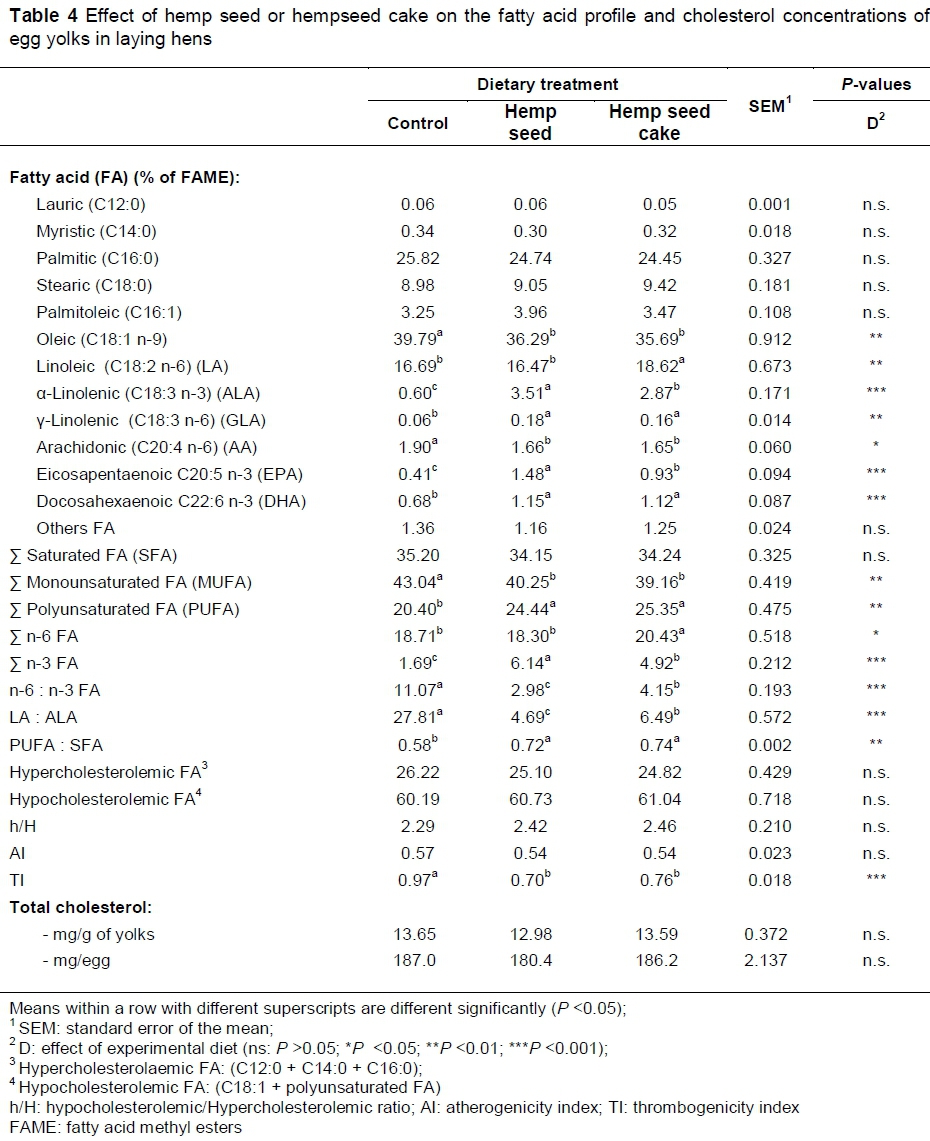
The fatty acid composition of the egg yolks is presented in Table 4. There was no significant difference between the SFAs in all the egg yolk samples. The C group had a lower PUFA concentration compared to the other groups, which was largely because of the low n-3 FAs.
