










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.5 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i5.13
ARTICLES
Serum biochemical profile of two broiler strains supplemented with vitamin E, raw ginger (Zingiber officinale) and L-carnitine under high ambient temperatures
Z. RehmanI; N. ChandI; R.U. KhanII; S. NazIII, #; I.A. AlhidaryIV
IDepartment of Poultry Science, Faculty of Animal Husbandry & Veterinary Sciences, The University of Agriculture, Peshawar, Pakistan
IIDepartment of Animal Health, Faculty of Animal Husbandry & Veterinary Sciences, The University of Agriculture, Peshawar, Pakistan
IIIDepartment of Zoology, GC University Faisalabad, Pakistan
IVDepartment of Animal Production, College of Food and Agriculture Sciences, King Saud University, Riyadh, Kingdom of Saudi Arabia
ABSTRACT
In this study, the authors evaluated the comparative effects of dietary supplementation of ginger (Zingiber officinale), vitamin E and L-carnitine on certain blood parameters of two strains of broilers in summer. A total of 320 broiler chicks (Hubbard and Cobb) were divided into two groups. The two groups were subdivided into four subgroups with four replicates with '0 chicks per replicate. In each strain subgroup was kept as control while other subgroups were provided with ginger (2%), L-carnitine (500 mg/kg) or vitamin E (250 mg/kg) in the basal diet up to 42 days old. Supplementation of the diet with vitamin E, ginger and L-carnitine significantly decreased mean cholesterol, alanine aminotransferase (ALT), glucose, triglyceride, low density lipoprotein (LDL), and increased high density lipoprotein (HDL) and serum protein concentrations. The results of vitamin E were comparatively better than those of the other two feed additives. The blood biochemical parameters did not vary between the broiler strains. From the findings of the current study, it was concluded that high ambient temperature had a negative effect on broiler hepatic and lipid profiles, and the supplementation of diets with vitamin E, ginger and L-carnitine improved broiler blood biochemical parameters.
Keywords: Cholesterol, glucose, liver health, antioxidants, heat stress
Introduction
A large number of broiler farms are in tropical and subtropical zones of the world. The majority still operate under the open-sided housing system (Khan et al., 2012a), which adversely affects the physiological biochemistry of broilers reared under high ambient temperatures (Chand et al., 20'6). In tropical and subtropical countries, the primary problem in summer is heat stress. Since poultry farmers cannot afford expensive artificially temperature controlled sheds, heat stress affects broiler production and physiology negatively (Khan et al., 2011).
A blood biochemical profile provides important information about the health of animals. Higher environmental temperature increases serum alanine aminotransferase (ALT, EC 2.6.1.2) and aspartate aminotransferase (AST, EC 2.6.1.1) activities (Shah et al., 2016), and glucose (Ihsanullah et al., 2017), cholesterol and triglyceride concentrations (Rehman et al., 2018) in broiler chicks. Various ways are being utilized to reduce the hazards of extreme environmental temperatures in poultry production. To avoid the high expense of cooling animal buildings, the focus has been diverted mostly to additives (Khan et al., 2011). The nutritional strategy in summer high temperatures is based on balancing protein, energy and electrolytes in the diet to cover the needs in heat-stressed birds (Laudadio et al., 2012; Khan et al., 2014).
Supplementation of vitamin E in the diet compensates for the negative effects of heat stress in poultry and significantly improves the blood biochemical profile in broilers (Shah et al., 2016). Ginger (Zingiber officinale) is an herbal plant that is being used widely to treat a variety of disorders (Khan et al., 2012b).
Important compounds in ginger include gingerdione, gingerdiol and shogaols (Rehman et al., 2017). These have antimicrobial, antioxidative and other pharmacological properties (Rehman et al., 2011; Khan et al., 2013). Rehman et al. (2011) found that a ginger extract reduced serum ALT and AST activities, and glucose concentration, while serum protein level increased significantly in broilers. L-carnitine is a vital micronutrient and is generated from amino acids (Bremer, 1983). Hassan et al. (2011) reported that supplementation of L-carnitine at rates of 100 mg/kg and 200 mg/kg decreased serum ALT and AST activities and cholesterol and triglyceride concentrations in broilers. Zhang et al. (2010) found that the cholesterol profile was significantly decreased in broilers supplemented with acetyl-L-carnitine.
Little research has been conducted to compare the effects of feed supplements at recommended concentrations under heat-stress conditions in various broiler strains. Thus, the present study was planned to investigate the suitability and efficacy of vitamin E, ginger and L-carnitine on biochemical and lipid profile of two strains of broilers that were exposed to high summer ambient temperatures.
