










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.6 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i6.16
ARTICLES
SHORT COMMUNICATION
Effects of non-genetic factors on the inter-calving period of Nguni cows in South Africa
F.N. MkhizeI; E.C. WebbI, #; M.M. ScholtzII, III
IDepartment of Animal and Wildlife Sciences, University of Pretoria, Private Bag X20, Hatfield, 0028, South Africa
IIAgricultural Research Council, Private Bag X2, Irene, 0062, South Africa
IIIUniversity of the Free State, Bloemfontein, 9300, South Africa
ABSTRACT
The purpose of this study was to evaluate the effect of geographical regions of South Africa on the inter-calving period of Nguni cows. Data of Nguni cows (n = 26 681) from 427 Nguni breeders in the Savannah and the Grassland Biomes were statistically analyzed to establish the impact of non-genetic factors on the inter-calving period (ICP) of these cows. Cows with an ICP of less than 315 and above 730 days were removed from the dataset before analysis, to comply with Nguni breed standards. Dam age was included as a covariant. The ICP increased slightly up to 48 months of age, after which a decline was noticed up to the age of 108 months. Cows in the Savannah Biome had a shorter ICP than those studied in the Grassland Biome. Within the Savannah Biome, the ICP of cows was significantly shorter in the Eastern Kalahari Bushveld bioregion than in any other bioregion, while in the Grassland Biome the ICP was significantly shorter for cows calving in the Drakensberg and the Dry Highveld bioregions. In terms of seasons, the ICP of cows bred in spring and summer was significantly shorter than that of cows bred in autumn and winter. Categorization of cows in year groupings indicates significant year effects.
Keywords: Biome, bioregion, dam age, reproduction, season
Introduction
The reproductive efficiency of cows is a key factor in the increase in numbers of any beef herd (Ladermann & Schoeman, 1994). One of the factors that is related to cow productivity is the inter-calving period (ICP), which is defined as the time period from one calving to the next. It has been described as a measure that involves three phases, namely the gestation period, postpartum anestrus, and the subsequent breeding period. Of the three phases, the postpartum period (from the last calving to the next oestrus) can be improved through management. According to Mukasa-Mugerwa (1989), the postpartum period which is instrumental in the re-establishment of the ovarian activity and preparation of the cow for re-conception can be shortened by proper nutrition. The postpartum period is the most critical of the three phases to ensure that the ICP is within the target range of less than 400 days on average for the herd if a cow is to calve every year. It is therefore recommended that the postpartum period should not exceed 80 - 85 days. The ICP is a major factor in the determination of the reproductive efficiency of a cow and it is affected, among others, by the availability and management of nutrition (Mukasa-Mugerwa, 1989).
Experience in practical situations as well as research have confirm ed that the productivity of an animal depends on its ability to maintain physiological activities at levels that allow it to grow and reproduce in a particular environment (Gregory, 1972; Van der Westhuizen, 1973; Maree & Casey, 1993; Webb et al., 2017). This is described as adaptability and may develop as a result of selection. Different cattle breeds differ in the way in which they are able to withstand and react to climatic stress, diseases, and low feed levels.
Factors such as rainfall, temperature and soil type determine and have an influence on the type of vegetation in each region (Acocks, 1988; Low & Rebelo, 1996). It is expected that an animal will perform well in an environment to which it is adapted (Bonsma, 1983). The Nguni breed is perceived to be adapted to harsh climatic conditions such as high temperatures resulting in poor feed quality, and the occurrence of ticks and tick-borne diseases (Schoeman, 1989; Collins-Lusweti, 2000; Muchenje et al., 2008). It can be assumed that this breed will perform better than others in most geographic areas as defined by these climatic conditions. The Savannah Biome is most suitable for livestock and/or wildlife grazing because of its good grass cover, shrubs and trees (Low & Rebelo, 1996). The Grassland Biome is generally characterized by large areas dominated by grasses and fewer trees, which renders it particularly suitable for cattle farming. The difference between the two biomes is largely the result of climatic variances, notably rainfall and temperature. These climatic differences have an effect on the types of grasses that grow in the different areas, and the quality of grazing at different times of the year. Although calf sex and culling rate were recorded in this study, these parameters were not discussed in detail due to their insignificant effect. The purpose of this study was to address the question whether non-genetic factors such as vegetation type and availability of feed sources have any effect on the reproduction of Nguni cattle, in particular on the ICP.
