










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.5 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i5.11
ARTICLES
Effect of Vachellia erioloba and Dichrostachys cinerea pod supplementation on performance of does and kids of Namibian Caprivi and Ovambo indigenous goats
L.N. MariusI, #; E.L.K. OsafoII; I.D.T. MpofuIII; E. LutaayaIV; K.L. ShiningavamweI; E. MissanjoV; V. Attoh-KotokuII
IMinistry of Agriculture, Water and Forestry, Division of Livestock Research and Production, Private Bag 13184, Windhoek, Namibia
IIKwame Nkrumah University of Science and Technology, Department of Animal Science, Kumasi, Ghana
IIIChinhoyi University of Technology, Chinhoyi, Zimbabwe
IVUniversity of Namibia, Department of Animal Science, Windhoek, Namibia
VMalawi College of Forestry and Wildlife, Department of Forestry, Dedza, Malawi
ABSTRACT
An on-station feeding trial was conducted to determine the effects of Vachellia erioloba (VE) and Dichrostachys cinerea (DC) pod supplementation on intake, doe weight changes and growth of kids. Forty-two indigenous lactating does with average weight of 35 ± 0.5 kg from Parities 2 to 4 and grouped by ecotype (Caprivi and Ovambo) were allocated to a 2 x 3 factorial arrangement of treatments in a randomized complete block design with six does per treatment. The main factors were two pod types (VE, DC) at three feeding levels (20%, 40%, 60%), benchmarked against positive control (commercial feed (COM)) resulting in seven treatment diets. Phosphate salt lick and water were available ad libitum. Data were analysed as repeated measures using Proc Mixed (SAS, 2008). Voluntary feed intake (VFI) increased steadily for all treatments, peaking at about Week 5, followed by a drop, and then gradual stabilization in Weeks 7 to 12. Intake of does dropped drastically with VE60 and DC60 diets. Does in Parity 3 had significantly higher VFI than those in Parity 2, but there were no differences with Parity 4 does. Does in Parities 3 and 4 were significantly heavier than those in Parity 2. Doe weights declined from Week 1 to Week 8 and then increased gradually. Male kids were heavier than female kids (13.3 ± 0.2 vs. 10.5 ± 0.1 kg). The regression of kid weights on age (days) was 0.030 ± 0.012. Kids from does in Parities 3 and 4 had significantly higher weights than those from does in Parity 2. Single kids were heavier than twins (13.3 ± 0.2 vs.10.5 ± 0.1 kg). Kid weights from does supplemented with DC20 were similar to those receiving COM, VE20 and VE40, but kid weights of does supplemented with VE60, DC40 and DC60 were inferior to other treatments. Supplementing lactating goats with pod-based diets, particularly V. erioloba at 40%, may lead to increased VFI, reduced bodyweight loss in does and increased kid pre-weaning weights.
Keywords: Lactating does, pod-based diets, voluntary feed intake, doe weights, pre-weaning weights of kids
Introduction
Livestock productivity in semi-arid areas is constrained by many factors, including poor rainfall, insufficient availability of grazing land, shortage of feed supplements and the use of poor quality and low quantity of crop residues in the long dry season. These result in low milk production, slow growth in young animals and ill-health (Kindness et al., 1999). Natural rangelands in semi-arid areas are a potential source of less expensive local feed resources in the form of grazing and browse materials (Mlambo & Mapiye, 2015), which could be used by resource-poor farmers. However, the use of woody plant materials is constrained by insufficient knowledge and skills regarding harvesting, storage, processing and inclusion amounts in animal feeding rations. The woody browses contain abundant amounts of antinutritional factors such as tannins, which tend to increase in response to season, temperature and light intensity (Mueller-Harvey, 2006; Waghorn, 2008). Woody plant pods, especially those from Vachellia species and D. cinerea, are valuable because they ripen and drop off well into the dry season (Timberlake et al., 1999). Pods are a good source of crude protein (100-190 g/kg dry matter (DM)) and have been reported to increase the growth of kids and of milk production in goats (Mlambo et al., 2004; Maphosa et al., 2009). Mlambo et al. (2004) reported a 60% reduction in urinary nitrogen excretion in goats that was offered untreated D. cinerea fruits. Supplementing lactating goats with 200 g D. cinerea and Vachellia nilotica fruits per day increased the milk yield of does, growth rate and kid survival (Smith et al., 2005). Wang et al. (1996) observed increases in milk yield, protein and lactose percentage in ewes fed Lotus corniculatus pods, which contained moderate amounts (44.5 g/kg DM) of condensed tannins. However, there was a decline in milk fat percentage.