Palmitic acid (C16:0), oleic acid (C18:1n-9) and linoleic acid (C18:2n-6) were the fatty acids with the highest concentrations in all egg yolk samples. Palmitic acid did not differ between the treatments whereas the oleic acid concentration was higher in C group compared to the HS and HC groups. Palmitic acid, oleic acid and linoleic acid were the most abundant fatty acids found in the egg yolks. This has also been shown in other studies (Omidi et al., 2015; Nyberg, 2017).
The total SFA concentration in egg yolks in this study ranged from 34.1% to 35.2%, and in the studies that were used for comparison, the SFA ranged from 30.8% to 38.8%, which shows that SFA is maintained within a very narrow range (Petrovic et al., 2012; Omidi et al., 2015; Nyberg, 2017). Cabrera et al. (2006) related this finding to the generally lower absorption of SFA, and the fact that hens tend to maintain an optimal saturated : unsaturated FA ratio for growth and hatchability of chicks. PUFAs (n-6 and n-3 series) in the diet are more effective in reducing MUFA than SFA in eggs through the inhibition of Δ-9 desaturase enzyme activity on the production of C18:1n-9 (Mazalli et al., 2004).
The HS and HC eggs did not show any difference in MUFA and PUFA concentration, but the egg yolks of hens in the HS group had a significantly higher amount of n-3 PUFA than the egg yolks of hens in the HC group; these having had a significantly higher amount of LA (C18:2n-6) in egg yolks. It is also important to note that the HS egg weight was higher compared to that of the HC eggs, which could also have an impact on fatty acid composition, but that was not investigated in this experiment.
The hemp product inclusion in the diet (HS and HC) led to an increase in the concentration of ALA in the laying hen diet and led to increases in the individual n-3 fatty acids (ALA, EPA and DHA) and the total n-3 fatty acid concentration in eggs yolks. The overall increase in the total n-3 content of the egg yolks occurred at the expense of the level of oleic acid, which decreased at inclusion of HS or HC in the laying hen diet. The ALA in the eggs from the hens fed the HS and HC diets were 5.8 and 4.7 times, respectively, higher than in the control eggs. Gakhar et al. (2012) and Souza et al. (2008) reported similar results, where higher levels of this FA in the eggs from the hens fed hemp seed and flaxseed or its oil were observed compared to those with corn, soybean, canola and sunflower oils. The LA concentration was significantly lower (P <0.01) in the eggs produced by the hens fed diets of hemp seed and the control diet.
The inclusion of hemp seed or hempseed cake dramatically reduced the LA : ALA ratio in egg yolks from 27.8 : 1 in the control diet to 4.6 - 6.4 : 1 in the HS and HC diets (see Table 4). This is due to the high amount of ALA in hemp seed and hempseed cake, consistent with the findings of previous studies which showed a reduced LA : ALA ratio in eggs from hens consuming diets containing hemp seed (Gakhar et al., 2012; Raza et al., 2016) or hempseed oil (Neijat et al., 2016).
Hemp seed has a high concentration of ALA (>21% - Table 1), which makes it a good ingredient for n-3 fatty acid enrichment in egg yolk, but, unfortunately, hemp seed lacks the n-3 FAs long-chain of EPA and DHA. The present study has shown that hemp seed and hempseed cake significantly increased both EPA and DHA in egg yolk. This is because hens have a high ability to convert ALA to n-3 FA long-chain (Howe et al., 2002). Hempseed cake is less efficient than whole raw seed in increasing eggs yolk ALA and EPA content. Van Elswyk (1997), comparing different sources of n-3 PUFA enrichment in eggs, showed that hens fed ground or crushed flaxseed showed higher levels of EPA and DHA in the yolk. The intake of ALA appeared to be the primary determinant of the total n-3 fatty acid content of egg yolks (Gakhar et al., 2012). Conversion of ALA to EPA and DHA occurs through the sequential actions of Δ-6 desaturase, elongase and Δ-5 desaturase (Gakhar et al., 2012).