Materials and Methods
Data were obtained from the Integrated Registration and Genetic Information Systems (INTERGIS) of the Agricultural Research Council (ARC) of the Republic of South Africa, and involved 26 681 Nguni cows. The data were collected previously from 427 Nguni breeders in the Savannah and the Grassland Biomes of South Africa. Nguni breeders from the Savannah Biome constituted 57% of the data, while 28.8% of the Nguni breeders were located in the Grassland Biome. Both biomes are in the summer rainfall areas of South Africa. Within the two biomes, eight bioregions were identified, four in each biome. Bioregions in the Savannah Biome were identified as the Central Bushveld, Eastern Kalahari Bushveld, Lowveld and Sub-Escarpment Savannah. The four bioregions which form part of the Grassland Biome are the Drakensberg, Dry Highveld bioregion, Mesic Highveld and the Sub-Escarpment grassland. Data collected over a period of 20 years (1990-2010) were categorized into four five-year groupings:1990-1995 - year grouping 1; 1996‑2000 - year grouping 2; 2001-2005 - year grouping 3; and 2006-2010 - year grouping 4. The groupings were divided into calving seasons, namely summer (November, December, January); autumn (February, March, April); winter (May, June, July); and spring (August, September, October).
Data that were identified as outliers and could not be used for analysis were removed before performing analysis. This included breeders with fewer than 50 cows and breeders that could not be allocated into specific biomes. Cows that were older than 39 months at first calving were excluded from the analysis. Cows with an ICP of less than 315 and above 730 days were also removed from the dataset before analysis, which constituted 1.86% and 2.04% of records, respectively. The breed average ICP of 409 days was used to compare the efficiency of Nguni cows in this study. These limits are in line with the Nguni breed standards and will ensure that the study can be translated into practical and realistic production practices for the Nguni breeders, country-wide.
Data were analyzed statistically using the General Linear Method (GLM) of SAS (2017) to establish the impact of non-genetic effects on the ICP as a measure of reproduction efficiency for all cows in the herd. The main factors considered in the analysis were biomes, bioregions, seasons, and year categories. Dam age was included as a covariant.
Results and Discussion
The inter-calving period (ICP) increased slightly up to 48 months of age, probably because first-calf cows tended to have a longer ICP, after which a decline was noticed up to 108 months of age, as shown in Figure 1. Mukasa-Mugerwa (1989) noted that if the cow is not in an ideal body condition, it will not conceive. As can be seen in Figure 1, the decrease in number of productive cows after the age of 108 months rendered conclusions unreliable. The observation is that a longer ICP plays a role in reducing the number of cows in production, since such cows will eventually be culled from the herd.
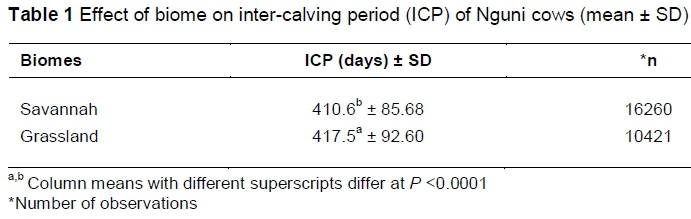
The different biomes had a significant effect (P <0.0001) on the mean ICP of the total cow herd. Cows in the Savannah Biome had a significantly shorter mean ICP, compared to those in the Grassland Biome (Table 1). The Savannah Biome has more grasses and shrubs (Low & Rebelo, 1996), compared to the Grassland biome. The types of grasses that occur in any biome are dictated by the climate, types of soil and grazing of the biome; however, the management of the grasses can alter the type of grasses in a biome. According to Tainton (1999), palatability and quality of grasses are better due to spring re-growth, after which the grass matures, resulting in reduced quality and palatability which affect cattle performance. Tainton (1999) further explains that the frequency of veld fires and overgrazing of grasses may change the grass communities. Overgrazing generally results in the emergence of unpalatable and grazing-tolerant species which in sour grassveld areas is an irreversible action. In this study, it is believed that the quality of grasses in the Savannah Biome resulted in better performance by the cowherd, compared to the Grassland Biome. Reducing the postpartum period has been noted as a critical measure (Mukasa-Mugerwa, 1989; Messine et al., 2004; Burns et al., 2010) to improve reproduction as well as the cow's productive life. According to Messine et al. (2004), suckling was noted to contribute substantively to longer ICPs in beef cattle, especially in Zebu cattle. This is also enhanced by the time of day for suckling as it has a highly significant effect on the resumption of ovarian activity in beef cattle (Escrivao et al., 2012). It must be noted that forage quality and quantity contribute to the necessary nutrition required to ensure that the cows are in good condition at breeding so that conception will be successful soon after parturition and thus shorten the postpartum period.