Vachellia erioloba and D. cinerea are shrubs whose pods are commonly used as feed supplements in Namibia, particularly during the dry season, but their nutritive value, concentration of antinutrients and optimal inclusion levels in diets are unknown. The objectives of this study were to determine i) the voluntary feed intake of pod-based supplements (V. erioloba and D. cinerea) for lactating goats; and ii) the effect of these supplements on the weight of lactating does and pre-weaning kids.
Materials and Methods
The feeding experiment was conducted at John Pandeni Research Station, under the Ministry of Agriculture, Water and Forestry (MAWF), between January and April 2015. The research station is located 20 km south-west of Grootfontein in Otjozondjupa Region of Namibia. The station lies at latitude 19° 48'S and longitude of 18° 00'E at an elevation of 1450 m above sea level. The research station has 6559.4 hectares, with mean annual rainfall of about 500 mm, and the temperature ranges between 13.5 °C and 28.7 °C. The main rainy season occurs from December/January to April, and the dry season is from May to December. The natural vegetation is dominated by shrubs of V. erioloba D. cinerea, Tarchonanthus camphoratus, Ziziphus mucronata and Grewia species. The predominant grass species in the original goat camps were Aristidas and Stipagrostis uniplumis, while the rest of the farm has Aristida meridionalis, Digitaria seriata, Heteropogon contortus, Schmidtia pappophoroides and Eragrostis spp.
The MAWF guidelines on the proper treatment of animals were followed. Does were hand-mated from July to September 2014 and kidded between 9 December 2014 and 2 January 2015. Does were kept on rangeland, but kraaled at night. They were given 0.650 kg per animal per day of commercial supplement (crude protein (CP): 140 g/kg; Ca: 10 g/kg; P: 6 g/kg) and lucerne hay two weeks prior to kidding. Before the experiment began, does were dipped in Alphamethrin (Pfizer Laboratories (Pty) Ltd, South Africa) to control external parasites and injected with 1% Dectomax (Pfizer Laboratories (Pty) Ltd, South Africa) against internal worms. The feeding trial was a two-way factorial with goats belonging to two ecotypes (Ovambo and Caprivi) being represented in each treatment. The two factors were pod type, V. erioloba (VE) and D. cinerea (DC), and three level (20%, 40% and 60%). A commercial supplement was used as positive control, giving a total of seven treatments. Forty-two lactating does in Parity 2 or more were stratified by ecotype and randomly allocated to the seven treatments, ensuring that each treatment had three does of each ecotype. Kid and doe weights were recorded prior to supplementation in the mornings, at kidding and weekly until the end of the experiment. Does were given a one-week pre-test period to adjust to the test environment and feed. The feeding trial lasted 12 weeks, excluding the pre-test period. The pens were located in a roofed structure without solid walls. Each doe was placed in a 2 m x 1 m pen with a feeding trough.