Materials and Methods
A total of 320 one-day-old broiler chicks of two strains (Hubbard and Cobb) were obtained from the local hatchery. The chicks were divided randomly into two groups (160 chicks/group). Each group (strain) was divided further into four subgroups (40 chicks/group) with four replicates (10 chicks per replicate) per group. In each strain, one group was kept as a control, while the basal diets of the other subgroups were supplemented provided with ginger (2%), L-carnitine (500 mg/kg) or vitamin E (250 mg/kg). For the first 14 days the chicks were reared according to brooding protocol. Sawdust was used as bedding material. On day 15, the chicks were exposed to heat stress. The stocking density of birds was 0.9 m2/bird. Birds had free access to feed and water throughout the experiment. A 24-hour light programme was provided.
The broilers were provided with a balanced diet according to the recommendations of the National Research Council (NRC, 1994), as shown in Table 1. Fresh ginger was obtained from a vegetable market. It was sundried and then ground to powder. The powder was added to the feed at the given concentration (2%). Supplementation started on day 15 and continued until the end of the experiment (42 days). Mortalities were recorded as they occurred.
House temperature was recorded continuously throughout the experimental period. Temperature and humidity were recorded every four hours in the centre of the house. The mean values of the temperature and humidity are presented in Table 2.
About 5 mL blood samples were collected weekly from randomly selected birds (three birds per replicate) for biochemical analysis. Serum was separated and analysed for glucose, cholesterol, triglyceride, total protein, AST, ALT, LDL and HDL with commercial kits with a spectrophotometer (Randox Laboratories, CA, USA)
Data were subjected to a two-factorial completely randomized design with four diets and two strains. The least significant test was used to compare the differences among treatments. All statistical analysis was carried out using statistical package SAS version 6.12 (SAS Institute, Cary, N.C., USA) with a completely randomized design.
Results
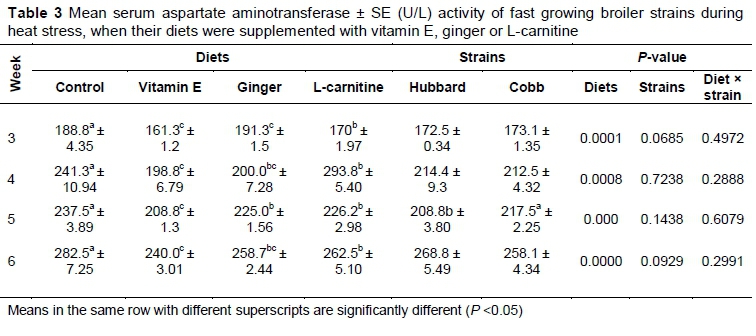
Mean serum AST activity was significantly lower in birds fed vitamin E and ginger supplementation on days 21, 28 and 35 compared with the control (P <0.05) (Table 3). There was no significant difference between ginger and L-carnitine on days 28, 35 and 42 (P >0.05).
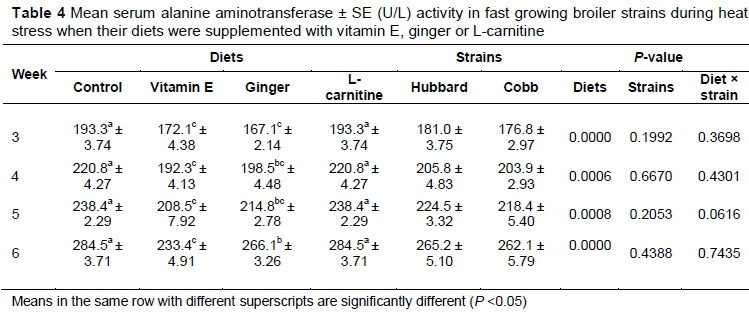
Mean serum ALT activity was significantly lower in the vitamin E-supplemented group compared with the control (P <0.05) (Table 4). However, serum ALT activity did not differ between the control and the birds treated with L-carnitine. In addition, on days 21, 28, and 35, mean activity in serum did not differ between the vitamin E- and ginger-supplemented groups. On day 42, serum ALT activity decreased significantly in vitamin E-supplemented birds compared with the ginger-fed birds (P <0.05).