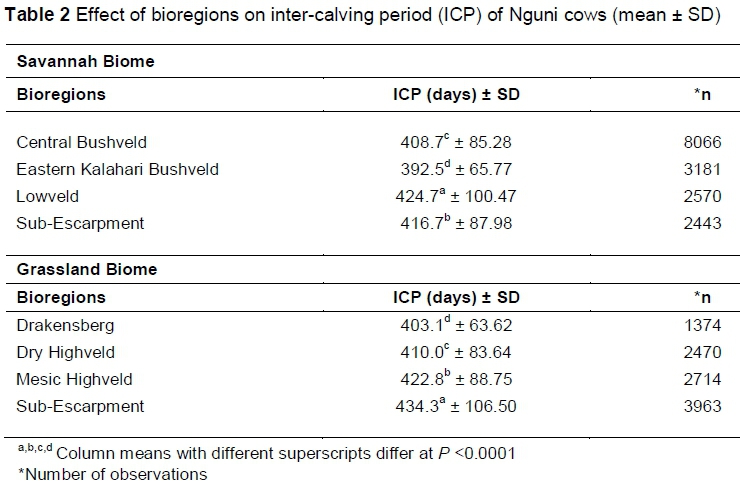
Bioregions had different effects (P <0.0001) on the ICP of Nguni cows within biomes (Table 2). In the Savannah Biome, the mean ICP was significantly shorter for cows bred in the Eastern Kalahari Bushveld bioregion than for cows bred in the other bioregions in the same biome. In the Grassland Biome, cows bred in the Drakensberg bioregion had a significantly shorter mean ICP than cows bred in the other bioregions of the Grassland Biome. This shows the effect of these bioregions on the productivity of Nguni cattle, despite the renowned adaptability of the breed. Cows bred in different parts of the Savannah or Grassland Biome can therefore be expected to differ in their reproductive efficiency as measured by the ICP. The variation of ICPs within bioregions is smaller relative to other breeds. In a study conducted on Bonsmara breeds, Webb et al. (2017) reported an ICP median of 407.7 days, compared to a mean of 422.8 days.
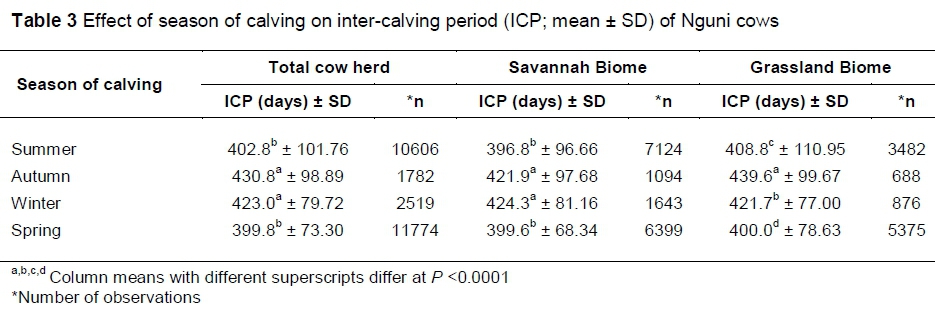
Calving seasons showed a statistically significant effect on the ICP of Nguni cows. Cows calving in spring and summer had shorter ICPs, compared to those calving in autumn and winter, as shown in Table 3. Some researchers (Bergh, 2004; Grobler et al., 2014) who explored seasonal breeding agree that choosing the breeding season carefully, results in improved weaning weights, reduces calf mortalities and also ensures good body condition even at lactation. In fact, seasonal breeding is one of the methods according to which the ICP can be reduced by ensuring good body condition of the cow after giving birth.
The interaction between biome and season of calving influenced (P <0.0001) the ICP of cows. In the Savannah Biome, the mean ICP was significantly shorter for cows calving in summer and spring, compared to those calving in autumn and winter. In the Grassland Biome, the ICP of cows calving in spring was significantly shorter, compared to that of cows calving in the other three seasons. In both biomes, autumn and winter seasons had the longest mean ICP, as shown in Table 3. Previous research has shown that seasonal effects on the productivity of cows can be reduced by applying management practices such as correct stocking rates and providing feed supplementation. According to Webb et al. (2017), the summer calving season and early winter weaning result in good weaning weight of calves as well as good body condition of cows after weaning.