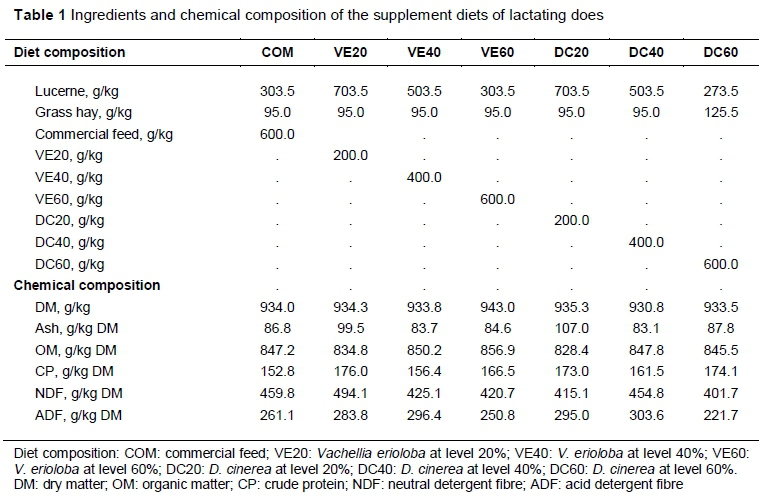
The phosphate salt lick and water were available as free choice to all treatment groups. A commercial supplement was obtained from Feed Master, a commercial feed supplier, whereas D. cinerea and V. erioloba pods were collected from Guinas and Omatako constituencies. Lucerne and grass hay (predominantly Stipagrostis uniplumis) were obtained from the research station. Lucerne, grass hay, VE and DC pods were air-dried and milled to pass through a 10-mm sieve using a Wiley Mill (Glen Creston Ltd, UK). Feeds were formulated from pods, lucerne, grass hay and commercial feed to attain iso-nitrogenous diets. Ingredient and chemical compositions of the supplement diets are outlined in Table 1. Does were supplemented with 200 g/head/day of the treatment diet and the amount was increased by 10% based on intake of the previous day, up to a maximum of 400 g. Does were given the supplemental diet at 07:00 and allowed to feed with their kids at foot for two hours before being led to graze under the guidance of a herder. Does returned to the kraals at 17:00. Feed refused from each doe was weighed and recorded daily in order to determine the DM intake. Samples of feed refused per doe were taken daily and pooled over each week. At the end of the experiment the weekly pooled samples were further pooled to yield a composite sample of refusal for each doe for chemical analysis. Chemical composition of the feed offered was determined from samples taken from each batch using AOAC (2005) method. Animal identification, mating dates, kidding dates, weight of doe at kidding, kid weight at birth, sex of kid, type of birth (single or twin) and parity of doe were recorded.
The data were checked for normality and outliers before analysis. Seventy-two kids were born from the 42 does. One kid, which was born as a triplet, was abandoned by its dam, had a much slower growth, rate and hence was excluded from the analysis. Data were modelled as repeated measures using Proc Mixed (SAS, 2008), which takes into account correlation due to a common contribution from the animal and also time trends, whereby measurements close in time may be more correlated than those far apart (Littell et al., 1998). The model for VFI of does on DM basis included treatment, week, parity and treatment-week interactions. The model for doe weights included treatment, week and parity. The model for kid weights included sex, type of birth (single or twin), week, treatment, age as a covariable, and parity of the dam. The appropriate covariance structure was selected based on BIC criterion (Littell et al., 1998). The ante-dependence (ANTE(1)) covariance structure (SAS, 2008) was selected as best for voluntary intake and kid weights. For doe weights, the Toeplitz (TOEP) covariance structure was selected. Average daily gain (ADG) of kids was estimated by regression of weight on age. Least squares means were separated by a PDIFF statement. Significance was declared at P <0.05. Trends were accepted if 0.05 < P < 0.10.