The amount of AA (C20:4n-6) in egg yolk decreases in HS and HC diets. The results obtained by Petrovic et al. (2012) show a significant decrease in AA content which was in correlation with the amount of n-3 PUFA added to the feed (r = -0.958). The increased ingestion of ALA in a diet results in a significant decrease in the formation of AA (da Silva et al., 2009) due to the greater utilization of Δ-6 desaturase in the n-3 FA with respect to the n-6 pathway. Thus, the high level of ALA limits the synthesis of AA from LA, because ALA competes with LA by the same Δ-6 desaturase enzymes (Cabrera et al., 2006; Ceylan et al., 2011).
Our results show that enrichment of laying hens' diets with n-3 PUFA does not affect the level of cholesterol in eggs, which is in accordance with the results of Ceylan et al. (2011) who demonstrated that different dietary oil sources rich in n-3 PUFA (fish oil, linseed oil, rapeseed oil) at two levels of inclusion (1.5% and 3.0%) in laying hen diets, affected FA composition of the egg yolk, while cholesterol content was not influenced. It has been hypothesized that the inability to markedly reduce egg cholesterol levels is due to a physiological control mechanism that ultimately causes the cessation of egg production when yolk cholesterol deposition is inadequate for embryo survival (Ceylan et al., 2011). In contrast to our finding, the study conducted by Shahid et al. (2015) showed that the yolk cholesterol level of laying hens whose diet had been treated with hemp seed at a rate of 15% - 25% was significantly lower (P <0.05), compared to the control group. The results of a study by Johansson (2010) suggest that hemp seed may lower the cholesterol levels in egg yolk.
The PUFA/SFA and n-6/n-3 PUFA ratios, atherogenity index (AI) and thrombogenity index (TI) are commonly used to assess the nutritional value of animal fat on consumer health. In general, a ratio of PUFA to SFA above 0.45 and a ratio of n-6/n-3 below 4.0 are required in human diets to combat 'lifestyle diseases' such as coronary heart disease and cancer (Simopoulos, 2002). In the present study, the PUFA/SFA ratios (0.57 - 0.74 : 1) were higher than the recommended values for all groups, whereas the n-6/n-3 FA ratios were within the recommended levels for the HS and HC groups (2.98 - 4.15 : 1), and considerably higher than the recommended values for C group (11.07:1) (Table 4). Basmacioglu et al. (2003) reported a decrease in the n-6/n-3 FA ratio from the usual value in commercial eggs (12:1) to a value below 3 : 1 using feed enriched with fish oil, linseed oil, or a combination of both, results which are in line with those of our study. The introduction of hemp seeds in laying hen feed (HS group) provided the most beneficial ratios of PUFA/SFA (0.71) and n-6/n-3 PUFA (2.98) in egg yolk. The high levels of ALA, EPA and DHA in the egg yolk of HS and HC groups are partly countered by the contents of MUFA, in particular oleic acid.
As shown in Table 4, the inclusion of hemp seed and hempseed cake in the diet decreased the value of TI, but did not alter the AI value and the hipocholesterolemic /hipercholesterolemic FA ratio in egg yolk.
The values of n-3 FA, ALA and EPA, higher, and n-6/n-3 FA ratio lower in HS than HC (P <0.05), suggest that the inclusion of hemp seed, rather than hempseed cake, appears to be more effective in ameliorating these indices in egg yolk. Storage had no effect on the concentration of SFA in eggs from the HS and HC groups (Table 5). However, on day 15 and day 30, a reduction in SFA and an increase in MUFA were observed in the control group. During storage (day 20 through to day 60), an increase (>4% to 9%) in MUFA content, mainly on the basis of oleic acid, in egg yolks, was observed by Hayat et al. (2010).
PUFAs were not affected by storage. An exception occurred in the HC group, where the day 15 and day 30 eggs showed a lower n-6 FA content than day 0 eggs (P <0.05). The n-3 FA in egg was stable during storage in all treatments. The reduction in n-6 FA in HC eggs during storage suggests that hempseed cake tocopherols were not effective in minimizing the degradation of PUFAs in eggs stored.