The effects of seasons on ICPs were demonstrated in the current study with long ICPs experienced in cows that calved in autumn and winter. Analysis to determine the effect of the season of calving on the ICP indicates that cows bred in summer and spring had a significantly shorter ICP expressed as a mean for all the cows. These two seasons both fall in the rainfall season when there is re-growth of grass. Improved quality and quantity of grass improved the performance of cattle. According to Bergh (2004), breeding in autumn and winter results in expensive production since the breeder has to use supplements to ensure growth of the calf and maintain good body condition of the cow. Breeding outside summer and spring can easily result in a long ICP due to the negative nutritional impact on the cow.
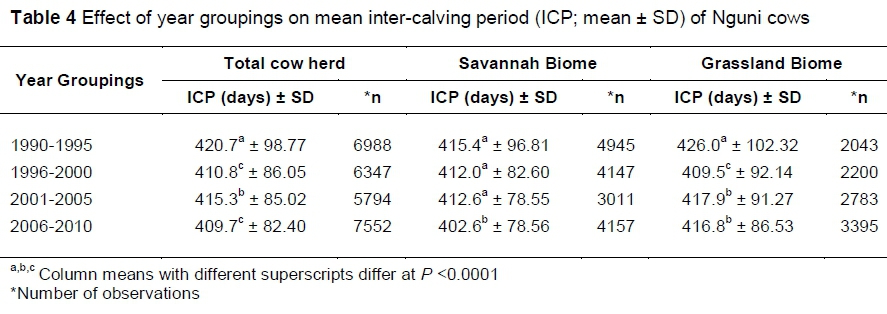
Year groupings of cows also influenced (P <0.0001) the ICP. Cows that calved in year group 2 (1996‑2000) and year group 4 (2005-2010) had a significantly shorter mean ICP (Table 4) than cows categorized in the other group years. The variances can probably be attributed to different weather conditions experienced in the year groupings as well as the management of the breeders.
The interaction between biome and year groupings had an influence (P <0.0001) on the ICP of cows. Cows categorized in year group 4 (2006-2010) of the Savannah Biome had a significantly shorter mean ICP than cows categorized in the other three year groupings. In the Grassland Biome, cows in year group 2 had a shorter mean ICP than those in the other year groupings in the same biome, but the differences were not significant. The effect of year groupings on the ICP of cows can only be explained by differences in prevailing climatic conditions in the particular years, which include rainfall and temperatures. However, information about temperature patterns for the 20-year period could not be verified in this study.
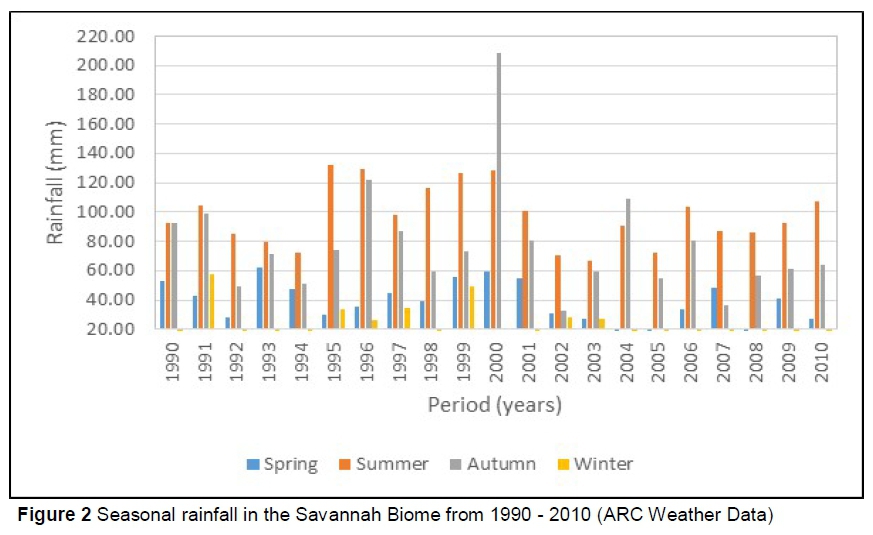
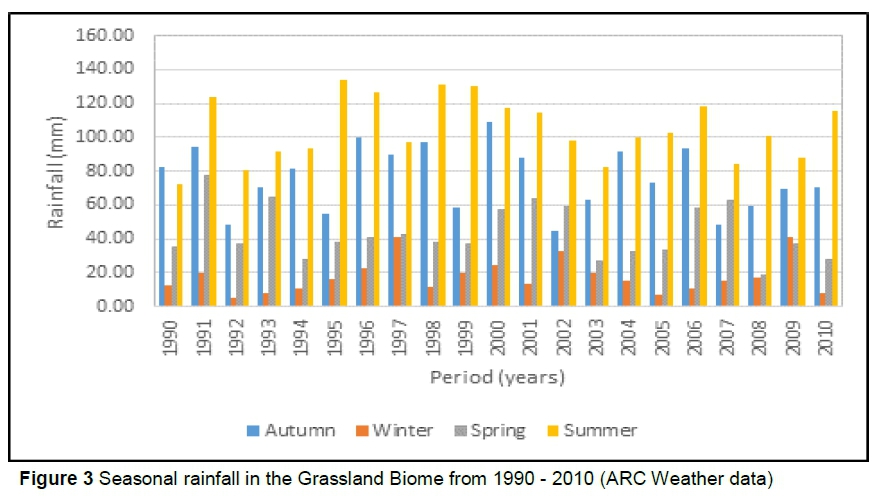
Figure 2 shows the rainfall in the Savannah Biome, while Figure 3 shows the rainfall pattern in the Grassland Biome.
From Figures 2 and 3 it is clear that both Savannah and Grassland Biomes had a higher rainfall in spring and summer, yielding more forage. This may explain the improved cow performance in the two seasons, compared to that of cows bred in autumn and winter. In the Grassland Biome, a higher rainfall was experienced in autumn, compared to the Savannah Biome. This will not contribute much to the quality and quantity of forage as the temperatures are too low to promote quality re-growth. It is evident from the performance