Results
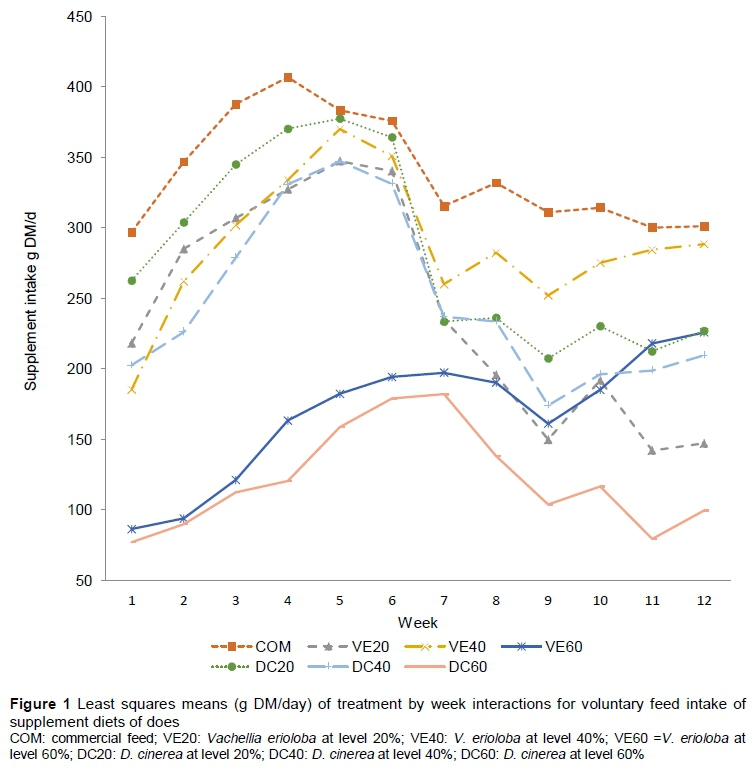
The VFI on DM basis of does was influenced (P <0.05) by treatment, week and treatment x week interactions. Parity showed a trend (P = 0.0754) in influencing VFI. With the exception of VE60, VFI increased steadily for all treatments, peaking at about Week 5, followed by a drop, and then gradual stabilization in Weeks 7 to 12 (Figure 1). While intake of DC20 exceeded that of VE40 for the first six weeks, the reverse occurred for Weeks 7-12. There was a fairly steady increase in VFI for treatment VE60 from Weeks 1 to 12. Does in Parity 3 had a higher (P = 0.0281) VFI than those in Parity 2, but there were no differences (P >0.05) with Parity 4 does. In general, the lowest intakes were observed at the 60% inclusion levels of VE and DC. Voluntary intake of DC20 and DC40 followed a similar pattern throughout the trial period, with intake of DC20 greater (P <0.05) at most time points. On the contrary, intake patterns of VE20 and VE40 were similar in Weeks 1 to 6, but then tended to diverge in Weeks 7 to 12.
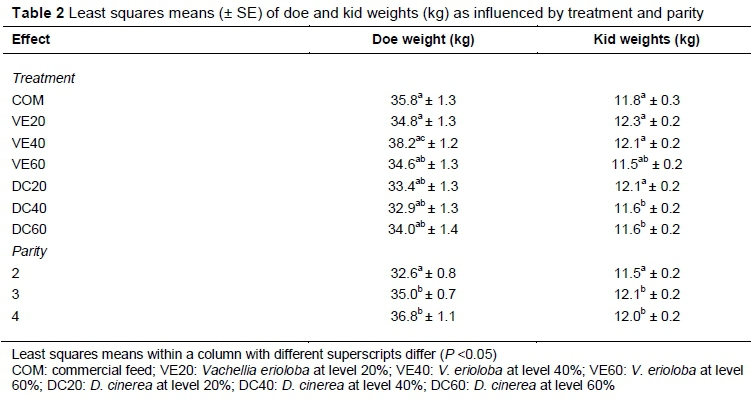
Doe weights were influenced (P <0.05) by week and parity. Treatment showed a trend (P = 0.0857) in influencing doe weights. Table 2 presents the least squares means for doe and kid weights. VE40 resulted in heavier doe weights compared with DC20, DC40 and DC60. VE40 tended to give heavier doe weights than VE20 (P =0.0627). Does in Parities 3 and 4 were heavier (P <0.05) than those in Parity 2; there were no differences (P = 0.1961) between doe weights in Parities 3 and 4. The least squares means (SE = 0.5) for Weeks 1 to 12 were 36.4, 35.5, 35.1, 35.0, 35.4, 35.4, 34.5, 33.2, 33.6, 34.2, 34.5 and 35.0. Doe weights tended to decline from Week 1 up to Week 8 and then gradually increased.
Average daily gain of does was influenced (P <0.01) by ecotype and ecotype x parity interactions, although parity also showed a trend (P = 0.06) in influencing ADG. Does of the Ovambo ecotype lost more weight compared with those of the Caprivi ecotype, but this was more pronounced in Parities 3 and 4. In contrast, does of the Caprivi ecotype belonging to Parity 4 gained weight (P <0.05) relative to those in Parities 2 and 3, which lost weight.
Weights of kids were affected by sex, week, treatment, parity, age and birth type (P <0.05). Male kids were heavier (P <0.0001) than female kids (13.3 ± 0.2 vs. 10.5 ± 0.1 kg). As would be expected, weight of kids increased with week of trial. The linear regression of kid weight on week of trial was 0.206 ± 0.030. The regression of kid weights on age (days) was 0.030 ± 0.012. Does in Parity 3 and 4 had higher (P <0.05) kid weights than those in Parity 2 (Table 2). Kids born as singles were heavier than twins (13.3 ± 0.2 vs.10.5 ± 0.1 kg; P <0.0001). Kid weights from does supplemented with DC20 were lighter (P = 0.0116) than those receiving COM, VE20 and VE40. Kid weights of does supplemented with VE60, DC40 and DC60 were inferior to other treatments (P = 0.0025).