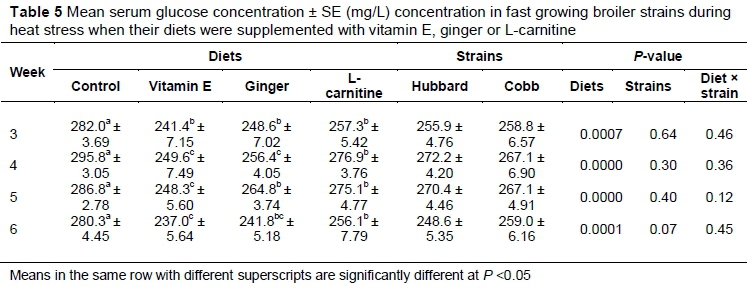
Throughout the experiment, mean serum glucose concentration was significantly (P <0.05) lower in the supplemented groups compared with the control (Table 5). There was no significant difference (P >0.05) on day 28 between vitamin E- and ginger-treated birds. On days 28 and 35 serum glucose concentration was significantly (P <0.05) lower in vitamin E-supplemented birds compared with ginger and L-carnitine supplemented groups.
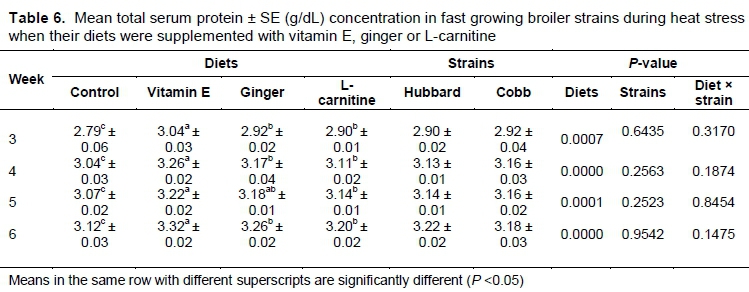
The mean serum protein level was significantly (P <0.05) higher in birds fed with vitamin E supplementation throughout the experiment (Table 6). There was no significant difference (P >0.05) between ginger- and L-carnitine-supplemented birds throughout the experiment (P <0.05).
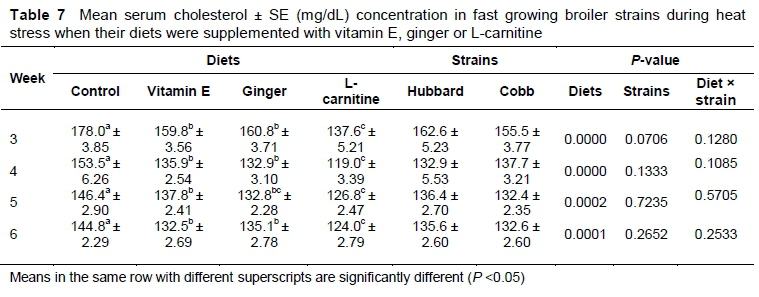
Mean serum cholesterol was significantly (P <0.05) lower in birds supplemented with L-carnitine compared to the other treatments (Table 7). There was no significant difference between ginger-and vitamin E-supplemented birds in mean serum cholesterol concentration (P >0.05).
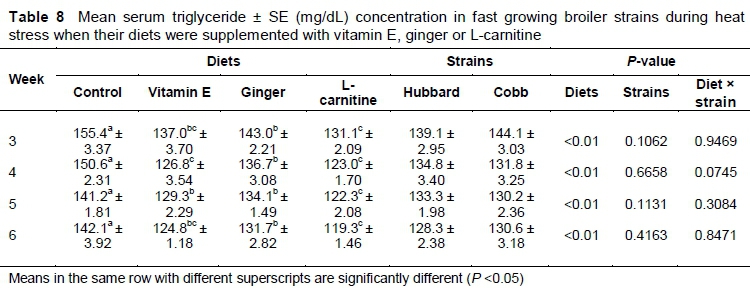
The results indicated that throughout the experiment mean serum triglyceride concentration was significantly (P <0.05) lower in birds supplemented with L-carnitine compared to the other treatments (Table 8). There was no significant difference in serum triglyceride between the vitamin E- and L-carnitine-supplemented birds except on day 35 (P <0.05).