The changes in n-6 : n-3 fatty acid ratios upon storage occurred in eggs from hens fed the control diet, when compared with the HS and HC groups, suggesting the role of α-tocopherol in modulating the desaturase and elongating the pathway in a favourable way.
Maximum α-tocopherol concentration was observed in the eggs from hens fed hemp seed (Table 5). Storage led to a reduction in egg α-tocopherol content in all groups. This reduction was increased between 15 day and 30 day of storage. At day 30 of storage, cumulative decreases in egg α-tocopherol of 30.7%, 25.6% and 28.3% for eggs from hens from the C, HS and HC groups, respectively, were observed. These findings corroborate previous results of Hayat et al. (2010), who noticed a decline of 28% - 32% in α-tocopherol concentration of eggs after 40 days of storage at 4 °C. The difference in the extent of α-tocopherol decrease may be due to the FA composition of the eggs.
The alpha isomer of tocopherol (vitamin E) has a high value in animal nutrition; yet, is relatively poor as an antioxidant (Helm, 2006). In contrast, the beta-, gamma- and delta-tocopherols are not nearly as effective as nutrients, but are 130, 200 and 500 times more powerful, respectively, in their antioxidant capacities (Helm, 2006). The primary tocopherol in hemp seed and hempseed cake is the gamma isomer (88.1% and 87.8% respectively, of total tocopherols) as its major antioxidant, with minor amounts of α-tocopherol (Table 1).
Malondialdehyde (MDA) is the main product of oxidation of PUFAs and it is widely used as a marker of lipid peroxidation in food. The MDA values of lipids in storage eggs were higher than those in fresh eggs. The highest MDA values related to storage were found in eggs from the control diet. The extent of lipid oxidation differed among the dietary treatments only in 30-day stored eggs, and revealed the lowest lipid oxidation in group HS. The lower than expected oxidation in relation to the n-3 FA content noted in the HS eggs may be attributed to the higher concentration of α-tocopherol in the yolk. The high MDA in the HC eggs may be attributed to the lower concentration of α-tocopherol in the yolk, possibly due to the lower amounts of total tocopherols in the hempseed cake (hempseed cake contributes 10.8 mg to the total tocopherols/kg of diet compared to hemp seed which contributes 17.7 mg to the total tocopherols/kg of the diet). Supplementing the diets of laying hens with α-tocopheryl acetate increases, in a dose dependent manner, the content of α-tocopherol in eggs (16.6, 49.8, 78.9 and 132.3 μg/g of egg for 0, 50, 100 and 200 mg/kg α-tocopheryl acetate respectively, in the diet) and significantly reduces lipid oxidation (Galobart et al., 2001).
These studies demonstrate that the level and type of PUFA, the level of α-tocopherol and duration of egg storage significantly affected the oxidative stability of eggs. There is an inverse relationship between MDA content, an indicator of egg yolk lipid peroxidation, and α-tocopherol level in eggs. Ren et al. (2013) also found that the MDA content in yolk increases significantly when stored at 4 °C for 28 days, and storage also results in a reducing the proportion of PUFAs and increases that of MUFA in egg yolks.
The correlation analysis showed the potential relationship between PUFAs and tocopherol content and lipid oxidation of storage eggs (Table 6). The most important observation is the presence of positive relationships of storage time and n-3 FA level, with MDA, indicating that an increase of storage time and n-3 FA level accompanies an increase in MDA, which is consistent with our results. In addition, there was an inverse correlation between yolk fat MDA concentrations and yolk α-tocopherol levels (r = -0.348, P <0.01) indicating that tocopherols from hemp products have positive potential as a natural antioxidant and could contribute to preventing lipid oxidation in egg yolks.