Discussion
The authors hypothesized that VFI would be similar for the various supplements, but differences in chemical composition and inclusion levels in the diet influenced the intake. The presence of treatment x week interactions on intake suggests that there may have been environment influences for example temperature and humidity, which influenced intake indirectly through the decreased quality of feed on the natural range prolonging the rumen-fill effect as the season progressed.
The treatment x week interactions on VFI may also be indicative of adaptation to or lack of it for the various supplements by the indigenous goats. Consumption patterns of VE40 and COM were similar and appeared to be converging with time. Does appeared initially to have increased intake of DC60, but after Week 7 there was a steady drop, suggesting failure to adopt to this dietary supplement. It is doubtful whether the CP concentration could explain the differential response, because all diets were well above the required CP for lactating goats (NRC, 2007). Adaptation to diets may also be influenced by nutrient imbalances and antinutrients, as discussed below.
The gradual increase in intake of VE60 would indicate slower adaptation of the indigenous goats to high inclusion levels of V. erioloba in the diet. Diet composition is known to influence rumen microbial activity (Duncan et al., 2000) and it would appear the efficiency of degradation of the VE60 diet increased with time, leading to the higher intake. Nutrient supply in the rumen may affect changes in microbial populations leading to further alterations in the rumen environment that are conducive to certain microbes. Pods of tropical legumes have high concentrations of carbohydrates, proteins and minerals (Aganga & Tswenyane, 2003), which may have resulted in enhanced microbial activity, leading to rapid degradation of pods, reduced residence time in the rumen and higher intake (Chilibroste et al., 1997; Kyriazakis, 2003). The results suggest a 40% optimal inclusion level of V. erioloba in the supplement to maximize intake. A 20% inclusion level appears to be the optimum for D. cinerea. In in vitro studies, D. cinerea tannins were shown to inhibit fermentation severely (Mlambo et al., 2008). Hence it can be surmised that at high inclusion levels (>20%), feed is degraded more slowly, reducing intake. With VE pods at a high inclusion level (60%), high concentrations of NDF and ADF could limit DM digestibility (Pineiro et al., 2011). Secondary metabolites such as condensed tannins in pods may influence digestibility of rations (Briceno-Poot et al., 2012), hence limiting intake. Mlambo et al. (2008) reported higher concentrations of soluble phenolics in D. cinerea than in V. erioloba (6.5 vs. 5.6 gallic acid equivalent μg/mg DM) and ytterbium precipitated phenolics (485 ± 18 vs. 206 ± 18 YbPh, g/kg DM), which may explain the lower intake of D. cinerea due to lower digestibility and fermentation (Sebata et al., 2011). Besides concentration, tannins may differ in structures (Tibe et al., 2013), which affects their biological activity.
Doe weights were influenced through the consumption of a protein supplement, with VE40 showing a tendency to be superior to other treatments. The VE40 may have stimulated increased ruminal microbial activity, enhancing intake and hence higher nutrient availability to support lactation and bodyweight gains of does. The DC pod supplements resulted in lower bodyweights of does compared with VE, which would indicate that they may require additional processing to enhance their utilization in ruminant diets. Although DC pods had higher CP than VE (161.5 ± 27.5 vs. 141.4 ± 22.5 g DM/kg), they had a higher concentration of soluble tannins (155.0 ± 31.0 vs. 93.0 ± 25.0 g DM/kg) (Marius, 2016), which might have interfered with protein digestion in the rumen and negatively affected intake. In further support of the current results, D. cinerea has been reported to have moderate levels of phenolics (e.g. 485 ± 18.0 g/kg DM) as measured by the Yb assay (Smith et al., 2005). Because these consist partly of tannins, intake may be reduced. Sebata et al. (2011) also attributed the lower in vitro digestibility and fermentation of D. cinerea to the presence of condensed tannins. Animal responses to supplementation with tanniniferous feeds have been attributed to differences in tannin structures and microbial adaptation (Silanikove et al., 1996; Caygill & Mueller-Harvey, 1999). The comparable influence of VE40 to the commercial diet on doe weights is an encouraging result, given the huge cost differential of these two supplements. It should also be borne in mind that pod supplements are available locally and pose no special storage requirements for small-scale farmers in remote rural areas. In another study, although doe weights of Matabele goats grazing on natural pasture were similar (P >0.05) to those that were supplemented with D. cinerea, kid weaning weights were higher (P <0.05) in those supplemented with D. cinerea compared with the unsupplemented group (Mlambo et al., 2004; Maphosa et al., 2009).