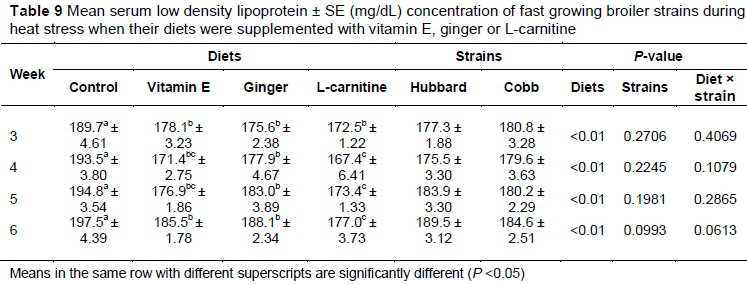
On day 21 the concentration of LDL was significantly (P <0.05) lower in the treatment groups compared with the control (Table 9). At the other stages of the experiment, LDL was significantly (P <0.05) lower in the L-carnitine-treated group compared with the control. There was no significant difference between vitamin E- and ginger-supplemented groups throughout the experiment (P >0.05).
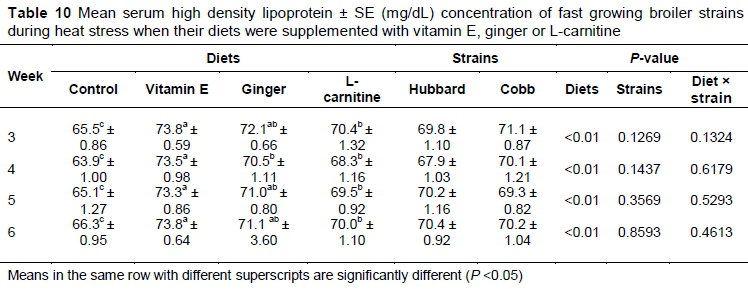
There was no significant difference in HDL concentration between ginger and L-carnitine-supplemented birds on all recorded stages of the experiment (Table 10). There was no significant (P >0.05) differences between the two strains during the experimental period.
Discussion
In the current study, bodyweight was improved significantly in vitamin E-supplemented birds compared with the control. Similarly, improved bodyweights were observed in the ginger- and L-carnitine-supplemented broilers. The improved feed intake and weight gain in vitamin E-supplemented birds may be due to the antioxidant effect, the secretion of the growth hormones, and better feed utilization and digestion (Khan et al., 2013). L-carnitine is known for producing energy from the fat reserve of the body and reducing oxidative stress (Derin et al., 2006). On the other hand, ginger exhibits antimicrobial activity and higher palatability, and releases digestive enzymes in the body, which lead to improved feed consumption and weight gain (Khan et al., 2012b).
The results of the current study revealed that supplementation of vitamin E, ginger and L-carnitine decreased serum liver enzyme activities in broilers. Higher activities of serum AST and ALT are the result of liver disease, probably because of oxidative damage to the tissue (Khan et al., 2013). The increase in serum enzyme activities at high ambient temperature could be the reason for free radicals at cellular level. The decrease in liver enzyme activity in the treated groups may be because of the strong antioxidant properties of vitamin E, ginger and L-carnitine, which scavenge free radicals (Khan et al., 2011, 2013), and maintain the function of antioxidant enzymes (Khan et al., 2013). The current findings are in line with the results of previous studies, which reported that vitamin E supplementation significantly decreased liver enzyme activities in broilers exposed to high ambient temperature (Sahin et al., 2002; Khan et al., 2013). The authors could not locate an authentic review in the literature on the effects of L-carnitine and ginger on serum AST activity under heat-stress conditions in broilers. However, under optimum conditions, a significant decrease has been reported in serum AST activity of broiler chicks fed a ginger-supplemented diet (Rehman et al., 2011). Hassan et al. (2011) reported that a significant decrease in serum AST activity of broiler in response to L-carnitine supplementation in the diet. The reduction in serum liver enzymes in response to ginger may be because of the positive effects of active compounds such as gingerdione, gingerdiol, shogaols and gingerol (Khan et al., 2012b).
The higher serum glucose level in the present study may have been triggered by the secretion of some of the adrenal glands, which accelerated the gluconeogenesis process, ultimately leading to increased serum glucose (Ihsanullah et al., 2017). The decrease in serum glucose level of heat-stressed broilers may be owing to the decrease in synthesis and production of corticosterone by vitamin E (Khan et al., 2011; Khan et al., 2013). Previous studies have reported that vitamin E supplementation in the diet reduced serum glucose levels in broilers (Habibian et al., 2014). The authors could not locate the effects of ginger and L-carnitine on serum glucose concentration in heat-stressed broilers in the literature. Lower serum glucose concentration under optimum conditions with ginger and L-carnitine supplementation had been reported (Saeid et al., 2010; Rehman et al., 2011).