Conclusions
The inclusion of HS and HC in the diets of laying hens does not have an adverse effect on the performance of laying hens, and these products can be included in the diets of laying hens to serve as alternative sources of ALA for the production of eggs with higher levels of total n-3 content. The diets do not significantly affect cholesterol content in yolk. Egg storage for 30 days led to a reduction in egg α-tocopherol and an increase in MDA content. The oxidative stability of lipids measured as MDA content in egg yolks was improved by hemp seed supplementation. The results of this study (the increase egg weight, the values of ALA, EPA, total n-3 FA higher and n-6/n-3 FA ratio, and MDA content lower in HS than HC) suggest that the inclusion of hemp seed in the diet of the hens appears to be more effective in ameliorating nutritional quality of eggs than hempseed cake.
Acknowledgements
This research was conducted in the context of the project "PN-II-IN-CI-2012-1-0257" sponsored by UEFISCDI-Romania. I'm are also grateful to RosbroAvicom SRL for providing the experimental facilities and supplying the hemp seed.
Conflict of Interest Declaration
The author declares that there was no conflict of interest.
References
AOAC, 1996. Official Methods of Analysis.Volume I. 16th ed. Association of Official Analytical Chemists, Arlington, VA., USA. [ Links ]
Attia, Y.A., Al-Harthi, M.A., Korish, M.A. & Shiboob, M.M., 2015. Fatty acid and cholesterol profiles and hypocholesterolemic, atherogenic, and thrombogenic indices of table eggs in the retail market. Lipids Health Dis. 14, 136-144. [ Links ]
Basmacioglu, H., Cabuk, M., Unal, K., Ozkan, K., Akkan, S. & Yalcin, H., 2003. Effects of dietary fish oil and flax seed on cholesterol and fatty acid composition of egg yolk and blood parameters of laying hens. S. Afr. J. Anim. Sci. 33, 266-273. [ Links ]
Cabrera, M.C., Saadoun, A., Grompone, A., Pagano, T., Salhi, M., Olivero, R. & del Puerto, M., 2006. Enriching the egg yolk in n-3 fatty acids by feeding hens with diets containing horse fat produced in Uruguay. Food Chem. 98, 767-773. [ Links ]
Ceylan, N., Ciftçi, I., Mizrak, C., Kahraman, Z. & Efil, H., 2011. Influence of different dietary oil sources on performance and fatty acid profile of egg yolk in laying hens. J. Anim. Feed Sci. 20, 71-83. [ Links ]
Cherian, G., Wolfe, F.H. & Sim, J.S., 1996. Dietary oils with added tocopherols: effects on egg or tissue tocopherols, fatty acids, and oxidative stability. Poult. Sci. 75, 423-431. [ Links ]
Council Regulation (EC) No 1420/98 of 26 June 1998 amending Regulation (EEC) No 619/71 laying down general rules for granting aid for flax and hemp. [ Links ]
Criste, F.L., Mierlitã, D., Simeanu, D., Boisteanu, P., Pop, I.M., Georgescu, B. & Nacu, G., 2018. Study of fatty acids profile and oxidative stability of egg yolk from hens fed a diet containing white lupine seeds meal. Rev. Chim. (Bucharest) 69 (9), 2454-2460. [ Links ]
da Silva, W.A., Elias, A.H.N., Aricetti, J.A., Sakamoto, M.I., Murakami, A.E., Gomes, S.T.M., Visentainer, J.V., De Souza, N.E. & Matsushita, M., 2009. Quail egg yolk (Coturnixcoturnix japonica) enriched with omega-3 fatty acids. Food Sci. Technol. 42, 660-663. [ Links ]
EU Council Directive, 2003. EU Council Directive no. 1782/ 2003. Official Journal of the European Communities, L270/52, 1920. [ Links ]