Weights of does increased by parity. Curvilinear trends in weights have been reported in livestock as they age (Carnicella et al., 2008). The increase in doe weights by parity reflects increasing maturity. Parallel to the increase in doe weights, kid weights increased, possibly reflecting increased energy partitioning towards milk production. Does in earlier parities need energy for maintenance in addition to body growth and lactation. By contrast, mature does partition energy only towards maintenance and lactation.
Kids whose dams had been supplemented with COM, VE20, VE40 and DC20 had heavier weights compared with other diet formulations, which can be attributed to the increased milk yield of their dams. The maternal effect of dams on the pre-weaning weight of their offspring is well established (Van Vleck et al., 1977) and is moderated through milk production, which is influenced by genetics and environment (e.g. nutrition, disease and housing). In a study with local goats, daily milk yield as estimated by the double weighing-suckling method was highly and positively correlated (r > 0.63, P < 0.01) with kid weight from kidding to 42 days' postpartum (Gonzalez et al., 2017). Gonzalez et al. (2017) also reported reduction in bodyweights of does postpartum, but the body condition score, although low at kidding, increased 42 days postpartum. In the current study, does on VE40 were heavier than those on other supplements, except COM, suggesting adequate energy intake to support higher milk yields and possibly earlier re-conception. This is a promising result, given the relative abundance of VE, which is currently underutilized and could replace expensive commercial feed in Namibia, especially among smallholder farmers.
In an attempt to respond to increased energy requirements, animals with high nutritional requirements (e.g. energy requirements) increase their intake (Hunter & Seibert, 1986), which corresponds to the trends observed in intake in the present study (Figure 1). Consistent with other studies (Gonzalez et al., 2017), does in early lactation lost weight up to about Week 8, which may be explained by inadequate energy intake to meet milk production for their kids in spite of the increased voluntary intake of supplement. Kids in early lactation are dependent solely on their dams' milk for their nutrient intake and, similar to what is observed in high milk producing dairy cattle breeds, does may be drawing on their energy reserves to support the high milk production (Fulkerson et al., 2008), leading to the observed weight losses. Gains in bodyweights of does after the eighth week were possibly due to decreased milk secretion and reduced dependence of kids on their dams for nutrient supply, as they obtain a greater proportion of their nutrient requirements from grazing.
Supplementation of does with VE20, VE40 and DC20 resulted in kid weights that were comparable with those from does fed COM. This suggests that these three pod formulations are able to elicit equivalent rumen microbial activity, which yields nutrient levels that supported the high milk yields over other pod-based treatments. The authors hypothesize that at the highest inclusion level of V. erioloba (VE60), bitterness and odours and high fibre content are deterrents to high intake of the supplement. It is evident that kid weights correspond to the doe weights for the treatments. Doe nutrition influences pre-weaning weight gain through milk production (Willham, 1963), which the present study confirmed. Based on the current results, the most appropriate inclusion level for V. erioloba is VE40, and for D. cinerea it would be DC20. Although a cost/benefit analysis was not done, the authors are inclined to suggest VE40 as the optimal inclusion level, taking into account the cost of lucerne. Nonetheless, future research should explore other cheaper alternatives to lucerne for combining with Vachellia pods, for instance Lablab purpureus. A linear trend was observed in the growth of kids, in contrast to an expected sigmoid growth curve for growing animals under ideal conditions, which suggests environmental constraints (e.g. inadequate nutrition, heat stress, diseases and parasites) in this production system. Nutrition improvement through supplementation of does with pod-based diets could contribute towards realizing the genetic potential in indigenous goats, but research on other environmental constraints is warranted.
Conclusion
Supplementation significantly improved voluntary feed intake of lactating does on 40% V. erioloba (VE40) having the highest values, reduced bodyweight loss in does and increased pre-weaning weights of kids thus leading to higher productivity in extensive systems.