of the cows kept in the two biomes that the Nguni breed does not need much interference nutritionally from the environment due to their ability to adapt to environmental changes. It appears that improving the nutrition of the cows through management (e.g. better pasture management or feed supplementation) may reduce the effects of winter and can give good results in terms of the production and reproduction of cows. The present study suggests that feed supplementation is not vital for Nguni cattle if enough forage is available, but it may improve cow production marginally.
It should be noted that in the present study, ICP data that fell outside the limits of the Nguni Cattle Breeders Society standards were discarded, which implies that cows with a long ICP should be culled. However, the decision to retain or cull cows differs among breeders and may explain some of the differences in cow performance observed. One of the practical implications of this is that breeders who cull less productive cows will be left with a smaller number of cows at every given breeding season, which may limit the availability of replacement cows. This supports the recommendation by Donkin (1973) that the ICP alone cannot be used as a measure of reproduction efficiency in beef cattle simply because it is easy to measure.
Conclusions
The results of this study indicate that dam age significantly influenced the ICP of Nguni cows, probably due to environmental factors such as the availability and quality of feed and to some extent the management of breeders.
The bioregions significantly affected the ICP of cows owing to differences in nutritional value of vegetation in different biomes. Seasonal differences in ICP were also observed due to the availability of nutritious herbage and weather conditions such as humidity, wind speed, and cold weather. The average ICP of Nguni cattle is 404 days (Scholtz, 2010) while the average ICP of cows in this study was 405 days. It follows that in both the Savannah and the Grassland Biomes the Nguni cows are able to perform as well as can be expected of an adapted animal. Cows kept in the Eastern Kalahari Bushveld of the Savannah Biome performed better than cows in the other bioregions of the same biome.
Acknowledgements
The authors would like to thank the Nguni Cattle Breeders Society for giving them permission to use the data.
Authors' Contributions
The study was conducted by FNM through interaction with Nguni cattle breeders in KZN. Guidance and supervision were provided by ECW and MMS.
Conflict of Interest Declaration
The authors declare that they have no conflicts of interest with regard to this work.