Fernandez, M., Ordonez, J.A., Cambero, I., Santos, C., Pin, C. & De la Hoz, L., 2007. Fatty acid compositions of selected varieties of Spanish dry ham related to their nutritional implications. Food Chem. 9, 107-112. [ Links ]
Folch, J., Lees, M. & Stanley, G.H.S., 1957. A simple method for the isolation and purification of total lipid from animal tissues. J. Biol. Chem. 226, 497-509. [ Links ]
Gakhar, N., Goldberg, E., Jing, M., Gibson, R. & House, J.D., 2012. Effect of feeding hemp seed and hemp seed oil on laying hen performance and egg yolk fatty acid content: Evidence of their safety and efficacy for laying hen diets. Poult. Sci. 91, 701-711. [ Links ]
Galobart, J.A., Barroeta, C., Baucells, M.D., Cortinas, L. & Guardiola, F., 2001. α-Tocopherol transfer efficiency and lipidoxidation in fresh and spray-dried eggs enriched with w3-polyunsaturated fatty acids. Poult. Sci. 80, 1496-1505. [ Links ]
González-Esquerra, R. & Leeson, S., 2000. Effect of feeding hens regular or deodorized menhaden oil on production parameter, yolk fatty acid profile and sensory quality of eggs. Poult. Sci. 79, 1597-1602. [ Links ]
Halle, I. & Schone, F., 2013. Influence of rapeseed cake, linseed cake and hemp seed cake on laying performance of hens and fatty acid composition of egg yolk. J. Verbr. Lebensm. 8, 185-193. [ Links ]
Hammershoj, M. & Johansen, N.F., 2016. Review: The effect of grass and herbs in organic egg production on egg fatty acid composition, egg yolk colour and sensory properties. Livest. Sci. 194, 37-43. [ Links ]
Hayat, Z., Cherian, G., Pasha, T.N., Khattak, F.M. & Jabbar, M.A., 2010. Oxidative stability and lipid components of eggs from flax-fed hens: Effect of dietary antioxidants and storage. Poult. Sci. 89, 1285-1292. [ Links ]
Helm New York, Inc., 2006. Difference between antioxidant and vitamin activity: Natural tocopherol isomers. http://www.helmnewyork.com/en/products/specification/tocopherols-natural-mixed.html [ Links ]
Howe, P.R.C., Downing, J.A., Grenyer, B.F.S., Grigonis-Deane, E.M. & Bryden, W.L., 2002. Tuna fishmeal as a source of DHA for n-3 PUFA enrichment of pork, chicken, and eggs. Lipids 37, 1067-1076. [ Links ]
Hurtaud, C. & Peyraud, J.L., 2007. Effects of feeding Camelina (seeds or meal) on milk fatty acid composition and butter spreadability. J. Dairy Sci. 90, 5134-5145. [ Links ]
ISO (International Organization for Standardization), 1999. Animal feeding stuffs. Determination of moisture and other volatile matter content. ISO, Geneva, Switzerland. [ Links ]
Johansson, A., 2010. Effects of genotype age and feed on the fat components of egg yolk. Ph.D. thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden. [ Links ]
Konca, Y., Cimen, B., Yalcin, H., Kaliber, M. & Beyzi, S.B., 2014. Effect of hempseed (Cannabis sativa sp.) inclusion to the diet on performance, carcass and antioxidative activity in Japanese Quail (Coturnixcoturnix japonica). Korean J. Food Sci. An. 34 (2), 141-150. [ Links ]
Mazalli, M.R., Faria, D.E., Salvador, D. & Ito, D.T., 2004. A comparison of the feeding value of different sources of fat for laying hens: 2. Lipid, cholesterol, and vitamin E profiles of egg yolk. J. Appl. Poult. Res. 13, 280-290. [ Links ]
Neijat, M., Gakhar, N., Neufeld, J. & House J.D., 2014. Performance, egg quality, and blood plasma chemistry of laying hens fed hempseed and hempseed oil. Poult. Sci. 93, 2827-2840. [ Links ]
Neijat, M., Suh, M., Neufeld, J. & House, J.D., 2016. Hempseed products fed to hens effectively increased n-3 polyunsaturated fatty acids in total lipids, triacylglycerol and phospholipid of egg yolk. Lipids 51, 601-614. [ Links ]