Acknowledgments
The authors gratefully acknowledge funding from Ministry of Agriculture, Water and Forestry in Namibia and EUERASMUS MUNDUS through Intra ACP Mobility Scholarship.
Authors' Contributions
The authors contributed equally to this work.
Conflict of Interest Declaration
There is no conflict of interest.
References
Aganga, A.A. & Tswenyane, S.O., 2003. Feeding value and anti-nutritive factors of forage tree legumes. Pakistan J. Nutr. 2, 170-177. Available at: http://docsdrive.com/pdfs/ansinet/pjn/2003/170-177.pdf. [ Links ]
AOAC, 2005. Official methods of analysis of AOAC International. 18th edition. Association of Official Analytical Chemists, Arlington VA, Gaithersburg, MD, USA. [ Links ]
Briceno-Poot, E.G., Ruiz-Gonzälez, A., Chay-Canul, A.J., Ayala-Burgos, A.J., Aguilar-Pèrez, C.F., Solorio-Sánchez, F.J. & Ku-Vera, J.C., 2012. Voluntary intake, apparent digestibility and prediction of methane production by rumen stoichiometry in sheep fed pods of tropical legumes. Anim. Feed Sci. Technol. 176, 117-122. [ Links ]
Carnicella, D., Dario, M., Ayres, M.C., Laudadio, V. & Dario, C., 2008. The effect of diet, parity, year and number of kids on milk yield and milk composition in Maltese goat. Small Rumin. Res. 77, 71-74. [ Links ]
Caygill, J.C. & Mueller-Harvey, I., 1999. Secondary plant products - Anti-nutritional and beneficial actions in animal feeding. Nottingham University Press, Nottingham, UK. 129 pp. [ Links ]
Chilibroste, P., Tamminga, S. & Boer, H., 1997. Effect of length of grazing session, rumen fill and starvation time before grazing on dry matter intake, ingestive behaviour and dry matter rumen pool sizes of grazing lactating dairy cows. Grass Forage Sci. 52, 249-257. [ Links ]
Duncan, A.J., Frutos, P. & Young, S.A., 2000. The effect of rumen adaptation to oxalic acid on selection of oxalic-acid-rich plants by goats. Br. J. Nutr. 83, 59-65. [ Links ]
Fulkerson, W.J., Davidson, T.M., Garcia, S.C., Hough, G., Goddard, M.E., Dobos, R. & Blockey, M., 2008. Holstein-Friesian dairy cows under a predominantly grazing system: Interactions between genotype and environment. J. Dairy Sci. 91, 826 -839. [ Links ]
Gonzalez, E.G.Y., Flores, J.A., Delgadillo, J.A., González-Quirino, T., Fernandez, I.G., Terrazas, A., Vielma, J., Nandayapa, E., Mendieta, E.S., Loya-Carrera, J. Flores, M.J. & Hernandez, H., 2017. Early nursing behaviour in ungulate mothers with hider offspring CCapra hircusj: Correlations between milk yield and kid weight. Small Rumin. Res. 151, 59-65. http://www.sciencedirect.com/science/article/pii/S0921448817301104#!. [ Links ]
Hunter, R.A. & Siebert, B.D., 1986. The effect of genotype age, pregnancy lactation and rumen characteristics on voluntary intake of roughage diets by cattle. Aust. J. Agric. Res, 37, 546-560. [ Links ]
Kindness, H., Sikosana, J.L.N., Mlambo, V. & Morton, J.F., 1999. Socio-economic surveys of goat keeping in Matobo and Bubi Districts. Natural Resources Institute Report No. 2451. Chatham Maritime, UK. [ Links ]
Kyriazakis, I., 2003. What are ruminant herbivores trying to achieve through their feeding behaviour and food intake? In: Proceedings of the Sixth International Symposium on the Nutrition of Herbivores. Merida, Mexico, 153-173. [ Links ]
Littell, R.C., Henry, P.R. & Ammerman, C.B., 1998. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 76, 1216-1231. [ Links ]
Maphosa, V., Sikosana, J.L.N. & Muchenje, V., 2009. Effect of doe milking and supplementation of Dichrostachys cinerea pods on kid and doe performance in grazing goats during the dry season. Trop. Anim. Heal. Prod. 41, 535-541. [ Links ]