References
Acocks, J.P.H., 1988. Veld types of South Africa. Mem. Bot. Surv. S. Afr. 57. Botanical Research Institute, Department of Agriculture and Water Supply, South Africa. [ Links ]
Bergh, L., 2004. Breeding seasons for beef cattle in South Africa. S. Afr. J. Anim. Sci. 5, 11-17. [ Links ]
Bonsma, J.C., 1983. Man must measure. Livestock Production. Agi Books, Cody, W.Y., USA. [ Links ]
Burns, B.M., Fordyce, G. & Holroyd, R.G., 2010. A review of factors that impact on the capacity of beef cattle females to conceive, maintain a pregnancy and wean a calf. Implications for reproductive efficiency in northern Australia. Anim. Reprod Science. 122 (1-2), 1-22. [ Links ]
Collins-Lusweti, E., 2000. The performance of the Nguni, Afrikander and Bonsmara cattle breeds in developing areas of Southern Africa. S. Afr. J. Anim. Sci. 30, 28-29. [ Links ]
Donkin, E.F., 1973. A survey of management policies for fertility in Rhodesian Dairy herds. S. Afr. J. Anim. Sci. 3, 91-93. [ Links ]
Escrivao, R.J.A., Webb, E.C., Garces, A.P.J.T. & Grimbeek. R.J., 2012. Effects of 48-hour calf withdrawal on conception rates of Bos indicus cows and calf weaning weights in extensive production systems, Trop. Anim. Health Prod. 44, 1779-1782. [ Links ]
Gregory, K.E., 1972. Livestock production systems for increased yield on resources. S. Afr. J. Anim. Sci. 2, 139-149. [ Links ]
Grobler, S.M., Scholtz, M.M., Greyling, J.P. & Neser, F.W.C., 2014. Reproduction performance of beef cattle mated naturally following synchronization in the Central Bushveld bioregion of South Africa. S. Afr. J. Anim. Sci. 44, 570-574. [ Links ]
Ladermann, E.E. & Schoeman, S.J., 1994. Factors influencing re-calving rate in lactating beef cows in the sweet dry bushveld of Northern Transvaal. S. Afr. J. Anim. Sci. 24, 30-33. [ Links ]
Low, A.B. & Rebelo, A.G., 1996. Vegetation of South Africa, Lesotho and Swaziland. A companion to the Vegetation Map of South Africa, Lesotho and Swaziland. Department of Environmental Affairs and Tourism. Pretoria. [ Links ]
Maree, C. & Casey, N.H., 1993. Livestock Production Systems. Agri Development Foundation, Brooklyn, South Africa. [ Links ]
Messine, O., Schwalbach, L.M.J. & Greyling, J.P.C., 2004. The effects of restricted suckling and early weaning on cow reproduction and weaner production performance in Gudali cattle. S. Afr. J. Anim. Sci. 34,119-121. [ Links ]
Muchenje, V., Dzama, K., Chimonyo, M., Raats, J.G. & Strydom, P.E., 2008. Meat quality of Nguni, Bonsmara and Aberdeen Angus steers raised on natural pasture in the Eastern Cape, South Africa. Meat Sci. 79, 20-28. [ Links ]
Mukasa-Mugerwa, E., 1989. A review of reproductive performance of female Bos indicus (Zebu) Cattle. International Livestock Centre for Afrika (ILCA). Addis Ababa, Ethiopia. [ Links ]
SAS, 2017. Statistical Analysis Systems user's guide (Statistics Version 9.2). SAS Institute Inc. Cary, North Carolina, USA. [ Links ]
Scholtz, M.M., 2010. Beef Breeding in South Africa. 2nd ed. Scholtz, M.M. (ed). Agricultural Research Council, Pretoria. [ Links ]
Schoeman, S.J., 1989. Recent research into production potential of indigenous cattle with special reference to the Sanga. S. Afr. J. Anim. Sci. 19, 55. [ Links ]
Tainton, N.M., 1999. Veld Management in South Africa. University of Natal Press. Pietermaritzburg. [ Links ]
Van der Westhuizen, J.M., 1973. Relationship of thyroid and adrenal function to growth rate in Bos indicus and Bos taurus cattle. S. Afr. J. Anim. Sci. 3, 25-27. [ Links ]
Webb, E.C., Visagie, P., Van der Westhuizen, J. & Snyman, H.A., 2017. The influence of the production environment and environmental factors on the growth, size and reproduction of Bonsmara cows. S. Afr. J. Anim. Sci. 47, 542-552. [ Links ]
Received 6 May 2018
Accepted 8 November 2018
First published online 4 January 2019
# Corresponding author: Edward.webb@up.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}