Oxidative stress at cellular level speeds up the production of reactive oxygen species (ROS) that damage biomolecules (Khan et al., 2011; Chand et al., 2018). Reduction in the digestibility of protein under heat-stressed conditions may also lead to low total serum protein concentration (Mohamed et al., 2012). The current findings showed that supplementation significantly reduced serum cholesterol concentration in summer-stressed broilers. L-carnitine was more effective than vitamin E and ginger in reducing serum cholesterol concentration of heat-stressed broilers. Higher serum cholesterol levels at high ambient temperature may be caused by hyperstimulation of the adrenal gland which produces adrenocorticotropin hormones (Ihsanullah et al., 2017) for which cholesterol acts as a precursor. Vitamin E supplementation in poultry decreased the production and synthesis of adrenocorticotropic hormones and corticosterone (Khan et al., 2013). Consequently, serum cholesterol concentration is decreased. ß-hydroxy-b-methyleglutaryl coenzyme A (HMG-CoA) is involved in cholesterol synthesis and ginger is a good HMGR-inhibiting drug and hence reduces cholesterol synthesis (Khan et al., 2012). L-carnitine supplementation in birds may lead to hyperactivity of lipid metabolism (Parizadian et al., 2011).
High ambient temperature releases corticosterone (Khan et al., 2011). Vitamin E supplementation reduces the production of adrenocorticotropic hormones and corticosterone (Khan et al., 2013) and hence decreases the production of triglyceride. Decreased serum triglyceride level in broilers fed an L-carnitine diet may probably be due to increased fatty acid oxidation. The serum triglyceride level is reduced because of the increased transfer of fatty acids to the inner mitochondrial membrane (Parizadian et al., 2011). Supplementation of L-carnitine in diets accelerates the activity of lipase enzyme, leading to increased hydrolysis of triglyceride to fatty acid and glycerol (Zhang et al., 2010).
L-carnitine decreases the activity and the rate of HMG-coA reductase in the liver and may be responsible for lower serum LDL in the L-carnitine-supplemented group in the present study (Hassan et al., 2011). Vitamin E is a major antioxidant, protects lipoprotein from oxidative injuries and may be responsible for the increased HDL level in the vitamin E-supplemented group. Habibian et al. (2014) reported that vitamin E supplementation in broiler diet significantly increased the serum HDL of broilers under heat stress. With increasing age, the birds' response did not change significantly, probably because the average temperature and humidity did not change during the experimental period.
Furthermore, this is perhaps the first study that compares the effects of various antioxidants in heat-stress conditions in broiler strains. In the present study, vitamin E was found to be more effective than ginger and L-carnitine, possibly because vitamin E seems to be a superior antioxidant.
Conclusion
Supplementation of diets with vitamin E, ginger and L-carnitine improved the hepatic and lipid profile of summer-stressed broilers. Moreover, in terms of blood biochemical parameters, the two broiler strains responded equally to heat stress and the treatments.
Acknowledgements
We are thankful to Higher Education Commission (HEC) for providing financial assistance for this study.
Authors' Contributions
Z.R.: conducted the study; N.C. and R.U.K: designed the study; S.N.: edited the paper; I.A.A: revised the study.
Conflict of Interest Declaration
Authors declare no conflict of interest.
References
Bremer, J., 1983. Carnitine-metabolism and functions. Phys. Rev. 63, 420-480. [ Links ]
Chand, N., Muhammad, S., Khan, R.U., Alhidary, I.A. & Zia ur Rahman., 2016. Ameliorative effect of synthetic γ-aminobutyric acid (GABA) on performance traits, antioxidant status and immune response in broiler exposed to cyclic heat stress. Environ. Sci. Poll. Res. 23, 23930-23935. [ Links ]
Derin, N., Izgut, U.V.N., Agac, A., Aliciguzel, Y. & Demir, N., 2004. L-carnitine protects gastric mucosa by decreasing ischemia-reperfusion induced lipid per oxidation. J. Physiol. Pharmacol. 55, 595-606. [ Links ]