Marius, L.N., 2016. Evaluation of local feed resources, their response on intake, growth, milk yield and composition and product properties of Namibian indigenous goats. PhD thesis, Kwame Nkrumah University of Science and Technology, Kumasi, Ghana. [ Links ]
Mlambo, V. & Mapiye, C., 2015. Towards household food and nutrition security in semi-arid areas: What role for condensed tannin-rich ruminant feedstuffs? Food Research International, Available at: http://dx.doi.org/10.1016/j.foodres.2015.04.011. [ Links ]
Mlambo, V., Smith, T., Owen, E., Mould, F.L., Sikosana, J.L.N. & Mueller-Harvey, I., 2004. Tanniniferous Dichrostachys cinerea fruits do not require detoxification for goat nutrition: In sacco and in vivo evaluations. Livest. Prod. Sci. 90, 135-144. [ Links ]
Mlambo, V., Mould, F.L., Sikosana, J.L.N., Smith, T., Owen, E. & Mueller-Harvey, I., 2008. Chemical composition and in vitro fermentation of tannin-rich tree fruits. Anim. Feed Sci. Technol. 140, 402-417. [ Links ]
Mueller-Harvey, I., 2006. Review Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 86, 2010-2037. [ Links ]
National Research Council (NRC), 2007. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World camelids The National Academies Press, Washington, DC. Available at: https://doi.org/10.17226/11654. [ Links ]
Pineiro, R., Maffucci, T. & Falasca, M., 2011. The putative cannabinoid receptor GPR55 defines a novel autocrine loop in cancer cell proliferation. Oncogene 30, 142-152. [ Links ]
Sebata, A., Ndlovu, L.R. & Dube, J.S., 2011. Chemical composition, in vitro dry matter digestibility and in vitro gas production of five woody species browsed by Matebele goats (Capra hircus L.) in a semi-arid savanna, Zimbabwe. Anim. Feed Sci. Technol. 170, 122-125. [ Links ]
Silanikove, N., Gilboa, N., Perevolotsky, A. & Nitsan, Z., 1996. Goats fed tannin-containing leaves do not exhibit toxic syndromes. Small Rumin. Res. 21, 195-201. [ Links ]
Smith, T., Mlambo, V., Sikosana, J.L.N., Maphosa, V., Mueller-Harvey, I. & Owen, E., 2005. Dichrostachys cinerea and Acacia nilotica fruits as dry season feed supplements for goats in a semi-arid environment. Anim. Feed Sci. Technol. 122, 149-157. [ Links ]
Statistical Analysis System (SAS), 2008. Statistics Software. Release 9.1.3, SAS Institute, Inc., Cary, NC, USA. [ Links ]
Tibe, O., Sutherland, I.A., Lesperance, L. & Harding, D.R.K., 2013. The effect of purified condensed tannins of forage plants from Botswana on the free-living stages of gastrointestinal nematode parasites of livestock. Vet. Paras. 197, 160-167. [ Links ]
Timberlake, J., Fagg, C. & Barnes, R., 1999. Field guide to the Acacias of Zimbabwe CBC, Harare, Zimbabwe. [ Links ]
Waghorn, G., 2008. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production - Progress and challenges. Anim. Feed Sci. Technol. 147, 116-139. [ Links ]
Wang, Y., Douglas, G.B., Waghorn, G.C., Barry, T.N. & Foote, A.G., 1996. Effect of condensed tannins in Lotus corniculatus upon lactation performance in ewes. J. Agric. Sci., Camb. 126, 353-362. [ Links ]
Willham, R.L., 1963. The covariance between relatives for characters composed of components contributed by related individuals. Biometrics 19, 18-27. [ Links ]
Yayneshet, T., Eik, L.O. & Moe, S.R., 2008. Influences of fallow age and season on the foraging behaviour and diet selection pattern of goats (Capra hircus L.). Small Rumin. Res. 77, 25-37. [ Links ]
Van Vleck, L.D., Louis, D.St. & Miller, J.I., 1977. Expected phenotypic response in weaning weight of beef calves from selection for direct and maternal genetic effects. J. Anim. Sci. 44,360. [ Links ]
Received 16 February 2017
Accepted 20 June 2018
First published online 20 November 2018
# Corresponding author: ndamonako@gmail.com


{kind=link}
{kind=link}
{kind=link}