Nyberg, J., 2017. Analysis of fatty acids in egg yolks of various production systems. Master's thesis. http://stud.epsilon.slu.se. [ Links ]
Omidi, M., Rahimi, S. & Torshizi, M.A.K., 2015. Modification of egg yolk fatty acids profile by using different oil sources. Vet. Res. Forum 6 (2), 137-141. [ Links ]
Parker, T.D., Adams, D.A., Zhou, K., Harris, M. & Yu, L., 2003. Fatty acid composition and oxidative stability of cold-pressed edible seed oils. J. Food Sci. 68, 1240-1243. [ Links ]
Petrovic, M., Gacic, M., Karacic, V., Gottstein, Z., Mazija, H. & Medic, H., 2012. Enrichment of eggs in n-3 polyunsaturated fatty acids by feeding hens with different amount of linseed oil in diet. Food Chem. 135, 1563-1568. [ Links ]
Raza, T., Chand, N., Khan, R.U., Shahid, M.S. & Abudabos, A.M., 2016. Improving the fatty acid profile in egg yolk through the use of hempseed (Cannabis sativa), ginger (Zingiber officinale), and turmeric (Curcuma longa) in the diet of Hy-Line White Leghorns. Arch. Anim. Breed. 59, 183-190. [ Links ]
Ren, Y., Perez, T.I., Zuidhof, M.J., Renema, R.A. & Wu, J., 2013. Oxidative stability of omega-3 polyunsaturated fatty acids enriched eggs. J. Agr. Food Chem. 61, 11595-11602. [ Links ]
Salih, A.M., Smith, D.M. & Dawson, L.E., 1987. Modified extraction 2-thiobarbituric acid method for measuring lipid oxidation in poultry. Poult. Sci. 66, 1483-1488. [ Links ]
SAS, 2001. SAS System for Microsoft Windows. Release 8.2.SAS Institute Inc., Cary, NC., USA. [ Links ]
Scheideler, S.E. & Froning, G.W., 1996. The combined influence of dietary flaxseed variety, level, form, and storage conditions on egg production and composition among vitamin E-supplemented hens. Poult. Sci. 75, 1221-1226. [ Links ]
Shahid, S., Chand, N., Khan, R., Suhail, M. & Khan, N., 2015. Alternations in cholesterol and fatty acids composition in egg yolk of Rhode Island red x Fyoumi hens fed with hemp seeds (Cannabis sativa L.). J. Chem. http://dx.doi.org/10.1155/2015/362936. [ Links ]
Sibbald, I.R., 1980. Metabolisable energy in poultry nutrition. Bio.Sci. 30, 736-741. [ Links ]
Silversides, F.G. & Lefrancois, M.R., 2005. The effect of feeding hemp seed meal to laying hens. Br Poult Sci. 46, 231-235. [ Links ]
Simopoulos, A.P., 2002. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed Pharmacother 56 (8), 365-379. [ Links ]
Souza, J., Costa, P., Queiroga, E., Silva, V., Schuler, P. & Goulart, C., 2008. Fatty acid profile of eggs of semi-heavy layers fed feeds containing linseed oil. Braz. J. Poult. Sci. 10, 37-44. [ Links ]
***TETRA SL LL, 2017. Commercial Management Guide, Bábolna Tetra Ltd., Radnóti, Hungary. http://www.babolnatetra.com/uploads/pdf/2017-02-23_TETRA_SL_LL_PARENT_management-guide_PRESS.pdf. [ Links ]
Ulbricht, T.L.V. & Southgate, D.A.T., 1991. Coronary heart disease: Seven dietary factors. Lancet 338, 985-992. [ Links ]
Van Elswyk, M.E.V., 1997. Comparison of n-3 fatty acid sources in laying hen rations for improvement of whole egg nutritional quality: a review. Br. J. Nutr. 78, 61-69. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Washburn, K.W. & Nix, D.F., 1974. A rapid technique for extraction of yolk cholesterol. Poult. Sci. 53, 1118-1122. [ Links ]
Received 22 October 2018
Accepted 2 March 2019
First published online 21 April 2019
# Corresponding author: dadi.mierlita@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}