Habibian, M., Ghazi, S., Moeini, M.M. & Abdolmohammadi, A., 2014. Effects of dietary selenium and vitamin E on immune response and biological blood parameters of broilers reared under thermoneutral or heat stress conditions. Int. J. Biometeorol. 58, 741-752. [ Links ]
Hassan, M.S.H., Youssef, S.F. & El-bahy, N.M.A., 2011. Effects of L-carnitine and ascorbic acid supplementation on productive, reproductive, physiological and immunological performance of golden Montazah laying hens. Egypt Poult. Sci. 31 (II), 557-578. [ Links ]
Ihsanullah, Qureshi, M.S., Suhail, S.M. & Khan, R.U., 2017. Postpartum endocrine activities, metabolic attributes and milk yield are influenced by thermal stress in crossbred dairy cows. Int. J. Biometeor. 61, 1561-1569. [ Links ]
Khan, R.U., Naz, S., Nikousefat, Z., Tufarelli, V., Javadani, M., Rana, N. & Laudadio, V., 2011. Effect of vitamin E in heat-stressed poultry. Wrld's Poult. Sci. J. 67, 469-478. [ Links ]
Khan, R.U., Naz, S., Nikousefat, Z., Tufarelli, V., Javdani, M., Qureshi, M.S. & Laudadio, V., 2012a. Potential applications of ginger (Zingiber officinale) in poultry diets. Wrld's Poult. Sci. J. 68, 183-190. [ Links ]
Khan, R.U., Naz, S., Nikousefat, Z., Selvaggi, M., Laudadio, V. & Tufarelli, V., 2012b. Effect of ascorbic acid in heat-stressed poultry. Wrld's Poult. Sci. J. 68, 477-490. [ Links ]
Khan, R.U., Rehman, Z., Javed, I. & Muhammad, F., 2013. Supplementation of vitamins, probiotics and proteins on oxidative stress, enzymes & hormones in post-moult male broiler breeders. Archiv. Tierzucht 61, 607-616. [ Links ]
Khan, R.U., Naz, S. & Dhama, K., 2014. Chromium, pharmacological applications in heat stressed poultry. Int. J. Pharmacol. 10, 213-317. [ Links ]
Laudadio, V., Dambrosio, A., Normanno, G., Khan, R.U., Naz, S., Rowghani, E. & Tufarelli, V., 2012. Effect of reducing dietary protein level on performance responses and some microbiological aspects of broiler chickens under summer environmental conditions. Avian Biol. Res. 5 (2), 88-92. [ Links ]
Mohamed, E.A.A., Ali, O.H.A., Huwaida, E.E. & Malik, Y.I.A., 2012. Effect of season and dietary protein level on some haematological parameters and blood biochemical compositions of three broiler strains. Int. J. Poult. Sci. 11 (12), 787-793. [ Links ]
Parizadian, B., Ahangari, Y.J., Shams Shargh, M. & Sardarzadeh, A., 2011. Effects of different levels of L-carnitine supplementation on egg quality and blood parameters of Laying Japanese quail. Int. J. Poult. Sci. 10 (8), 621-625. [ Links ]
Rehman, S., Durrani, F.R., Chand, N., Khan, R.U. & Rehman, F.U., 2011. Comparative efficacy of different schedules of administration of medicinal plants infusion on hematology and serum biochemistry of broiler chicks. Res. Opin. Anim. Vet. Sci. 1, 8-14. [ Links ]
Rehman, Z., Chand, N., Khan, R.U., Khan, S. & Qureshi, M.S. 2018. An assessment of the growth and profitability potential of meat-type broiler strains under high ambient temperature. Pak. J. Zool. 50, 429-432. [ Links ]
Saeid, J.M., Arkan, B.M. & Al-Baddy, M.A. 2010. Effect of aqueous extract of ginger (Zingiber officinale) on blood biochemistry parameters of broiler. Int. J. Poult. Sci. 9, 944-947. [ Links ]
Sahin, K., Kucuk, O., Sahin, N. & Sari, M., 2002. Effects of vitamin C and vitamin E on lipid peroxidation status, some serum hormone, metabolite and mineral concentrations of Japanese quails reared under heat stress (34 °C). Int. J. Vit. Nutr. Res. 71, 91-100. [ Links ]
Shah, A.A., Khan, M.S., Khan, S., Ahmad, N., Alhidary, I.A. & Khan, R.U., 2016. Effect of different levels of alpha tocopherol on performance traits, serum antioxidant enzymes and trace elements in Japanese quail (Coturnix coturnix japonica) under low ambient temperature. Rev. Bras. Zootec. 45, 622-626. [ Links ]
Zhang, Y., Ma, Q., Bai, X., Zhao, L., Wang, Q. & Ji, C. 2010. Effects of dietary acetyl- L-carnitine on meat quality and lipid metabolism in Arbor Acres broilers. Asian-Austral. J. Anim. Sci. 23, 1639-1644. [ Links ]
Received 8 March 2018
Accepted 30 July 2018
First published online 20 November 2018
# Corresponding author: drshabananaz@gcuf.edu.pk


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}